
Возникновение и распространение возбуждения по миокарду кратко
Обновлено: 04.07.2024
Возникновение и проведение возбуждения в сердце
Процесс возбуждения , вызывающий сокращение сердца , возникает у млекопитающих в области устья полых вен. Здесь расположен синоатриальный узел, описанный Кисом и Флеком и часто называемый их именами.
Синоатриальный узел относится к проводящей возбуждение системе сердца. Он состоит из малодифференцированных мышечных волокон, приближающихся по структуре к эмбриональным и морфологически сходных с волокнами Пуркине в желудочках. Подобные же атипичные мышечные волокна рассеяны в близких к синоатриальному узлу участках предсердий, к которых разветвляются волокна узла. В области узла имеется, кроме того, большое число нервных клеток, нервных волокон и их окончаний, образующих ганглиозную нервную сеть.
Доказательства того, что возбуждение первично возникает именно в синоатриальном узле, получены различными способами. Весьма убедительные результаты достигнуты с помощью электрофизиологической методики: путем регистрации посредством тонких электродов, приложенных к разным участкам сердца, изменений разности электрических потенциалов. Таким способом обнаружено, что электрические изменения, являющиеся характернейшим проявлением процесса возбуждения, первично возникают именно в области синоатриалыюго узла и затем уже распространяются на другие участки предсердий и на желудочки.
Другое доказательство места первичного возникновения возбуждения в сердце дают опыты со строго ограниченным охлаждением или согреванием синоатриального узла (опыт Гаскелла). Для этого к области синоатриального узла прикладывают стеклянный капилляр, чорез который протекает ледяная или теплая вода. Местное охлаждение узла вызывает резкое замедление сердечной деятельности или даже временное ее прекращение. Приложение холода к другим участкам сердца не вызывает такого замедления деятельности всего сердца. Противоположный эффект — учащение сердечной деятельности — получается при местном строго ограниченном согревании синоатриального узла. Третьим доказательством того, что возбуждение первично возникает в синоатриальном узле, служат результаты локального повреждения или отравления некоторыми ядами указанного участка сердца. При этом частота сердечных сокращений сильно замедляется и может наступить остановка сердца.
Отчетливые результаты получаются при наложении лигатуры между венозным синусом и предсердиями у лягушки, т. е. при отделении их перевязкой. Такой опыт, произведенный впервые Станниусом, заключается в следующем: если перевязать сердце лягушки по границе между синусом и предсердиями (лигатура Станниуса), то синус продолжает сокращаться в прежнем ритме, а предсердия и желудочек останавливаются.Это обусловлено тем, что перевязка препятствует доступу к предсердиям и желудочку импульсов, рождающихся у лягушки в венозном синусе сердца.
Совокупность указанных опытов свидетельствует, что синоатриальный узел является водителем ритма сердца, т. е. тем очагом, в котором первично возникает возбуждение, вызывающее сердечное сокращение. От синоатриального узла возбуждение переходит к мышечным волокнам правого, а затем левого предсердия. Распространяясь по мышце предсердий, возбуждение доходит до атриовенгрикулярного узла, являющегося частью проводящей системы сердца ( рис. 10 ), функция которой состоит в передаче возбуждения от предсердий к миокарду желудочков.
Рис. 10. Схема строения проводящей системы сердца: 1 — верхняя полая вена; 2 — нижняя полая вена; 3 — ушко правого предсердия; 4—синоатриальнып узел; 5 — атриовентрикулярный узел; 6 — общая ножка пучка Гиса; 7 и 8 — правая и левая ножки пучка Гиса; 9 — папиллярные мышцы; 10 и 11 — правое и левое предсердии; 12 и 13 — правый и левый желудочки.
Атриовентрикулярный узел расположен в правом предсердии, в области межпредсердной перегородки, вблизи соединительнотканного кольца, отделяющего правое предсердие от желудочка. От атриовентрикулярного узла берет начало пучок Гиса, который представляет собой мышечный мостик, проводящий возбуждение от предсердий к желудочкам. Начальная часть этого пучка (ее называют общей ножкой пучка Гиса), войдя в желудочек но межжелудочковой перегородке, делится на две ветви (правую и левую ножки пучка Гиса), одна из которых идет к правому, а другая к левому желудочку сердца.
Конечные разветвления проводящей системы представлены широко распространенной, расположенной под эндокардом сетью волокон Пуркине, которые анастомозируют с мышечными волокнами миокарда. По разветвлениям проводящей системы возбуждение доходит до всей массы сердечной мышцы, вызывая ее сокращение.
Таким образом, импульс, вызывающий сокращение сердца, зарождается в синоатриальном узле, распространяется по сократительному миокарду правого и левого предсердий и доходит до атриовенгрикулярного узла. От этого узла по пучку Гиса импульс переходит к правому и левому желудочкам и вызывает их систолу.
Мышца сердца (миокард), подобно нервной ткани и скелетным мышцам, принадлежит к возбудимым тканям. Это означает, что волокна миокарда обладают потенциалом покоя, отвечают на надпороговые стимулы генерацией потенциалов действия и способны проводить эти потенциалы без затухания (бездекрементно). Межклеточные соединения в сердце (к которым относятся, в частности, так называемые вставочные диски, выявляемые при микроскопии) не препятствуют проведению возбуждения. Мышечная ткань предсердий и желудочков ведет себя как функциональный синцитий: возбуждение, возникающее в каком-либо из этих отделов, охватывает все без исключения невозбужденные волокна. Благодаря этой особенности сердце подчиняется закону “все или ничего”:на раздражение оно либо отвечает возбуждением всех волокон, либо (если раздражитель подпороговый) не реагирует вовсе. Этим оно отличается от нервов и скелетных мышц, где каждая клетка возбуждается изолированно, и поэтому только в тех клетках, на которые наносят надпороговые раздражения, возникают потенциалы действия.
Автоматизм.Ритмические сокращения сердца возникают под действием импульсов, зарождающихся в нем самом. Если изолированное сердце поместить в соответствующие условия, оно будет продолжать биться с постоянной частотой. Это свойство называется автоматизмом. В норме ритмические импульсы генерируются только специализированными клетками водителя ритма (пейсмекера) и проводящей системы сердца.
Клетки, способные к автоматической генерации потенциала действия, образуют узлы автоматии (водители ритма, или пейсмекеры).
Выделяют три узла автоматии: 1) синоатриальный узел, расположенный в районе венозного входа в правом предсердии (узел Кис-Фляка). Именно этот узел является реальным водителем ритма в норме.
2) Атриовентрикулярный узел (Ашоффа-Тавара), который расположен на границе правого и левого предсердий и между правым предсердием и правым желудочком. Этот узел состоит из трех частей: верхней, средней и нижней.
3) Волокна Пуркинье — это конечная часть пучка Гиса, миоциты которой расположены в толще миокарда желудочков. Они являются водителями 3-го порядка, их спонтанный ритм — самый низкий, поэтому в норме являются лишь ведомыми, участвуют в процессе проведения возбуждения по миокарду.
Синоатриальный узел представляет собой соединительнотканный остов, в котором расположены специализированные мышечные клетки — в основном так называемые Р-клетки, собранные в агрегаты. Каждая из клеток этого узла способна к автоматии — благодаря высокой проницаемости для ионов натрия, и способна генерировать медленную диастолическую деполяризацию. До сих пор остается неясной причина, порождающая высокую проницаемость для ионов натрия и те особенности электрической активности, которые и приводят к генерации спонтанного потенциала действия.
Единый пейсмекерный ритм строится на основе интегративного взаимодействия всех моментов гетерогенного пейсмекера.
Роль водителя ритма первого порядка — синоатриального узла — огромна. Все регулирующие воздействия, меняющие ритм сердечной деятельности, оказывают свое влияние на сердце посредством воздействия на водитель ритма первого порядка. Если этот водитель выходит из работы, то ни симпатическая, ни парасимпатическая системы сами по себе не могут запустить деятельность сердца. В случае, когда синоатриальный узел повреждается ипри этом человеку успевают оказать квалифицированную медицинскую помощь, больному вживляют стимулятор, задающий самостоятельно ритм для работы сердца. Благодаря такому способу удалось сохранить жизнь многих пациентов.
Возбудимость клеток проводящей системы и рабочего миокарда имеет ту же биоэлектрическую природу, что и в поперечно-полосатых мышцах. Наличие заряда на мембране здесь также обеспечивается разностью концентраций ионов калия и натрия возле ее внешней и внутренней поверхности и избирательной проницаемостью мембраны для этих ионов. В покое мембрана кардиомиоцитов проницаема для ионов калия и почти непроницаема для ионов натрия. В результате диффузии ионы калия выходят из клетки и создают положительный заряд на ее поверхности. Внутренняя сторона мембраны становится электроотрицательной по отношению к наружной.
В клетках атипического миокарда, обладающих автоматией, мембранный потенциал способен спонтанно уменьшаться до критического уровня, что приводит к генерации потенциала действия. В норме ритм сердечных сокращений задается всего несколькими наиболее возбудимыми клетками синоатриального узла, которые называются истинными водителями ритма, или пейсмекерными клетками. В этих клетках во время диастолы мембранный потенциал, достигнув максимального значения, соответствующего величине потенциала покоя (60—70 мВ), начинает постепенно снижаться. Этот процесс называют медленной спонтанной диастолической деполяризацией. Она продолжается до того момента, когда мембранный потенциал достигает критического уровня (40—50 мВ), после чего возникает потенциал действия.
Частота возбуждения пейсмекерных клеток у человека составляет в покое 70—80 в минуту при амплитуде потенциала действия 70—80 мВ. Во всех остальных клетках проводящей системы потенциал действия в норме возникает под влиянием возбуждения, приходящего из синоатриального узла. Такие клетки называют латентными водителями ритма. Потенциал действия в них возникает раньше, чем их собственная медленная спонтанная диастолическая деполяризация достигает критического уровня. Латентные водители ритма принимают на себя ведущую функцию только при условии разобщения с синоатриальным узлом. Частота спонтанной деполяризации таких клеток у человека составляет 30—40 в минуту.
Спонтанная медленная диастолическая деполяризация обусловлена совокупностью ионных процессов, связанных с функциями плазматических мембран. Среди них ведущую роль играют медленное уменьшение калиевой и повышение натриевой и кальциевой проводимости мембраны во время диастолы, параллельно чему происходит падение активности электрогенного натриевого насоса. К началу диастолы проницаемость мембраны для калия на короткое время повышается, и мембранный потенциал покоя приближается к равновесному калиевому потенциалу, достигая максимального диастолического значения. Затем проницаемость мембраны для калия уменьшается, что и приводит к медленному снижению мембранного потенциала до критического уровня. Одновременное увеличение проницаемости мембраны для натрия и кальция приводит к поступлению этих ионов в клетку, что также способствует возникновению потенциала действия. Снижение активности электрогенного насоса дополнительно уменьшает выход натрия из клетки и, тем самым, облегчает деполяризацию мембраны и возникновение возбуждения.
Ионные механизмы возникновения потенциала действия кардиомиоцитов. Соотношения возбуждения, возбудимости и сократимости в различные фазы кардиоцикла. Экстрасистолы, механизм формирования компенсаторной паузы.
Клетки миокарда обладают возбудимостью, но им не присуща автоматия. В период диастолы мембранный потенциал покоя этих клеток стабилен, и его величина выше (80—90 мВ), чем в клетках водителей ритма. Потенциал действия в этих клетках возникает под влиянием возбуждения клеток водителей ритма, которое достигает кардиомиоцитов, вызывая деполяризацию их мембран.
Конечная реполяризация в клетках миокарда обусловлена постепенным уменьшением проницаемости мембраны для кальция и повышением проницаемости для калия. В результате входящий ток кальция уменьшается, а выходящий ток калия возрастает, что обеспечивает быстрое восстановление мембранного потенциала покоя. Длительность потенциала действия кардиомиоцитов составляет 300—400 мс, что соответствует длительности сокращения миокарда.
Экстрасистолы.Внеочередные раздражения, действующие по окончании рефракторной фазы и ведущие к внеочередной систоле, т. е. к экстрасистоле, могут появляться в различных участках проводящей системы сердца. Если внеочередное раздражение возникает в синусном узле, то оно приводит к преждевременному возникновению внеочередного цикла сердечной деятельности, протекающему с обычной последовательностью сокращений предсердий и желудочков. Характерно для такой синусной экстрасистолы, что пауза после этого внеочередного сокращения сердца короче, чем пауза между обычными сокращениями.
После экстрасистолы желудочков наступает удлиненная пауза между нею и следующей (очередной) систолой желудочков. Эта удлиненная пауза, называемая компенсаторной паузой, зависит от того, что экстрасистола, так же как нормальная систола, оставляет за собой рефракторную фазу. Очередной импульс, в обычном порядке зарождающийся в синусном узле, приходит в желудочки тогда, когда они оказываются в рефракторной фазе после экстрасистолы. Этот импульс не может вызвать систолы желудочков, и она наступит лишь в ответ на следующий импульс.
Т.е. возбуждение по проводящей системе сердца в 5 раз быстрее скорости распространения возбуждения по рабочему миокарду.
Напомню, что проведением возбуждения по нервным волокнам составляет 0,5 – 120 м[Б47] ·с -1 .
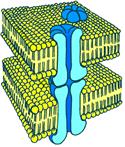
Рис[Б50] . 810132328. Нексус. Объяснение в тексте
Нексусы, как правило, соединяют группы синхронно функционирующих клеток, образующих функциональный синцитий.
Как устроены нексусы? В каждой из двух соседних клеточных мембран находятся регулярно распределенные коннексоны, пронизывающие всю толщу мембраны. Они расположены так, что в месте контакта клеток находятся друг против друга и их просветы оказываются на одной линии. У образованных таким образом каналов крупные внутренние диаметры и, значит, высокая проводимость для ионов. Через них могут приходить даже относительно крупные молекулы с молекулярной массой до 1000 (около 1,5 нм в поперечнике). Коннексон состоит из субъединиц числом до шести с молекулярной массой примерно 25000 каждая.
Но главное через нексусы свободно проходят ионы.
Принцип передачи возбуждения через нексус показан на рис[Б51] . 810132331.
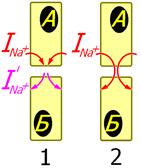
Рис[Б52] . 810132331. Натриевые токи (INa+) при передаче возбуждения в химическом синапсе (1), электрическом синапсе или между миокардиоцитами (2).
Принцип передачи возбуждения между миокардиоцитами аналогичен принципу передачи возбуждения в электрическом синапсе и существенно отличается от принципа передачи возбуждения в химическом синапсе.
При возбуждении клетки А натриевый ток (INa+) входит в нее через открытые потенциалуправляемые натриевые каналы. При этом часть тока входит через участок мембранного контакта в клетку Б, вызывая ее деполяризацию. Уровень деполяризации клетки Б гораздо ниже, однако он может оказаться выше критического уровня деполяризации мембраны клетки Б и в ней генерируется потенциал действия.
В химическом синапсе входящие натриевые токи клетки А прерываются на пресинаптической мембране. Деполяризующие входящие натриевые токи на постсинаптической мембране клетки Б возникают при активации медиаторами рецепторуправляемых натриевых каналов. При достижении критического уровня деполяризации мембраны клетки Б открываются потенциалуправляемые натриевые каналы.
Таким образом, деполяризующие токи в клетке Б в химическом синапсе образуются в самой клетке Б, а в электрическом синапсе и при контакте миокардиоцитов деполяризующие токи в клетку Б входят из клетки А.
Для [Б53] миокарда важна регулируемость щелевых контактов. Каналы миокардиоцитов закрываются при снижении pH или повышении концентрации Са 2+ . Это неизбежно происходит в случае повреждения клеток или глубокою нарушения обмена. За счет такого механизма пораженные места изолируются от остальной части функционального синцития, и распространение патологии ограничивается (например, при инфаркте миокарда).
В миокарде, как функциональном синцитии, возбуждение одной точки миокарда неизбежно вызывает возбуждение всех остальных областей миокарда. Это означает, что сердечная мышца отвечает на возбуждение по правилу "все или ничего", а градуальная зависимость, типичная для скелетной мышцы, здесь при нормальных условиях не наблюдается[Б54] .
Благодаря проводящей системе сердца волна возбуждения синхронно возбуждает рабочие миоциты.
Однако в атриовентрикулярном узле волна возбуждения на участке длиной 1,5‑2 мм задерживается, бежит с малой скоростью (2-5 см/с). Тем самым обеспечивается так называемая атриовентрикулярная задержка, благодаря которой возбуждение желудочков и их сокращение начинается через 100 мс после начала сокращения предсердий, а не раньше. Эта зона расположена в верхней части атриовентрикулярного узла.

Полагают, что снижение скорости проведения в этой зоне обусловлено особенностями контакта миоцитов - ход волокон перпендикулярен направлению волны возбуждения, что и обусловливает замедление ее движения по этому участку.
Важно отметить, что проведение волны возбуждения через атриовентрикулярный узел осуществляется лишь в том случае, если одновременно возбуждаются несколько миоцитов этого узла. Это защитный механизм от возникновения аритмий и появление эктопических очагов возбуждения.
Как правило, вначале возбуждается правое предсердие, а затем - с небольшим интервалом - левое предсердие.
| [17] |
| Типы (механизмы) проведения возбуждения по миокарду: 1. С участием специализированной проводящей системы сердца. 2. Передачей возбуждения от одного рабочего миокардиоцита к другому. |
| Скорость проведения возбуждения по миокарду: · предсердий составляет порядка 1 м/с · желудочков - 0,8 м/с · по проводящей системе - до 4-5 м/с |
Т.е. возбуждение по проводящей системе сердца в 5 раз быстрее скорости распространения возбуждения по рабочему миокарду.
Напомню, что проведением возбуждения по нервным волокнам составляет 0,5 – 120 м[Б47] ·с -1 .
Рис[Б50] . 810132328. Нексус. Объяснение в тексте
Нексусы, как правило, соединяют группы синхронно функционирующих клеток, образующих функциональный синцитий.
Как устроены нексусы? В каждой из двух соседних клеточных мембран находятся регулярно распределенные коннексоны, пронизывающие всю толщу мембраны. Они расположены так, что в месте контакта клеток находятся друг против друга и их просветы оказываются на одной линии. У образованных таким образом каналов крупные внутренние диаметры и, значит, высокая проводимость для ионов. Через них могут приходить даже относительно крупные молекулы с молекулярной массой до 1000 (около 1,5 нм в поперечнике). Коннексон состоит из субъединиц числом до шести с молекулярной массой примерно 25000 каждая.
Но главное через нексусы свободно проходят ионы.
Принцип передачи возбуждения через нексус показан на рис[Б51] . 810132331.
Рис[Б52] . 810132331. Натриевые токи (INa+) при передаче возбуждения в химическом синапсе (1), электрическом синапсе или между миокардиоцитами (2).
Принцип передачи возбуждения между миокардиоцитами аналогичен принципу передачи возбуждения в электрическом синапсе и существенно отличается от принципа передачи возбуждения в химическом синапсе.
При возбуждении клетки А натриевый ток (INa+) входит в нее через открытые потенциалуправляемые натриевые каналы. При этом часть тока входит через участок мембранного контакта в клетку Б, вызывая ее деполяризацию. Уровень деполяризации клетки Б гораздо ниже, однако он может оказаться выше критического уровня деполяризации мембраны клетки Б и в ней генерируется потенциал действия.
В химическом синапсе входящие натриевые токи клетки А прерываются на пресинаптической мембране. Деполяризующие входящие натриевые токи на постсинаптической мембране клетки Б возникают при активации медиаторами рецепторуправляемых натриевых каналов. При достижении критического уровня деполяризации мембраны клетки Б открываются потенциалуправляемые натриевые каналы.
Таким образом, деполяризующие токи в клетке Б в химическом синапсе образуются в самой клетке Б, а в электрическом синапсе и при контакте миокардиоцитов деполяризующие токи в клетку Б входят из клетки А.
Для [Б53] миокарда важна регулируемость щелевых контактов. Каналы миокардиоцитов закрываются при снижении pH или повышении концентрации Са 2+ . Это неизбежно происходит в случае повреждения клеток или глубокою нарушения обмена. За счет такого механизма пораженные места изолируются от остальной части функционального синцития, и распространение патологии ограничивается (например, при инфаркте миокарда).
В миокарде, как функциональном синцитии, возбуждение одной точки миокарда неизбежно вызывает возбуждение всех остальных областей миокарда. Это означает, что сердечная мышца отвечает на возбуждение по правилу "все или ничего", а градуальная зависимость, типичная для скелетной мышцы, здесь при нормальных условиях не наблюдается[Б54] .
Благодаря проводящей системе сердца волна возбуждения синхронно возбуждает рабочие миоциты.
Однако в атриовентрикулярном узле волна возбуждения на участке длиной 1,5‑2 мм задерживается, бежит с малой скоростью (2-5 см/с). Тем самым обеспечивается так называемая атриовентрикулярная задержка, благодаря которой возбуждение желудочков и их сокращение начинается через 100 мс после начала сокращения предсердий, а не раньше. Эта зона расположена в верхней части атриовентрикулярного узла.
Полагают, что снижение скорости проведения в этой зоне обусловлено особенностями контакта миоцитов - ход волокон перпендикулярен направлению волны возбуждения, что и обусловливает замедление ее движения по этому участку.
Важно отметить, что проведение волны возбуждения через атриовентрикулярный узел осуществляется лишь в том случае, если одновременно возбуждаются несколько миоцитов этого узла. Это защитный механизм от возникновения аритмий и появление эктопических очагов возбуждения.
Как правило, вначале возбуждается правое предсердие, а затем - с небольшим интервалом - левое предсердие.
АТМВ миокарда образуют так называемую проводящую систему. Она представляет собой совокупность узлов и пучков атипичной мышечной ткани, функцией которой является генерация ПД, служащих стимулами для ТМВ, то есть задание определенного ритма сердечных сокращений. Строение проводящей системы обеспечивает строго согласованное и последовательное возбуждение и сокращение различных отделов сердца.

В норме водителем ритма является синоатриальный узел, расположенный в стенке правого предсердия в месте впадения в него верхней полой вены. Частота разрядов СА в покое составляет около 70 1/мин. От этого узла возбуждение вначале распространяется по рабочему миокарду предсердий (со скоростью порядка 1 м/с).
Единственный путь, по которому возбуждение может пройти к желудочкам, образует атриовентрикулярный узел (АВ), лежащий в предсердно-желудочковой перегородке (остальная часть атриовентрикулярного соединения образована невозбудимой соединительной тканью). В АВ узле скорость проведения значительно падает (в 20-50 раз; 0,02-0,05 м/с) за счет снижения диаметра волокон АВ-узла и поперечного их расположения Передача возбуждения - через боковые щелевые контакты. Это приводит к тому, что возбуждение "задерживается" в АВ-узле (АВ-задержка необходима для полного перехода крови из предсердий в желудочки во время сокращения предсердий).
Далее возбуждение распространяется по пучку Гиса, ножкам пучка Гиса и волокнам Пуркинье к верхушке сердца со все возрастающей (до 4-5 м/с) скоростью (увеличение диаметра АТМВ), а затем переходит на рабочие волокна миокарда, по которым распространяется в обратном направлении – от верхушки сердца к основанию. За волной возбуждения следует сокращение ТМВ миокарда.
Следует отметить, что при нарушении функции водителя ритма первого порядка (СА-узла) его роль могут выполнять другие отделы проводящей системы (им также присущ автоматизм), но направление распространения возбуждения по сердцу при этом будет нарушено (да и собственная частота генерации ПД у этих отделов проводящей системы ниже – 40-60 – у АВ-узла, у других отделов – еще меньше). Это приводит к тяжелым нарушениям насосной функции сердца (пример – полная поперечная блокада).
5. Электрокардиография. Электрокардиограмма. Интегральный электрический вектор сердца.
Сложный характер распространения возбуждения по сердцу отображается в электрокардиограмме (ЭКГ), по форме которой можно судить о возбудимости и проводимости различных отделов сердца (но не о сократимости волокон миокарда!)
Если рассмотреть отдельное миокардиальное волокно, то в покое его наружная поверхность имеет положительный, а внутренняя – отрицательный потенциал. При возбуждении (ПД) возбужденный участок мембраны меняет свою полярность (см. схему). Возбужденное волокно можно рассматривать как диполь, обладающий определенным дипольным моментом. Векторная сумма дипольных моментов всех волокон миокарда называется интегральным электрическим вектором сердца (ИЭВС). Этот вектор в каждый момент времени направлен от наиболее возбужденного (электроотрицательного) к наименее возбужденному (электроположительному) участку сердца, и величина и направление его в ходе сердечного цикла многократно меняются.
Как известно, движущиеся заряды создают вокруг себя переменное электрическое поле, которое распространяется в пространстве. Поэтому работающее сердце также является источником электрического поля, которое можно зарегистрировать на поверхности тела. Для этого на различные точки поверхности тела накладывают отводящие электроды и регистрируют разность потенциалов между ними. Регистрирующий прибор (электрокардиограф) по сути представляет собой усилитель переменного тока и регистрирующее устройство (самописец). Кривая, отображающая зависимость этой разности потенциалов от времени, называется электрокардиограммой. Она представляет собой периодическое (Т = 1/ЧСС) колебание сложной формы.
Величина разности потенциалов, регистрируемой между двумя электродами, находящимися на поверхности тела человека будет зависеть от величины интегрального электрического вектора и угла между направлением этого вектора и осью отведения (проведенной между этими электродами) (см. схему). Таким образом, ЭКГ представляет собой динамику во времени проекции ИЭВС на ось отведения.
Какова же форма ЭКГ?

По предсердиям возбуждение распространяется в направлении сверху вниз; это означает, что ИЭВС ориентирован по направлению к верхушке сердца. На ЭКГ регистрируется зубец Р, отображающий деполяризацию предсердий. Во время возбуждения всех отделов предсердий разность потенциалов временно исчезает, так как ПД всех предсердных клеток находятся в фазе плато. В это время возбуждение распространяется по проводящей системе желудочков, но общее количество возбужденных клеток при этом невелико и существенной разности потенциалов не возникает (сегмент PQ). Длительность этого сегмента несет информацию о величине атриовентрикулярной задержки. При переходе на рабочий миокард желудочков в сердце снова появляются значительные градиенты напряжения. Возбуждение желудочков начинается с деполяризации левой поверхности межжелудочковой перегородки (ИЭВС направлен к основанию сердца (начало комплекса QRS). Затем вектор быстро меняет направление на противоположное (к верхушке) = распространение возбуждения через стенку желудочков от эндокарда к эпикарду (регистрируется самый крупный зубец R); в последнюю очередь возбуждается участок правого желудочка в области основания легочного ствола (ИЭВС направлен вправо и вверх). Когда желудочки полностью охвачены возбуждением, разность потенциалов временно исчезает (сегмент ST). Затем следует фаза реполяризации желудочков (зубец Т), в ходе этой фазы ИЭВС ориентирован влево (это связано с тем, что разные отделы миокарда желудочков реполяризуются с разной скоростью).
Выводы и заключение
Таким образом, в ходе сегодняшней лекции вы познакомились с особенностями распространения возбуждения по возбудимым мембранам (т.е. по мембранам мышечных и нервных волокон), основными принципами передачи возбуждения с одной возбудимой клетки на другую, а также с особенностями биоэлектрогенеза такого важного и сложного органа как сердце и с происхождением электрокардиограммы, имеющей огромное значение для диагностики многих видов патологии сердца. Естественно, что эти знания будут углублены и дополнены на практическом занятии, посвященном этой теме, а в дальнейшем - на кафедрах пропедевтики внутренних болезней, кардиологии и многих других, где вы познакомитесь с изменениями электрокардиограммы при нарушениях возбудимости и проводимости различных отделов миокарда.
Разработала заведующая кафедрой биологической и медицинской физики кандидат физико-математических наук доцент Новикова Н.Г.
Читайте также: