
Доказательство генетической роли нуклеиновых кислот кратко
Обновлено: 01.07.2024
Первый успех в молекулярной генетике был достигнут при изучении генетической трансформации у бактерий.
Трансформация в генетике, внесение в клетку генетической информации при помощи изолированной дезоксирибонуклеиновой кислоты (ДНК). Трансформация приводит к появлению у трансформированной клетки (трансформанта) и её потомства новых признаков, характерных для объекта — источника ДНК. Явление Трансформация было открыто в 1928 английским учёным Ф. Гриффитом, наблюдавшим наследуемое восстановление синтеза капсульного полисахарида у пневмококков при заражении мышей смесью убитых нагреванием капсулированных бактерий и клеток, лишённых капсулы. Организм мыши в этих экспериментах играл роль своеобразного детектора, так как приобретение капсульного полисахарида сообщало клеткам, лишённым капсулы, способность вызывать смертельный для животного инфекционный процесс (см. схему). В последующих экспериментах было установлено, что Трансформация имеет место и в том случае, когда вместо убитых клеток к лишённым капсулы пневмококкам добавляли экстракт из разрушенных капсулированных бактерий. В 1944 О. Эйвери с сотрудниками (США) установил, что фактором, обеспечивающим Трансформация, являются молекулы ДНК. Эта работа — первое исследование, доказавшее роль ДНК как носителя наследственной информации.
В 1944 американский учёный О. Т. Эйвери с сотрудниками обнаружил, что наследственные признаки одного штамма пневмококков могут быть переданы другому, генетически отличному штамму путём введения в его клетки дезоксирибонуклеиновой кислоты (ДНК), выделенной из первого штамма. Впоследствии подобная генетическая трансформация с помощью ДНК была осуществлена у других бактерий, а в последнее время — и у некоторых многоклеточных организмов (цветковые растения, насекомые). Т. о., было показано, что гены состоят из ДНК. Этот вывод был подтвержден опытами с ДНК-содержащими вирусами: для размножения вируса достаточно введения молекул вирусной ДНК в клетку восприимчивого хозяина; все др. компоненты вируса (белки, липиды) лишены инфекционных свойств и генетически инертны. Аналогичные опыты с вирусами, содержащими вместо ДНК рибонуклеиновую кислоту (РНК), показали, что у таких вирусов гены состоят из РНК. Выяснение генетической роли ДНК и РНК послужило мощным стимулом для изучения нуклеиновых кислот биохимическими, физико-химическими и рентгеноструктурными методами.
Составными частями нуклеиновых кислот являются нуклеотиды. Молекула нуклеотида состоит из пентозы, азотистого основания и фосфорной кислоты. В зависимости от типа сахара различают рибонуклеиновую кислоту (РНК; в её состав входит рибоза) и дезоксирибонуклеиновая кислота (ДНК; в её состав входит сахар дезоксирибоза, у которого на один атом кислорода меньше). В обоих типах нуклеиновых кислот содержатся четыре типа оснований: аденин (А), гуанин (Г), цитозин (Ц), тимин (Т; в РНК вместо него содержится урацил (У)). Первые два основания относятся к классу пуринов, остальные – к пиримидинам. Фосфорная кислота определяет кислотные свойства нуклеиновых кислот.
Выяснить структуру ДНК удалось в 1953 году английским ученым Д. Уотсону и Ф. Крику.
Они создали модель трехмерной двухспиральной структуры молекулы ДНК. Эта модель отвечала всем основным требованиям, необходимым генетическому материалу для выполнения биологических функций. Химическая структура гена, связанная с линейным расположением нуклеотидов в цепи, позволяла сохранять закодированную с помощью генетического кода наследственную информацию.
Благодаря принципу комплементарного связывания двух цепей молекула ДНК способна к ферментативному матричному аутокатализу — репликации, что позволяет точно копировать генетическую информацию и поддерживать наследственное постоянство при делении клеток (митоз, мейоз). На основе матричного синтеза генетическая информация может переписываться на посреднические молекулы иРНК (транскрипция). Информация о последовательностях нуклеотидов в иРНК переводится на рибосомах в последовательность аминокислот в полипептиде в процессе трансляции. Схематически это выглядит следующим образом: ДНК репликация 2ДНК транскрипция иРНК трансляция полипептид.
Модель двойной спирали ДНК и триплетности генетического кода позволила предсказать молекулярные механизмы возникновения спонтанных и индуцированных генных мутаций: во-первых, замена основания в одном кодоне приводит к изменению одной аминокислоты в белке; во-вторых, вставка или выпадение одного нуклеотида в одной цепи ДНК приводит к изменению всех последующих кодонов и отсутствию синтеза специфического белка, кодируемого соответствующим геном. Последствия такой мутации в гене могут быть губительными для клетки и целого организма. Например, в результате замены одного основания в гене, контролирующем синтез b-цепи гемоглобина, происходит замена одной аминокислоты — глутамина — в шестом положении на валин, что приводит к синтезу аномальной молекулы гемоглобина, изменению формы эритроцитов и к болезни (См. Серповидноклеточная анемия). Одних лишь гемоглобинопатийнасчитывается несколько десятков. На основе подобного анализа выяснена природа многих сотен молекулярных болезней человека и разработаны методы пренатальной молекулярно-генетической диагностики (см. Наследственные болезни).
После доказательства генетической роли нуклеиновых кислот и расшифровки структуры молекулы ДНК С. Бензер в экспериментах на бактериофаге Т4 показал, что наименьшими мутирующими элементами гена являются отдельные пары нуклеотидов, и кроссинговер может происходить между двумя парами нуклеотидов. Было окончательно постулировано, что ген представляет собой определенный участок ДНК, состоящий из нескольких тысяч пар нуклеотидов, способных мутировать и быть разделенными рекомбинацией, но функционально представляющий единое целое.
8. Функции нуклеиновых кислот в реализации генетической информации: репликация, транскрипция и трансляция. Методологическое значение принципа передачи генетической информации: ДНК —* РНК —* белок.
Транскрипция ДНК в ходе деления клеток начинается с разделения двух цепей, каждая из которых становится матрицей, синтезирующей нуклеотидную последовательность новых цепей. Хеликаза, топоизомераза и ДНК-связывающие белки расплетают ДНК, удерживают матрицу в разведённом состоянии и вращают молекулу ДНК. Правильность репликации обеспечивается точным соответствием комплементарных пар оснований. Репликация катализуется несколькими ДНК-полимеразами, а транскрипция – ферментом РНК-полимеразой. После репликации дочерние спирали закручиваются обратно уже без затрат энергии и каких-либо ферментов.
Репликация эукариот при такой же схеме длилась бы несколько месяцев (скорость движения репликативных вилок составляет всего микрометр в минуту). Поэтому в ДНК эукариот процесс начинается одновременно в сотнях и тысячах точек. Все хромосомы в клетке должны реплицироваться одновременно, и одновременно в клетке работают многие тысячи вилок.
Между репликацией и транскрипцией есть существенная разница: в первом случае копируется вся молекула ДНК, во втором, как правило, только отдельные гены. Минимальная длина и-РНК определяется длиной полипептидной цепи, для которой она предназначена. В идентификации последовательностей нуклеотидов, обозначающих начало и конец синтезирующих РНК генов, ещё много неясного.
Молекулы р-РНК и т-РНК образуются из более длинных предшественников – гетерогенных ядерных РНК (гя-РНК). Длина гя-РНК увеличена за счет нетранслирующихся интронов, которых в конечных РНК уже нет. Интроны удаляются при помощи малой ядерной РНК. мя-РНК комплементарна нуклеотидам на концах интронов – она временно соединяется с ними, стягивая интрон в петлю. Концы кодирующих фрагментов соединяются, после чего интрон благополучно удаляется из цепи.
Некоторые РНК-содержащие вирусы животных при помощи РНК-зависимой ДНК-полимеразы способны синтезировать ДНК, комплементарную по отношению к вирусной РНК. Она встраивается в геном эукариотической клетки, где может многие поколения оставаться в скрытом состоянии. При определённых условиях (например, воздействии канцерогенов) вирусные гены могут активироваться, и здоровые клетки превратятся в раковые.
Синтез белка (трансляция) является самым сложным из биосинтетических процессов: он требует очень большого количества ферментов и других специфических макромолекул, общее количество которых, видимо, доходит до трёхсот. Часть из них к тому же объединены в сложную трёхмерную структуру рибосом. Но несмотря на большую сложность синтез протекает с чрезвычайно высокой скоростью (десятки аминокислотных остатков в секунду). Процесс может замедляться и даже останавливаться ингибиторами-антибиотиками.
В пятидесятых годах XX века было установлено, что синтез белка происходит в рибонуклеопротеиновых частицах, называющихся рибосомами. Диаметр рибосомы бактерии E. coli составляет 18 нм, а их общее количество – десятки тысяч в клетке. Рибосомы эукариот несколько крупнее (21 нм). Сам процесс протекает в пять этапов.
1. Активация аминокислот. Каждая из 20 аминокислот белка соединяется ковалентными связями к определённой т-РНК, используя энергию АТФ. Реакция катализуется специализированными ферментами, требующими присутствия ионов магния.
2. Инициация белковой цепи. и-РНК, содержащая информацию о данном белке, связывается с малой частицей рибосомы и с инициирующей аминокислотой, прикреплённой к соответствующей т-РНК. т-РНК комплементарна с находящимся в составе и-РНК триплетом, сигнализирующим о начале белковой цепи.
3. Элонгация. Полипептидная цепь удлиняется за счёт последовательного присоединения аминокислот, каждая из которых доставляется к рибосоме и встраивается в определённое положение при помощи соответствующей т-РНК. В настоящее время генетический код полностью расшифрован, то есть всем аминокислотам поставлены в соответствие триплеты нуклеотидов. Элонгация осуществляется при помощи белков цитозоля (так называемые факторы элонгации).
4. Терминация. После завершения синтеза цепи, о чём сигнализирует ещё один специальный кодон и-РНК, полипептид высвобождается из рибосомы.
5. Сворачивание и процессинг. Чтобы принять обычную форму, белок должен свернуться, образуя при этом определённую пространственную конфигурацию. До или после сворачивания полипептид может претерпевать процессинг, осуществляющийся ферментами и заключающийся в удалении лишних аминокислот, присоединении фосфатных, метильных и других групп и т. п.
Синтез белка требует больших затрат энергии – 24,2 ккал/моль. После окончания синтеза белок при помощи специального полипептидного лидера доставляется к месту своего назначения.
роли ДНК. Еще одним шагом в доказательстве генетической роли нуклеиновой кислоты является открытие правила эквивалентности, согласно которому в ДНК, выделенных из организмов различных видов, соотношение пуриновых и пиримидиновых оснований всегда одно и то же и составляет 1:1 (правило Э.Чаргаффа).

В 1953 г. Дж. Уотсон и Ф. Крик на основании результатов рентгеноструктурного анализа и биохимических данных предложили пространственную модель структуры ДНК, объясняющую все ее свойства. Согласно предложенной модели молекула ДНК состоит из двух комплементарных (соответствующих) нитей. М. Мезельсон и Ф. Сталь доказали полуконсервативный механизм репликации (удвоения) ДНК.
Выяснение структуры и функции нуклеиновых кислот позволило понять, каким образом живые организмы воспроизводят себя и как осуществляются кодирование генетической информации, ее хранение и реализация, необходимые для протекания всех жизненных процессов.
К настоящему времени существенным образом обогащены знания о структуре и функции ДНК, значительно расширены возможности для исследований. Было обнаружено, что ДНК может повреждаться и может восстанавливаться, что молекулы ДНК могут обмениваться друг с другом частями, закручиваться и раскручиваться. Было показано, что ДНК служит матрицей для синтеза РНК, а также сама способна синтезироваться в процессе обратной транскрипции с РНК. ДНК функционирует не только в ядре, но и в митохондриях. В настоящее время исследователи способны определять последовательность нуклеиновых оснований в ДНК и осуществлять ее синтез.
роли ДНК. Еще одним шагом в доказательстве генетической роли нуклеиновой кислоты является открытие правила эквивалентности, согласно которому в ДНК, выделенных из организмов различных видов, соотношение пуриновых и пиримидиновых оснований всегда одно и то же и составляет 1:1 (правило Э.Чаргаффа).
В 1953 г. Дж. Уотсон и Ф. Крик на основании результатов рентгеноструктурного анализа и биохимических данных предложили пространственную модель структуры ДНК, объясняющую все ее свойства. Согласно предложенной модели молекула ДНК состоит из двух комплементарных (соответствующих) нитей. М. Мезельсон и Ф. Сталь доказали полуконсервативный механизм репликации (удвоения) ДНК.
Выяснение структуры и функции нуклеиновых кислот позволило понять, каким образом живые организмы воспроизводят себя и как осуществляются кодирование генетической информации, ее хранение и реализация, необходимые для протекания всех жизненных процессов.
К настоящему времени существенным образом обогащены знания о структуре и функции ДНК, значительно расширены возможности для исследований. Было обнаружено, что ДНК может повреждаться и может восстанавливаться, что молекулы ДНК могут обмениваться друг с другом частями, закручиваться и раскручиваться. Было показано, что ДНК служит матрицей для синтеза РНК, а также сама способна синтезироваться в процессе обратной транскрипции с РНК. ДНК функционирует не только в ядре, но и в митохондриях. В настоящее время исследователи способны определять последовательность нуклеиновых оснований в ДНК и осуществлять ее синтез.
2. Строение и сравнительная характеристика молекул ДНК, РНК.
3. Репликация ДНК, генетический код, реализация генетической информации.
4. Строение гена и его свойства.
Генетическая роль нуклеиновых кислот.
Молекулярная биология стала активно развиваться с конца 19 века. Долгое время оставалось неизвестным, что представляет собой вещество способное к саморепликации, мутациям и фенотипическому проявлению.
Нуклеиновые кислоты впервые описал швейцарский биохимик И.Ф. Лишнер (Липшер, Мишер) в 1869 году. Обнаруженное в ядре клетки вещество он первоначально назвал нуклеон от лат. Nucleus - ядро. До начала XX века предполагалось, что наследственную информацию передают белки. Благодаря совместной работе физиков, химиков, биологов, генетиков в XX веке началось активное изучение биологически активных молекул, входящих в состав клетки и оказалось, что наследственная информация зашифрована (закодирована) в молекуле ДНК. Впервые это доказал бактериолог Ф. Гриффит в 1928 году при изучении бактерий пневмококков, которые вызывают пневмонию. В 1927 году русский биолог Н.К. Кольцов сформулировал принцип редупликации наследственных структур. В 1940 году американские учёные Джордж Бидл и Эдвард Татум доказали, что в генах закодирована информация о структуре белка. В 1953 году Френсис Крик и Джеймс Уотсон предложили модель строения ДНК. В 1969 году индийский учёный Г. Корана впервые осуществил искусственный синтез гена. Фундаментальные открытия в области молекулярной биологии позволили понять, как работают механизмы наследственности на молекулярном уровне, с которого начинается формирование любых фенотипических признаков человека.
Генетическая роль нуклеиновых кислот (ДНК, РНК) стала известна, как только были открыты законы наследственности. Их функция состоит в хранении, передачи и воспроизводстве генетической информации и регуляции жизнедеятельности клетки.
Строение и сравнительная характеристика молекул ДНК и РНК.
Строение молекул ДНК, РНК.
Нуклеиновые кислоты (ДНК, РНК) представляют собой биополимеры (макромолекулы, полинуклеотиды), мономерами которых являются нуклеотиды. Нуклеиновые кислоты — это очень длинные молекулы, которые могут содержать до нескольких миллиардов нуклеотидов.
Нуклеотид состоит из трёх частей: 1) азотистое основание (аденин, гуанин, цитозин, тимин, уроцил), 2) углевод (рибоза, дезоксирибоза), 3) остаток фосфорной кислоты.
1) Азотистые основания бывают двух типов:
1. Пуриновые (аденин, гуанин)
2. пиримидиновые (тимин, цитозин, урацил)
В состав молекулы ДНК входят только четыре типа азотистых оснований: аденин, гуанин, цитозин, тимин. В молекулу РНК также входят четыре типа азотистых оснований: аденин, гуанин, цитозин, но вместо тимина содержится уроцил.
2) Углевод, входящий в состав нуклеотида содержит пять атомов углерода т.е. относится к классу пентоз. В состав молекулы ДНК входит дезоксирибоза, а в состав молекулы РНК входит рибоза.
Нуклеиновые кислоты являются кислотами потому, что в их состав входит остаток фосфорной кислоты.
Соединение нуклеотидов в нити происходит посредством ковалентных связей через углерод одного нуклеотида и остаток фосфорной кислоты соседнего нуклеотида. Каждый нуклеотид обозначается заглавной буквой в зависимости от содержащегося в нём азотистого основания: Л - адениловый, Г - гуаниловый, Ц - цитидиловый,
Т - тимидиловый, У -урациловый.

1.Первое правило:
2.Второе правило: А+Г=Ц+Т
3.Третье правило: А+Ц=Г+Т
Согласно модели, предложенной в 1953 году Джеймсом Уотсоном и Френсисом Криком, молекула ДНК представляет собой две параллельные неразветвленные полинуклеотидные цепочки, закрученные вокруг общей оси в двойную спираль. Цепочки в молекуле расположены антипараллельно т.е. навстречу друг другу. Они удерживаются рядом за счёт водородных связей. Водородные связи возникают между комплементарными (подходящие друг к другу как ключ к замку) азотистыми основаниями. Аденин комплементарен тимину (А = Т) две водородные связи, гуанин комплементарен цитозину (Г = Ц) три водородные связи. Комплементарность двух нитей ДНК приводит к тому, что число пуриновых оснований равно числу пиримидиновых А=Т, Г=Ц или (А+Г) / (Т+Ц) = 1. Молекулы ДНК бывают либо линейными, либо замкнутыми в кольцо. В ядре клетки находятся линейные ДНК (ядерные), а в митохондриях (митохондриальные ДНК) и пластидах (хлоропласты, лейкопласты) кольцевые ДНК. Размеры ДНК могут меняться в широких пределах - от нескольких пар нуклеотидов до миллиардов пар нуклеотидов. Средняя длина молекулы ядерной ДНК около 4 см, а общая длина всех молекул ДНК в клетках человеческого организма около 176 см.
Генетический код.
Для молекулы наследственности, которой является ДНК, свойственно не только самоудвоение (репликация), но и кодирование информации с помощь определённой последовательности нуклеотидов. Известно, что ДНК состоит из четырёх видов нуклеотидов, то есть информация в ДНК записывается 4 буквами (А, Т, Г, Ц). Математические расчёты показывают, что
Таким образом, комбинации из 3 нуклеотидов будет достаточно, чтобы закодировать 20 аминокислот. Из 64 возможных триплетов 61 триплет кодирует 20 незаменимых аминокислот, обнаруженных в составе клеточных белков, а 3 триплета являются стоп- сигналами или терминаторами, которые прекращают считывание информации.
Сочетания из трёх нуклеотидов, кодирующие определённые аминокислоты, называются кодом ДНК, или генетическим кодом. В настоящее время генетический код полностью расшифрован, то есть известно, какие триплетные сочетания нуклеотидов кодируют 20 аминокислот. Пользуясь комбинацией, состоящей из трёх нуклеотидов, можно закодировать больше аминокислот, чем необходимо для кодирования 20 аминокислот. Оказалось, что каждая аминокислота может кодироваться несколькими триплетами, кроме метионина и триптофана. Аминокислоты входящие в состав природных белков могут относится к разным группам, заменимые кислоты (З), незаменимые (НЗ).
Генетический код - это система записи генетической информации в ДНК в виде определённой последовательности нуклеотидов (или способ записи последовательности аминокислот в белке с помощью нуклеотидов).
Генетический код обладает несколькими свойствами (7 свойств).
Таблица. Аминокислоты входящие в состав природных белков.
| № | Аминокислота | Назв | Гр | Кодирующие триплеты (кодоны) | ||||
| Аланин | Ала | ГЦУ | ГЦЦ | ГЦА | ГЦГ | |||
| Аргинин | Арг | ЦГУ | ЦГЦ | ЦГА | ЦГГ | АГА | АГГ | |
| Аспарагин | Асн | ААУ | ААЦ | |||||
| Аспарагиновая кислота | Асп | ГАУ | ГАЦ | |||||
| Валин | Вал | нз | ГУУ | ГУЦ | ГУА | ГУГ | ||
| Гистидин | Гис | ЦАУ | ЦАЦ | |||||
| Глицин | Гли | ГГУ | ГГЦ | ГГА | ГГГ | |||
| Глутамин | Глн | ЦАА | ЦАГ | |||||
| Глутаминовая кислота | Глу | ГАА | ГАГ | |||||
| Изолейцин | Иле | нз | АУУ | АУЦ | АУА | |||
| Лейцин | Лей | нз | ЦУУ | ЦУЦ | ЦУА | ЦУГ | УУА | УУГ |
| Лизин | Лиз | нз | ААА | ААГ | ||||
| Метионин | Мет | нз | АУГ | |||||
| Пролин | Про | ЦЦУ | ЦЦЦ | ЦЦА | ЦЦГ | |||
| Серин | Сер | УЦУ | УЦЦ | УЦА | УЦГ | АГУ | АГЦ | |
| Тирозин | Тир | УАУ | УАЦ | |||||
| Треонин | Тре | нз | АЦУ | АЦЦ | АЦА | АЦГ | ||
| Триптофан | Три | нз | УГГ | |||||
| Фенилаланин | Фен | нз | УУУ | УУЦ | ||||
| Цистеин | Цис | УГУ | УГЦ | |||||
| Не кодирующие триплеты | УГА | УАГ | УАА |
Таблица. Генетический код.
В таблице приведён состав триплетов, которыми закодированы все 20 аминокислот (названия сокращены). Так как при синтезе полипептидной цепи молекулы белка информация считывается с и-РНК, то назван состав триплетов нуклеотидов и-РНК, а в скобках указаны комплементарные основания ДНК.
Правила пользования таблицей. Первый нуклеотид в триплете берётся из левого вертикального ряда. Второй нуклеотид берётся из верхнего горизонтального ряда. Третий нуклеотид берётся из правого вертикального ряда. Там, где пересекутся линии, идущие от трёх нуклеотидов, и находится искомая аминокислота.
| Первое | Второе основание | Третье основание | |||
| основание | У (А) | Ц(Г) | А(Т) | Г(Ц) | |
| У (А) | Фен | Сер | Тир | Цис | У (А) |
| Фен | Сер | Тир | Цис | Ц (Г ) | |
| Лей | Сер | - | - | А (Т) | |
| Лей | Сер | - | Три | Г (Ц) | |
| Ц (Г) | Лей | Про | Гис | Арг | У(А) |
| Лей | Про | Гис | Арг | Ц (Г) | |
| Лей | Про | Глн | Арг | А (Т) | |
| Лей | Про | Глн | Арг | Г (Ц) | |
| А (Т) | Иле | Тре | Асн | Сер | У (А) |
| Иле | Тре | Асн | Сер | Ц (Г) | |
| Иле | Тре | Лиз | Арг | А (Т) | |
| Мет | Тре | Лиз | Арг | Г (Ц) | |
| Г (Ц) | Вал | Ала | Асп | Гли | У (А) |
| Вал | Ала | Асп | Гли | Ц(Г) | |
| Вал | Ала | Глу | Гли | А(Т) | |
| Вал | Ала | Глу | Гли | Г (Ц) |
Лекция № 3
1. Генетическая роль нуклеиновых кислот.
2. Строение и сравнительная характеристика молекул ДНК, РНК.
3. Репликация ДНК, генетический код, реализация генетической информации.
4. Строение гена и его свойства.
Генетическая роль нуклеиновых кислот.
Молекулярная биология стала активно развиваться с конца 19 века. Долгое время оставалось неизвестным, что представляет собой вещество способное к саморепликации, мутациям и фенотипическому проявлению.
Нуклеиновые кислоты впервые описал швейцарский биохимик И.Ф. Лишнер (Липшер, Мишер) в 1869 году. Обнаруженное в ядре клетки вещество он первоначально назвал нуклеон от лат. Nucleus - ядро. До начала XX века предполагалось, что наследственную информацию передают белки. Благодаря совместной работе физиков, химиков, биологов, генетиков в XX веке началось активное изучение биологически активных молекул, входящих в состав клетки и оказалось, что наследственная информация зашифрована (закодирована) в молекуле ДНК. Впервые это доказал бактериолог Ф. Гриффит в 1928 году при изучении бактерий пневмококков, которые вызывают пневмонию. В 1927 году русский биолог Н.К. Кольцов сформулировал принцип редупликации наследственных структур. В 1940 году американские учёные Джордж Бидл и Эдвард Татум доказали, что в генах закодирована информация о структуре белка. В 1953 году Френсис Крик и Джеймс Уотсон предложили модель строения ДНК. В 1969 году индийский учёный Г. Корана впервые осуществил искусственный синтез гена. Фундаментальные открытия в области молекулярной биологии позволили понять, как работают механизмы наследственности на молекулярном уровне, с которого начинается формирование любых фенотипических признаков человека.
Генетическая роль нуклеиновых кислот (ДНК, РНК) стала известна, как только были открыты законы наследственности. Их функция состоит в хранении, передачи и воспроизводстве генетической информации и регуляции жизнедеятельности клетки.
'В 1869 г. швейцарский биохимик Ф. Мишер впервые описал вещество, содержащееся в ядрах клеток, и назвал его нуклеином, а позже оно было переименовано в нуклеиновые кислоты (от лат. Nucleus - ядро). К ним относятся дезоксирибонуклеиновая кислота - ДНК (в ее состав входит сахар дезоксирибоза) и рибонуклеиновая кислота - РНК (входит сахар рибоза).
В 1928 г. бактериолог Ф. Гриффит изучал бескапсульные невирулентные пневмококки (не вызывающие заболевания) и вирулентные в полисахаридной капсуле (вызывающие воспаление легких) для получения вакцины против пневмококка. Он показал, что при инъекции мышам живых бескапсульных пневмококков мыши выживали, а при введении живых капсульных - погибали. При введении смеси убитых при нагревании капсульных и живых
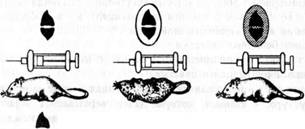


1 2 3
1- бескапсульные живые пневмококки; 2- капсульные живые пневмококки; 3-капсульные убитые нагреванием пневмококки
Рис.3.Схема опыта, демонстрирующего явление трансформации
бескапсульных пневмококков мыши погибали, из них удалось выделить живых капсульных пневмококков. Таким образом, способность образовывать капсулу перешла от убитого капсульного пневмококка к живому бескапсульному (рис. 3).
В 1944 г. О. Эвери с сотрудниками выяснили природу этого загадочного явления. Фактором, превращающим непатогенные (бескапсульные) в патогенные (капсульные) пневмококки, является ДНК, а само явление назвали трансформацией (от лат. transformatio - преобразование, превращение). Следовательно, трансформация - это преобразование признака у одного штамма бактерии в результате проникновения в нее ДНК другого штамма. Явление трансформации стало одним из основных доказательств того, что ДНК является носителем генетической (наследственной) информации.
Позже, в 1952 году Дж.Ледербергом и Н.Циндером была выявлена передача генетического материала от одного штамма бактерий к другому с помощью бактериофага, это было названо трансдукцией (от лат. Transduction- перемещение, передача) (рис. 4). U-образная трубка в нижней части разделена бактериальным фильтром. В одну половину были помещены штаммы сальмонеллы (S. typhi murium), не синтезирующие аминокислоту триптофан (Т-), а в другую- сальмонеллы, синтезирующие триптофан (Т+) и бактериофаги. После инкубации среди сальмонелл, не синтезирующих триптофан, были выделены бактерии Т+. Это объясняется тем, что бактериофаги проходили через бактериальный фильтр и переносили части ДНК от бактерии Т+ к бактериям Т-.
 |
Благодаря процессу транскрипции в клетке осуществляется передача информации от ДНК к белку по цепочке: ДНК → и-РНК → белок.
Генетический код - это система записи информации о последовательности расположения аминокислот в белках с помощью последовательности расположения нуклеотидов в ДНК и и-РНК. Участок молекулы ДНК, состоящий из трех нуклеотидов, называется триплетом или кодоном.
Свойства генетического кода:
1. Код триплетен - каждая из 20 аминокислот зашифрована последовательно расположенными тремя нуклеотидами. Из 4 нуклеотидов (так как существует 4 варианта азотистых оснований) можно создать 64 различные комбинации по 3 нуклеотида в каждом (4x4x4 = 64).
2. Код вырожден - каждая аминокислота шифруется более чем одним кодоном (от двух до шести), исключение составляют аминокислоты: метионин, который кодируется только триплетом АУГ и триптофан - УГГ.
3. Код специфичен - каждый кодон шифрует только одну аминокислоту.
4. Код универсален - один триплет кодирует одну и ту же аминокислоту у всех живых организмов.
5. Код неперекрываем - каждый нуклеотид входит лишь в какой-либо один триплет и переписывание информации происходит строго потриплетно.
6. Триплеты УАА, УАГ, УГА обозначают прекращение синтеза одной полипептидной цепи, так как к ним нет аминокислот. Они находятся в конце каждого гена.
Читайте также: