
Кто предположительно предок покрытосеменных кратко
Обновлено: 05.07.2024
Покрытосеменные – самый крупный тип растений, к которому относится более половины всех известных видов, характеризуются рядом четких, резко отграничивающих их признаков. Наиболее характерно для них наличие пестика, образованного одним или несколькими плодолистиками (макро- и мегаспорофиллами), сросшимися своими краями, так что в нижней части пестика образуется замкнутое полое вместилище – завязь, в которой развиваются семяпочки (макро- и мегаспорангии). После оплодотворения завязь разрастается в плод, внутри которого находятся развившиеся из семяпочек семена (или одно семя). Кроме того для покрытосеменных характерны: восьмиядерный, или производный из него, зародышевый мешок, двойное оплодотворение, триплоидный эндосперм, образующийся только после оплодотворения, рыльце у пестика, улавливающее пыльцу, и для подавляющего большинства – более или менее типичный цветок с околоцветником. Из анатомических признаков для покрытосеменных характерно наличие настоящих сосудов (трахей), тогда как у голосеменных развиты только трахеиды, а сосуды встречаются крайне редко. Ввиду большого количества общих признаков можно предположить монофилетическое происхождение покрытосеменных от какой-то более примитивной группы голосеменных.
Наиболее ранние и очень отрывочные ископаемые остатки покрытосеменных (пыльца, древесина) известны из юрского геологического периода. Из нижнемеловых отложений известны тоже немногочисленные достоверные остатки покрытосеменных, а в отложениях середины мелового периода они встречаются сразу в больших количествах и в значительном разнообразии форм, которые все принадлежат ко многим различным ныне живущим семействам и даже родам. В качестве предполагаемых предков покрытосеменных указывали различные группы нижестоящих в системе растений – кейтониевые, семенные папоротники, беннеттиты, гнетовые.
Теория происхождения покрытосеменных из гнетовых предполагает, что наиболее примитивныеы покрытосеменные имели мелкие однополые цветки без околоцветника или с невзрачным околоцветником. Но по ряду соображений в настоящее время более примитивными цветками считают крупные, обоеполые цветки.
Поэтому можно предположить, что предками современных покрытосеменных были какие-то вымершие, очень примитивные голосеменные с обоеполыми цветками типа шишки (стробилами), в которых на длинном цветоложе (оси) были спирально расположены свободные (несросшиеся друг с другом) листочки однородного околоцветника, микроспорофиллы (тычинки) и мегоспорофиллы (плодолистики). В системе голосеменных эта группа должна была стоять где-то между семенными папоротниками и уже более специализированными беннеттитами и саговниками.
Покрытосемянность несомненно представляла большое преимущество в смысле защиты семяпочек и развивающихся семян от всяких неблагоприятных внешних воздействий и в первую очередь от сухости воздуха.
Происхождение покрытосеменных
Но одной покрытосемянностью все же трудно объяснить быстрое мощное развитие покрытосеменных и вытеснение ими господствовавших ранее на земле архегониальных растений.
Русский ботаник М. И. Голенкин высказал (в 1927 г.) интересную гипотезу о причинах победы покрытосеменных в борьбе за существование. Он предполагает, что в середине мелового периода по каким-то общим космогоническим причинам по всей Земле произошло резкое изменение освещения и влажности воздуха. Густые облака, окутывавшие ранее постоянно Землю, рассеялись и дали доступ ярким солнечным лучам, в связи с чем резко увеличилась сухость воздуха. Громадное большинство высших архегониальных растений того времени, не приспособленных и не сумевших приспособиться к яркому освещению и сухости воздуха, начало вымирать или резко сократило области своего распространения (кроме хвойных, наиболее ксерофитных).
Наоборот, покрытосеменные, имевшие до того очень ограниченное распространение и представление небольшим числом форм, выработали способность хорошо переносить яркий солнечный свет и сухость воздуха. Это обстоятельство, а также чрезвычайная эволюционная пластичность их, способность к возникновению разнообразнейших приспособлений к различным внешним условиям и обусловили быстрое победное распространение покрытосеменных по всей Земле и вытеснение ранее господствовавших групп высших архегониальных растений.
Цветковые, или покрытосеменные – сравнительно молодая группа высших растений, появившаяся в меловом периоде, стремительно размножившаяся в середине – конце мела и с тех пор занимающая господствующее положение на суше. Проблеме происхождения цветковых посвящено огромное количество публикаций, однако в этом вопросе по-прежнему много неясного. Общепризнано, что цветковые произошли от голосеменных.
Д. Д. Соколов и А. К. Тимонин подробно рассмотрели основные теории происхождения покрытосеменных. Авторы обращают внимание на парадоксальный факт: цветы покрытосеменных до такой степени разнообразны, что не существует ни одного признака или структурной особенности, которая была бы присуща всем цветкам без исключения. Поэтому и сформулировать, что такое цветок, весьма трудно. Тем не менее, ясно, что именно появление цветка было ключевым событием в происхождении покрытосеменных.
Впрочем, если не рассматривать мужские цветки, у остальных все-таки есть общая черта, отличающая цветок покрытосеменных от фруктификаций голосеменных. У цветковых плодолистики свернуты и образуют, во-первых, рыльце, улавливающее пыльцу, во вторых – замкнутую полость (завязь), в которой развиваются семязачатки. У некоторых голосеменных есть нечто похожее – замкнутые капсулы с семязачатками, однако у них нет завязи (пыльцу улавливают сами семязачатки при помощи длинных отростков, высовывающихся из узкого отверстия капсулы).
К тому же исходный листочек, несущий семязачатки (мегаспорофилл, у цветковых ему соответствует плодолистик) свернут у голосеменных нижней стороной внутрь, а у цветковых – наоборот, верхней стороной внутрь. Соответственно и семязачатки сидят на разных сторонах листочка. Эти различия представляются настолько радикальными, что большинство ботаников не видят возможности постепенного перехода из одного состояния в другое и предполагают сальтационное превращение (одним скачком). Это вполне возможно, если учесть, что развитие фруктификаций у растений управляется небольшим числом генов-регуляторов, мутации в которых действительно могут приводить к скачкообразным изменениям плана строения. Аналогичная ситуация наблюдается и у животных, план строения которых в основном определяется небольшим набором генов-регуляторов.
В начале XX века была популярна теория происхождения цветковых от беннеттитов – вымершей группы голосеменных, у которых имелись обоеполые фруктификации (стробилы) – в отличие от других голосеменных, имеющих однополые фруктификации. Из стробилов беннеттитов, действительно напоминающих цветы, ботаники и пытались вывести типичный для покрытосеменных обоеполый цветок. Но тут возникла проблема: у беннеттитов не обнаружилось никаких структур, из которых можно было бы вывести плодолистики. У них вообще нет мегаспорофиллов, а семязачатки сидят прямо на оси стробила. Проблема казалась неразрешимой.

Рис. 1. Лист беннеттитового Pterophyllum(поздний триас)
В 80-х годах XX века крупнейший российский палеоботаник С. В. Мейен выдвинул гипотезу, разрешающую это противоречие. Он предположил, что у каких-то беннеттитовых сальтационным путем произошла перестройка программы развития фруктификации. В результате ключевые элементы программы развития мужских элементов стробилы (микроспорофиллов, у цветковых это тычинки) стали реализовываться при развитии женских элементов.
Ничего невероятного в таком сценарии нет, исходя из того, что известно об общих принципах генетической регуляции развития. Гипотеза Мейена объясняет противоречие, казавшееся неразрешимым. Ее нельзя проверить классическими методами палеоботаники и морфологии (поскольку никаких переходных форм гипотеза Мейена не предполагает). Однако ее можно проверить методами сравнительно-генетического анализа.
Генетическая регуляция развития фруктификаций у семенных растений стала активно изучаться в начале 90-х годов XX века. Оказалось, что у всех цветковых главным регулятором развития цветка является ген LFY.
Эволюционные реконструкции, основанные на морфологии, подтверждали родство покрытосеменных и беннеттитовых (а также родственных беннеттитам гнетовых, которые дожили до наших дней). Однако реконструкции, основанные на молекулярных данных, показали иное. Они показали, что все дожившие до современности голосеменные представляют собой монофилетичную группу (т.е. группу, происходящую от одного общего предка и включающую всех без исключения потомков этого предка).
Это значит, что последний общий предок всех современных голосеменных, в том числе гнетовых (а стало быть и беннеттитов тоже) не был предком покрытосеменных.
Таким образом, предка цветковых нужно искать среди других вымерших голосеменных, более древних, точнее, обособившихся раньше, чем беннеттитовые. Фролик и Паркер предложили свою кандидатуру – користоспермовые семенные папоротники (Corystospermataceae). У них, правда, микроспорангии (пыльники) располагались на нижней, а не на верхней стороне микроспорофиллов.
Авторы предполагают, что причина всех этих противоречий, вероятно, кроется в недостаточной надежности эволюционных реконструкций, основанных на молекулярных данных. По их мнению, голосеменные растения плохо подходят для молекулярно-филогенетического анализа, поскольку большинство семейств голосеменных давно вымерло. Мы, таким образом, имеем недостаточно репрезентативную выборку таксонов, что, как хорошо известно, может приводить к ошибочным молекулярным филогениям. В конечном счете, по мнению авторов, на сегодняшний день нет оснований для безоговорочного отрицания идеи Мейена о происхождении цветковых от беннеттитовых. Эту идею вполне можно сохранить в качестве рабочей гипотезы.
По вопросу о месте первоначального возникновения покрытосеменных высказывались разные предположения. Одни считают, что они впервые появились на гипотетическом тропическом материке, расположенном между Америкой, Азией и Австралией и впоследствии погрузившемся в воды Тихого океана. Другие считают колыбелью их области современной арктической суши, третьи – горы субтропической и умеренно теплой зоны северного полушария. Большинство ботаников в настоящее время считают, что первичные покрытосеменные были древесными растениями, имевшими невысокие стволы, моноподиально разветвлявшиеся на немногочисленные толстые ветви.
Из них уже развились более крупные симподиально ветвящиеся деревья с многочисленными толстыми и тонкими ветвями. Из древесных же форм в разное время и разных филогенетических линиях развивались кустарники, полукустарники и травянистые формы, вначале многолетние, затем в различных родах в связи со специфическими условиями климата и местообитаний – двухлетники и однолетники.
Благодаря большой пластичности покрытосеменных, у них в процессе эволюции выработалось огромное разнообразие вегетативных органов, особенно в листьях, многочисленные метаморфозы, а также бесконечное разнообразие в цветках и плодах. Сложность и разнообразие химического состава и физиологических реакций также очень характерно для них.
Эволюция цветка, на строении которого главным образом базируется систематика покрытосеменных, говоря в общем и схематическом виде шла у них от цветков с длинным цветоложем (типа шишки) от обоеполых, актиноморфных со спиральным расположением свободных (несросшихся) и не фиксированных в числе членов, с верхней завязью и многочисленными семяпочками – к цветкам циклическим, зигоморфным, раздельнополым, со строго фиксированным числом более или менее сросшихся членов на плоском цветоложе, с нижней одногнездною завязью и немногими или одной семяпочкой. Эта эволюция цветка покрытосеменных происходила в разных эволюционных рядах их независимо друг от друга.
Покрытосеменные – самый крупный тип растений, к которому относится более половины всех известных видов, характеризуются рядом четких, резко отграничивающих их признаков. Наиболее характерно для них наличие пестика, образованного одним или несколькими плодолистиками (макро- и мегаспорофиллами), сросшимися своими краями, так что в нижней части пестика образуется замкнутое полое вместилище – завязь, в которой развиваются семяпочки (макро- и мегаспорангии). После оплодотворения завязь разрастается в плод, внутри которого находятся развившиеся из семяпочек семена (или одно семя). Кроме того для покрытосеменных характерны: восьмиядерный, или производный из него, зародышевый мешок, двойное оплодотворение, триплоидный эндосперм, образующийся только после оплодотворения, рыльце у пестика, улавливающее пыльцу, и для подавляющего большинства – более или менее типичный цветок с околоцветником. Из анатомических признаков для покрытосеменных характерно наличие настоящих сосудов (трахей), тогда как у голосеменных развиты только трахеиды, а сосуды встречаются крайне редко. Ввиду большого количества общих признаков можно предположить монофилетическое происхождение покрытосеменных от какой-то более примитивной группы голосеменных.
Наиболее ранние и очень отрывочные ископаемые остатки покрытосеменных (пыльца, древесина) известны из юрского геологического периода. Из нижнемеловых отложений известны тоже немногочисленные достоверные остатки покрытосеменных, а в отложениях середины мелового периода они встречаются сразу в больших количествах и в значительном разнообразии форм, которые все принадлежат ко многим различным ныне живущим семействам и даже родам.
52. Гипотезы о месте, времени, условиях происхождения покрытосеменных.
В качестве предполагаемых предков покрытосеменных указывали различные группы нижестоящих в системе растений – кейтониевые, семенные папоротники, беннеттиты, гнетовые.
Теория происхождения покрытосеменных из гнетовых предполагает, что наиболее примитивныеы покрытосеменные имели мелкие однополые цветки без околоцветника или с невзрачным околоцветником. Но по ряду соображений в настоящее время более примитивными цветками считают крупные, обоеполые цветки.
Поэтому можно предположить, что предками современных покрытосеменных были какие-то вымершие, очень примитивные голосеменные с обоеполыми цветками типа шишки (стробилами), в которых на длинном цветоложе (оси) были спирально расположены свободные (несросшиеся друг с другом) листочки однородного околоцветника, микроспорофиллы (тычинки) и мегоспорофиллы (плодолистики). В системе голосеменных эта группа должна была стоять где-то между семенными папоротниками и уже более специализированными беннеттитами и саговниками.
Покрытосемянность несомненно представляла большое преимущество в смысле защиты семяпочек и развивающихся семян от всяких неблагоприятных внешних воздействий и в первую очередь от сухости воздуха. Но одной покрытосемянностью все же трудно объяснить быстрое мощное развитие покрытосеменных и вытеснение ими господствовавших ранее на земле архегониальных растений.
Литература о происхождении покрытосеменных необозрима, но в своем подавляющем большинстве касается современных таксонов, из сравнения которых делаются попытки вывести наиболее примитивные семейства, по ним составить представление об облике предка, а далее реконструировать возможные филогенетические линии, связывающие этого предка с современными таксонами. Филогенетических схем очень много. Одни схемы монофилетические, помещающие в истоки раналиевый комплекс, другие — полифилетические. В этом случае раналиевому комплексу отводится роль лишь одного из корней покрытосеменных. Филогенетические схемы, основанные преимущественно на анализе современных форм, ниже не рассматриваются. Имеющихся же палеоботанических материалов недостаточно, чтобы предложить сколько-нибудь конкретную филогению таксонов и указать группу голосеменных в ранге порядка, давшую начало покрытосеменным. Поэтому ограничимся знакомством с древнейшими известными покрытосеменными и лишь в общих чертах обсудим вопрос о том, какие голосеменные в принципе могли дать начало покрытосеменным.
Больше всего дискуссий связано с происхождением плодолистика и гинецея в целом. Популярна гипотеза кондупликатного происхождения плодолистика, выводимого из листа, несущего семезачатки (филлосперма или кладосперма). Согласно этой гипотезе, в онтогенезе предка такой лист был сначала заключен в почке в сложенном виде, семезачатками вовнутрь. Эта ранняя онтогенетическая стадия предка и удержалась у взрослых форм первых покрытосеменных. Семезачатки созревали, а лист оставался сложенным вдоль средней жилки. Край листа постепенно преобразовывался в рыльцевую поверхность для приема пыльцы. В качестве возможных предков при этом указывались птеридоспермы с семенами, сидящими на вайях. Приводился пермский род Archaeocycas (его считали возможным предком и цикадовых). Можно представить, как листовая пластинка Archaeocycas разрастается, а семезачатки оказываются спрятанными в подобие листовки.
При установлении предка покрытосеменных следует учитывать следующие обстоятельства. Во-первых, некоторые признаки голосеменных оказываются чрезвычайно выдержанными в течение долгой эволюции основных таксонов. Например, среди Pinopsida за всю их историю не появилось листьев с сетчатым жилкованием. В классе Ginkgoopsida семена прикрепляются к уплощенным семеносным органам только абаксиально. В классе Cycadopsida отмечается повторное образование сетчатого жилкования и почти не отмечается тенденция к расслоению экзины с образованием мешков. С другой стороны, в разных классах голосеменных проявляются некоторые сходные преобразования, приводящие к независимому появлению очень сходных органов. Таково, например, поразительное сходство пыльцы (включая внутриклеточный гаметофит) Callistophytales и хвойных, листьев семейства Cardiolepidaceae и разных хвойных. Если изложенная выше филогения голосеменных в основных чертах правильна, то многочисленные черты сходства гинкговых с хвойными возникли параллельно.
Следовательно надо не просто искать среди голосеменных формы, сходные с покрытосеменными по выборочным, пусть даже очень заметным признакам, которые всегда могут оказаться возникшими независимо. Надо попытаться найти у покрытосеменных признаки, особенно хорошо выдерживающиеся у голосеменных в их эволюции и дающие возможность разделения голосеменных на главные группы. Если такие признаки обнаружатся, можно установить, от каких голосеменных они унаследованы.
Как уже отмечалось, в ходе эволюции разных классов голосеменных из всех их признаков наиболее устойчивы типы семян — платиспермический, радиоспермический и вторично-платиспермический. Платиспермические семена всегда бескупульные, а если сопровождаются купулоподобным кладоспермом (вроде воротничка гинкго), то сохраняют главные признаки платиспермии. Вторично-платиспермические семена Pinopsida от карбоновых кордаитантовых до современных хвойных неизменно имеют интегумент, возникающий из двух примордиев и васкуляризованный двумя пучками у менее продвинутых форм (дополнительные пучки, если они есть, отходят от главных и расположены в одной с ними плоскости; у более продвинутых форм васкуляризация исчезает). У Cycadopsida сохраняется радиоспермия и купула, а у более продвинутых форм — образовавшийся из купулы внешний интегумент.
Учитывая поразительную устойчивость этих признаков семян в филогенетических линиях, можно ожидать, что они удержались и при переходе от голосеменных к покрытосеменным. Можно исключить из числа возможных предков всех Pinopsida, филогенетическая независимость которых от покрытосеменных видна по широкому комплексу признаков. Если допустить, что все остальные голосеменные принадлежат Ginkgoopsida или Cycadopsida (а не какому-нибудь еще неоткрытому классу), то следует сделать выбор между этими двумя классами, проанализировав тип семян покрытосеменных. У них, в отличие от голосеменных, количество интегументов сильно варьирует, иногда даже в пределах рода. Если интегумент один, то нельзя точно указать, какому органу голосеменных он соответствует, а именно: единственному интегументу Ginkgoopsida, внутреннему интегументу (васкуляризованному нуцеллюсу) Cycadopsida или купуле, слившейся с интегументом. В битегмических (с двумя интегументами) семезачатках васкуляризация внешнего интегумента очень разнообразна или отсутствует. Васкуляризация внутреннего интегумента отмечается редко, но если во внутренний интегумент проходит более одного пучка, то они располагаются по кругу, т. е. как у радиоспермических форм. Можно предположить, что покрытосеменным в целом свойственна радиоспермия. Соответственно их предков надо искать среди радиоспермических голосеменных. Тогда всех представителей класса Ginkgoopsida, в том числе каллистофитовых, арбериевых, пельтаспермовых, кейтониевых, пентоксилеевых и лептостробовых, уже нельзя рассматривать как возможных предков. С этим согласуется и уже упоминавшееся абаксиальное прикрепление семян у Ginkgoopsida. Покрытосеменные с листоиодобными плодолистиками характеризуются адаксиальной (на верхней поверхности) или краевой, но не абаксиальной плацентацией.
Приняв происхождение покрытосеменных от радиоспермических голосеменных, обратимся к мезозойским таксонам последних (лагеностомовые и тригонокарповые, видимо, вымерли до конца палеозоя), из которых известны лишь цикадовые и беннеттитовые. Полиспермы цикадовых могли преобразоваться в плодолистики, тем более, что у цикадовых встречаются краевая и адаксиальная планцентации. Труднее установить преемственность цикадовых и покрытосеменных по строению мужских фруктификаций, типу прохождения листового следа и некоторым другим признакам. Беннеттиты считались предками покрытосеменных, но не удавалось вывести плодолистик из головчатых полиспермов. Однако это затруднение можно преодолеть, если принять во внимание возможность гамогетеротопии (переноса признаков с одного пола на другой), известной у многих животных и предполагаемой у кордаитантовых; плодолистик мог возникнуть путем переноса структуры мужских фруктификаций на женские. Тогда предками покрытосеменных могли быть голосеменные с такими мужскими фруктификациями, которые по общей структуре близки к плодолистикам, но место семезачатков занимают синангии, сорусы или спорангии. Именно таково строение микроспорофиллов некоторых беннеттитовых. Мысленно заменив синангии битегмическими семезачатками, можно представить орган, который достаточно сложить вдоль, чтобы получить примитивную листовку. Тогда становятся понятными многочисленные черты сходства покрытосеменных и беннеттитов (парацитные устьица, строение древесины, неслоистая нэкзина, присутствие нектарников и др.). Находит объяснение и давно отмечаемое сходство покрытосеменных и гнетовых. Происхождение гнетовых от беннеттитовых считается весьма вероятным. Следовательно, сходство гнетовых и покрытосеменных оказывается связанным с общим происхождением обеих групп от беннеттитовых. Изложенная гипотеза согласуется с общей фитогеографической ситуацией в начале мела. Считается, что покрытосеменные появились в семиаридных районах экваториального пояса, т. е. там, где беннеттиты были обычным компонентом растительности.

Цветко́вые расте́ния, покры́тосеменны́е (лат. Magnoliophyta , или Angiospermae) — отдел высших растений, имеющих цветок. Эта важнейшая группа наземных растений насчитывает свыше 165 порядков, 540 семейств, более 13 000 родов и, вероятно, не менее 250 000 видов. По числу видов цветковые растения значительно превосходят все остальные группы высших растений, вместе взятые. Они составляют одну из двух групп семенных растений.
Содержание
Морфологические особенности
Важнейшая особенность цветковых растений — наличие специализированного генеративного органа — цветка, берущего на себя функции полового размножения и привлечения агентов опыления. Цветковые растения заключают свои семязачатки (семяпочки) в полость завязи, которая образована срастанием когда-то открытого плодолистика. Стенки завязи после оплодотворения разрастаются и видоизменяются, давая образование под названием плод.
В другой группе семенных растений, Голосеменные (Pinophyta, или Gymnospermae), семязачаток не скрыт от опыления, а семена не заключены в истинный плод, но иногда семя могут покрывать мясистые структуры, например, у представителей рода Тис.
Размножение
Происхождение
Первые остатки покрытосеменных датируются Юрским периодом приблизительно 140 миллионов лет назад. Базируясь на современных данных, можно предположить, что предки покрытосеменных и гнетовых дивергировали в триасе (220—202 миллионов лет назад). Первые отпечатки растений с признаками покрытосеменных обнаружены в пластах юрского и раннего мелового периодов (135-65 миллионов лет назад), но это были довольно-таки малочисленные и примитивные формы. Древнейшими покрытосеменными являются растения из группы нимфейных [1] . Следы широкого развития и распространения покрытосеменных появились в палеонтологической летописи в период среднего мела (около 100 миллионов лет назад). Но уже в позднем мелу покрытосеменные оказались доминирующей формой растительной жизни, и в многих фоссилиях узнаются представители современных семейств (например, бук, дуб, клен и магнолия)
Одно из важнейших направлений эволюции растительного царства — приспособление к изменчивым условиям наземной жизни. Цветковые растения являются ярчайшим выражением этой линии и доминируют на земной поверхности в данную эпоху. От полюсов до экватора нет такого участка, где возможна растительная жизнь, но не найдено покрытосеменных.
К широчайшему географическому разнообразию прибавляется разнообразие форм и способов роста. Банальная ряска, покрывающая поверхность пруда, представляет собой крошечный зелёный побег с простым корешком, вертикально погруженным в воду, и с очень нечёткими листиками и частями стебля. Могучее лесное дерево столетия развивало свою сложную систему стволов и ветвей, покрытых бесчисленными веточками и листвой, а под землёй соответствующую площадь занимает мощная, хорошо развитая корневая система. Между этими двумя крайностями — бесконечные градации: водные и земные травы, ползучие, прямостоящие или карабкающиеся, кусты и деревья, гораздо большее разнообразие, чем в другом отделе семенных растений — Gymnospermae.
Известны многочисленные водные покрытосеменные растения, они в изобилии встречаются в долинах рек и чистых озёрах, в меньшем количестве — в солёных озёрах и морях. Однако такие водные покрытосеменные не являются примитивными формами, а возникли путём приспособления наземного предка к водной среде.
Систематическое положение
Цветковые растения обычно рассматриваются как отдел. Так как эта систематическая категория более высокого ранга, чем семейство, есть определённая свобода в выборе названия. Статья 16 Международного Кодекса Ботанической Номенклатуры позволяет использовать как и традиционные исторические названия, так и название, образованное от рода. Официальное униноминальное название этого таксона — Magnoliophyta, от названия рода Magnolia. Но традиционно укоренились такие имена, как Angiospermae и Anthophyta (цветковые растения).
Классификация
История понятия
Благодаря постоянному пересмотру взглядов на родство цветковых растений, внутренняя систематика этой группы подвергалась и подвергается изменениям. Две широко используемые, хотя и несколько устаревшие, системы цветковых растений — система Тахтаджяна и система Кронквиста, не отражают филогению таксона. Таким образом, классификация цветковых растений сейчас активно дорабатывается и исправляется.
Отдел цветковых растений традиционно подразделяется на 2 класса — Magnoliopsida (двудольные) от названия рода Magnolia и Liliopsida (однодольные) от названия рода Lilium. Более популярны традиционные названия этих таксонов — Dicotyledones и Monocotyledones. Происхождение этих названий вполне очевидно: Dicotyledones имеют две семядоли в семени, тогда как у Monocotyledones семядоля одна.
Классы двудольных и однодольных, в свою очередь, подразделяются на подклассы, которые делятся на порядки (иногда объединяемые в надпорядки), семейства, роды и виды со всеми промежуточными категориями. Имеется целый ряд современных систем классификации цветковых растений.
Класс Двудольные
В классе Двудольные (Magnoliopsida, или Dicotyledones) описано 8 подклассов, 128 порядков, 418 семейств, приблизительно 10 000 родов и около 190 000 видов двудольных растений. Выделяют подклассы:
- Подкласс 1. Магнолиевые (Magnoliidae)
- Подкласс 2. Гамамелисовые (Hamamelididae)
- Подкласс 3. Гвоздичные (Caryophyllidae)
- Подкласс 4. Диллениевые (Dilleniidae)
- Подкласс 5. Розоцветные (Rosidae)
- Подкласс 6. Сложноцветные (Asteridae)
Класс Однодольные
В класс однодольных растений (Liliopsida, или Monocotyledones) включаются 5 подклассов, 37 порядков, около 120 семейств, 30 000 родов и больше 60 000 видов. Выделяют подклассы:
Отдел цветковых, или покрытосеменных, растений — это самая многочисленная и хорошо приспособленная к современным условиям группа представителей флоры. Они довольно разнообразны и включают в себя травы, кустарники и деревья. Их основной отличительной чертой является наличие цветка как органа размноженья, а также замкнутого вместилища у семян, откуда и происходит название отдела.

Происхождение и эволюция
Считается, что предком этой группы растений были водоросли, не сохранившиеся до современности. Затем они эволюционировали в папоротники, которые в свою очередь дали начало как покрытосеменным, так и голосеменным видам Они произошли от общих предков, разделившихся на разные ветви в триасе.

Всего цветковые существуют около 140 миллионов лет — первые представители фауны с похожим строением появились в юрском периоде. Это были довольно примитивные и малочисленные виды. Древнейшими представителями цветковых считаются представители семейства нимфейные или кувшинковые. В середине мелового периода (около 100 миллионов лет назад) покрытосеменные уже были довольно широко распространены в мире. В конце мела они оказались доминирующей формой жизни и покрывали значительную часть суши. Примерно тогда же начали появляться предки многих современных растений — дуба, клёна, магнолии, бука. Одной из главной особенностей эволюции цветковых стала их высокая приспосабливаемость к изменяющимся условиям, что позволило им произрастать на разных географических широтах. Они отличаются огромным многообразием форм, размеров, способов развития и размножения. Среди цветковых можно найти водные и наземные, ползучие, вьющиеся, карабкающиеся и прямостоящие растения, которые растут и процветают во всех уголках Земли, от тропических лесов и долин рек до солёных морей и альпийских лугов. Другой интересной особенность эволюции покрытосеменных является большое разнообразие форм водяных растений. Они не произошли от водорослей напрямую, а эволюционировали от наземного предка в результате смены условий, потому отличаются сложным строением и большим количеством приспособительных механизмов.
Основные признаки
Для того чтобы дать краткую характеристику покрытосеменным растениям, нужно посмотреть на их основные признаки. Их список выглядит так:

- растение состоит из стебля, корневой системы, листьев и цветка;
- семя защищено околоплодником — оболочкой, которая обеспечивает его сохранность при распространении;
- семязачатки и семена хранятся в завязи и плоде;
- женский заросток представляет собой зародышевый мешок с восемью ядрами, а мужской — пыльцевое зерно, состоящее из вегетативных и генеративных клеток;
- двойное оплодотворение — спермии (мужские гаметы) цветка воздействуют одновременно на яйцеклетку и ядро зародышевого мешка;
- для размножения растению необходимо опыление, которое может осуществляться по воздуху или воде, а также с помощью переноса пыльцы птицами и насекомыми;
- завязь оплодотворённого цветка преобразуется в плод, семяпочка — в семя, зигота — в зародыш семени, вторичное ядро — в эндосперм;
- половое размножение с постоянной сменой поколений.
К другим типичным признакам покрытосеменных относятся ситовидные трубки на стебле, широкие пластинчатые листья со сложной системой сетчатых жилок, цветки циклической формы. Но стоит отметить, что ни один из характерных признаков группы не свойственен всем без исключения её членам. Принадлежность вида к цветковым определяется по общему числу признаков, но некоторые из них могут отсутствовать.
Классы покрытосеменных
Всех представителей отдела покрытосеменных делят на однодольные и двудольные растения. Как можно понять из их названия, они отличаются друг от друга по количеству семядолей. Сравнительная характеристика этих классов:
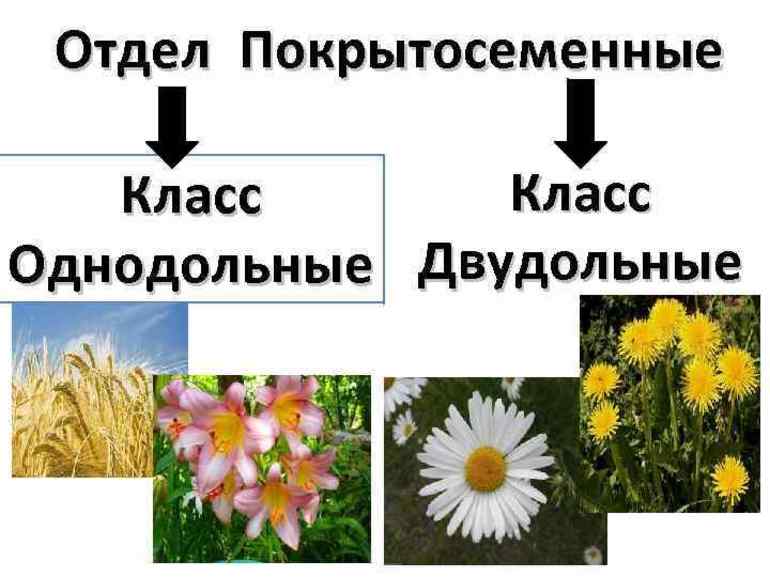
- для однодольных характерна корневая система мочковатой формы, состоящая в основном из придаточных отростков, а для двудольных — мощный главный корень в форме стержня;
- листья растений первого класса имеют простую структуру и состоят из обычной цельной пластинки, в то время как у второго они могут иметь более сложное строение;
- жилкование у однодольных параллельное или дуговое, а у двудольных — сетчатое;
- для растений с одной семядолей характерно количества частей цветка, кратное трём, а с двумя — четырём или пяти;
- у однодольных покрытосеменных нет камбия — образовательной ткани, обеспечивающей рост стебля и корней в толщину;
- проводящие пучки у двудольных расположены в виде кольца, а у родственного класса — беспорядочно и хаотично.
В класс двудольных входят высокоразвитые деревья и кустарники, в том числе многочисленные плодовые и декоративные культуры. Их общее количество достигает 180—190 тысяч.
Семейства однодольных
Всего в современной классификации насчитывается более 500 семейств покрытосеменных, из которых 12 — двудольные. Они отличаются строением стебля и листьев, формулой цветка, длительностью жизненного цикла, местами, где они предпочитают расти и многими другими особенностями. Глядя на фото и названия цветковых растений разных видов, трудно не поразиться их разнообразию — в один отдел входят самые разные формы жизни, от водяной ряски до огромного дуба.

- стебель-соломина;
- небольшие корни с развитыми придатками;
- сворачивающиеся при жаре листки;
- простые цилиндроподобные цветки, собранные в злаки;
- сухие невскрывающиеся плоды;
- опыление ветром.

- подземные луковицы или клубнелуковицы, которые формируются из побегов и могут защитить почки от неблагоприятного воздействия внешней среды во время жары заморозков;
- прямой или вьющийся надземный стебель, который называется цветочной стрелкой;
- узкие мясистые листья;
- широкое разнообразие соцветий;
- цветки обоеполые;
- плод-коробочка или ягода;
- опыление насекомыми, в тропических широтах также птицами, редко — ветром.
К примерам цветковых растений из семейства лилейных можно отнести самые разные виды, от ландышей до лука. Бо́льшую часть из них составляют многолетние травы, реже встречаются деревья и кустарники.
Двудольные растения
Среди группы двудольных растений выделяется класс розоцветных. Эти растения насчитывают 300 видов в 100 родах. Распространены в субтропиках и местах с умеренным климатом. Отличаются особым строением цветоложа. Опыляются насекомыми, привлекая их нектаром или выделяя очень большое количество пыльцы, чтобы размножаться. Все представители класса имеют очень похожие цветки. Но их плоды отличаются широким разнообразием — среди них можно найти ягоды, орешки, листовки, яблоки, коробочки. Многие из них приспособлены к распространению животными.

Семейство бобовых получило своё название за плоды в виде бобов или стручков. Оно состоит из 24,5 тысяч растений, объединённых в 946 родов. К особенностям, которые могут указать на принадлежность к этому классу, относятся стоячие вьющиеся стебли, сложные по строению листья с прилистником и цветы из пяти лепестков. Цветы обоеполые. Опыляются насекомыми. К семейству паслёновых относятся травы, кусты и маленькие деревья. К особенностям, которые могут показать принадлежность вида к ним, относится расположение листьев. В вегетативной части они растут попеременно, а в цветоносной — попарно. Из-за этого стебли у представителей флоры кажутся разветвлёнными на две или три части. Другие особенности семейства — соцветия в форме завитков, небольшое количество тычинок равной длины, похожие на почки семена, плоды в виде ягод или коробочек.

Крестоцветные растения разделены на 4 тысячи видов, которые включают 400 разновидностей. В семейство входит много двулетних монокарпических растений с особой схемой цветения — они размножаются только один раз на последнем году жизни. К особенностям также относится стержневая корневая система (реже встречаются корнеплоды), простые листья без прилистника, толстый стебель, 6 тычинок, плод-стручок и кистеобразные соцветия. Ещё одним интересным семейством двудольных растений являются астровые, или сложноцветные. Это одна из самых больших групп, которая насчитывает почти 40 тысяч видов в 1911 родах. Их можно найти в любом уголке земного шара, от тундры до тропиков. Основной особенностью семейства является соцветие-корзинка, которое образуется из множества очень мелких цветков. Кроме того, для них характерен развитый корень-стержень и плод-семянка, не содержащий белка.
Хозяйственное значение
Трудно переоценить то, сколько пользы человеку приносят покрытосеменные растения. Их используют в следующих сферах жизни:

- пищевая промышленность, изготовление кормов для животных;
- медицина — как лекарственные растения в составе различных препаратов;
- дизайн интерьера и ландшафта — как декорации;
- ремесло — для изготовления духов, косметических средств, красителей, масел и прочего;
- в культуре как подарки и сувениры;
- в науке для исследований генетики.
Экологическое положение
Сегодня покрытосемянные являются самыми многочисленным растениями, господствующими по всему земному шару. Чтобы понять, почему они смогли занять эту нишу, достаточно посмотреть на их эволюционный путь — эти представители флоры развивались в мезозойскую и кайнозойскую эру, когда условия были очень близки к современным.

Эти растения получили центральную нишу в экосистемах и в какой-то мере повлияли на эволюцию почти всех животных и птиц, для которых они были источником пищи и основой среды обитания. Между тем большую роль в этом деле сыграла и деятельность человека — в частности, окультуривание растений и выведение новых сортов, приспособленных под разные условия.
Несмотря на это, отдельные виды покрытосеменных находятся под угрозой исчезновения и занесены в Красную книгу. Причиной этого является в основном деятельность людей — вырубка лесов, пожары, осушение и затопление зон для ведения сельского хозяйства, накопление газа в атмосфере.
Покрытосеменные растения — наиболее распространённая и разнообразная группа представителей флоры. Они играют очень важную роль как в жизни человека, так и в развитии экосистем. Разные классы, подклассы, семейства и виды цветковых могут очень сильно отличаться друг от друга и не иметь почти никаких общих внешних черт и особенностей жизнедеятельности, кроме защищённого околоплодником семени.
Читайте также:
- Как создать незатухающие колебания в контуре кратко
- Почему испарение происходит тем быстрее чем выше температура жидкости кратко
- Как отразились на внешнеполитическом курсе ссср внутренние проблемы развития страны кратко
- Профессиональная деятельность специалистов доу
- Что имеют общего и чем отличаются наречия от других частей речи кратко