
Как происходит регуляция процессов сокращения и расслабления мышечных волокон кратко
Обновлено: 01.07.2024
Управление мышцей, состоящей из значительного числа двигательных единиц (ДЕ), осуществляется совокупностью мотонейронов, иннервирующих мышцу, получившей название мотонеронный пул (МП). Известно, что мотонейроны, посылающие свои аксоны к той или иной мышце, могут располагаться не только в пределах одного сегмента спинного мозга, но и занимать соседние. Таким образом, структурно МП может быть разнесен на достаточно большое расстояние в пределах нескольких сегментов передних рогов спинного мозга. Функционально МП является конечной инстанцией, где формируется структура командных сигналов на мышцу, обеспечивающих ее включение в двигательный акт. Именно на МП осуществляется интеграция входных воздействий на мотонейроны от надсегментарных структур и от рецепторов двигательной периферии.
Основная функция МП - дозирование силы сокращения мышцы - обеспечивается двояко - частотой импульсации входящих в него мотонейорнов и количеством активированных мотонейронов данного пула.
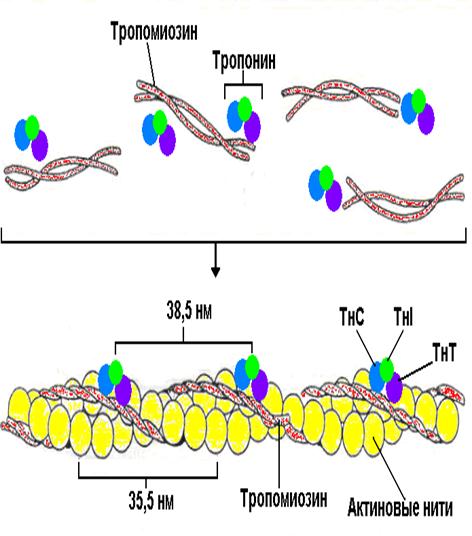
Рис. 34. Структура тонкого филамента.
Через 50 мс после произвольной внутренней команды начинается сокращение мышцы скелетной. За это время команда передается от коры к мотонейронам спинного мозга и по двигательным волокнам к мышце. Медиатором в нервно-мышечном синапсе является ацетилхолин, который содержится в синаптических пузырьках пресинапса. Нервный импульс вызывает опорожнение синаптических пузырьков и выход ацетилхолина в синаптическую щель. Там медиатор действует на постсинаптический рецептор, после чего разрушается. По мере расходования запасы ацетилхолина пополняются путем синтеза в пресинаптической мембране, но если импульсы идут часто и долго, то расход превышает пополнение и нарушается проведение возбуждения через нервно-мышечный синапс. В результате наступает утомление.
Сокращение мышцы является результатом сокращения составляющих ее мышечных клеток (волокон). Сокращение мышечного волокна – это результат укорочения каждого его саркомера за счет взаимодействия тонких и толстых филаментов.
В покоящейся мышце тонкие филаменты контактируют с Z –линиями и не достигают центра саркомера, а толстые находятся в центре, но не достигают Z-линии. Только по бокам А-диска в пространство между толстыми филаментами немного входят тонкие (Рис. 33, В).
При умеренном усилии происходит умеренное сокращение за счет того, что тонкие филаменты движутся навстречу друг другу, поэтому расстояние между Z – линиями уменьшается и длина мышцы уменьшается.
При максимальном сокращении толстые филаменты касаются Z –линий.
Физиолого-биохимический механизм сокращения состоит в том, что после взаимодействия медиатора с рецептором при достаточной частоте нервных импульсов на мышечной мембране развивается мышечный потенциал действия, который быстро распространяется вдоль мышечного волокна, вызывая выход Са 2+ из саркоплазматического ретикулума. Затем Са 2+ проникает в миофибриллы к центрам связывания на молекуле тропонина. В покоящейся мышце тропомиозин препятствует присоединению миозиновой головки к ближайшему с ним мономеру актина. Связывание Са с тропонином изменяет его пространственную структуру, что ослабевает связь между его тропомиозинсвязывающей субъединицей и актином. В результате молекула тропомиозина начинает двигаться по желобку тонкого филамента, освобождая скрытый до этого миозинсвязывающий центр на поверхности молекулы актина. Начинается взаимодействие актина с миозином, что сближает молекулы, принадлежащие тонким и толстым филаментам. В результате расстояние между Z-линиями уменьшается. При связывании актина и миозина молекула АТФ распадается на АДФ и фосфор неорганический. Распад АТФ ведет к расслаблению мышцы из-за изменения конформации миозина. Для восстановления способности к сокращению к головке миозина должна присоединиться следующая молекула АТФ. Весь процесс от появления мышечного потенциала до сокращения мышечного волокна называется электромеханической связью (сопряжением). Сокращение гладких мышц имеет особенность: Са 2+ связывается с особым белком – кальмодулином.
Управление мышцей, состоящей из значительного числа двигательных единиц (ДЕ), осуществляется совокупностью мотонейронов, иннервирующих мышцу, получившей название мотонеронный пул (МП). Известно, что мотонейроны, посылающие свои аксоны к той или иной мышце, могут располагаться не только в пределах одного сегмента спинного мозга, но и занимать соседние. Таким образом, структурно МП может быть разнесен на достаточно большое расстояние в пределах нескольких сегментов передних рогов спинного мозга. Функционально МП является конечной инстанцией, где формируется структура командных сигналов на мышцу, обеспечивающих ее включение в двигательный акт. Именно на МП осуществляется интеграция входных воздействий на мотонейроны от надсегментарных структур и от рецепторов двигательной периферии.

Основная функция МП - дозирование силы сокращения мышцы - обеспечивается двояко - частотой импульсации входящих в него мотонейорнов и количеством активированных мотонейронов данного пула.
Рис. 34. Структура тонкого филамента.
Через 50 мс после произвольной внутренней команды начинается сокращение мышцы скелетной. За это время команда передается от коры к мотонейронам спинного мозга и по двигательным волокнам к мышце. Медиатором в нервно-мышечном синапсе является ацетилхолин, который содержится в синаптических пузырьках пресинапса. Нервный импульс вызывает опорожнение синаптических пузырьков и выход ацетилхолина в синаптическую щель. Там медиатор действует на постсинаптический рецептор, после чего разрушается. По мере расходования запасы ацетилхолина пополняются путем синтеза в пресинаптической мембране, но если импульсы идут часто и долго, то расход превышает пополнение и нарушается проведение возбуждения через нервно-мышечный синапс. В результате наступает утомление.
Сокращение мышцы является результатом сокращения составляющих ее мышечных клеток (волокон). Сокращение мышечного волокна – это результат укорочения каждого его саркомера за счет взаимодействия тонких и толстых филаментов.
В покоящейся мышце тонкие филаменты контактируют с Z –линиями и не достигают центра саркомера, а толстые находятся в центре, но не достигают Z-линии. Только по бокам А-диска в пространство между толстыми филаментами немного входят тонкие (Рис. 33, В).
При умеренном усилии происходит умеренное сокращение за счет того, что тонкие филаменты движутся навстречу друг другу, поэтому расстояние между Z – линиями уменьшается и длина мышцы уменьшается.
При максимальном сокращении толстые филаменты касаются Z –линий.
Физиолого-биохимический механизм сокращения состоит в том, что после взаимодействия медиатора с рецептором при достаточной частоте нервных импульсов на мышечной мембране развивается мышечный потенциал действия, который быстро распространяется вдоль мышечного волокна, вызывая выход Са 2+ из саркоплазматического ретикулума. Затем Са 2+ проникает в миофибриллы к центрам связывания на молекуле тропонина. В покоящейся мышце тропомиозин препятствует присоединению миозиновой головки к ближайшему с ним мономеру актина. Связывание Са с тропонином изменяет его пространственную структуру, что ослабевает связь между его тропомиозинсвязывающей субъединицей и актином. В результате молекула тропомиозина начинает двигаться по желобку тонкого филамента, освобождая скрытый до этого миозинсвязывающий центр на поверхности молекулы актина. Начинается взаимодействие актина с миозином, что сближает молекулы, принадлежащие тонким и толстым филаментам. В результате расстояние между Z-линиями уменьшается. При связывании актина и миозина молекула АТФ распадается на АДФ и фосфор неорганический. Распад АТФ ведет к расслаблению мышцы из-за изменения конформации миозина. Для восстановления способности к сокращению к головке миозина должна присоединиться следующая молекула АТФ. Весь процесс от появления мышечного потенциала до сокращения мышечного волокна называется электромеханической связью (сопряжением). Сокращение гладких мышц имеет особенность: Са 2+ связывается с особым белком – кальмодулином.
Рассмотрим, к чему сводятся представления о механизме попеременного сокращения и расслабления мышц. В настоящее время принято считать, что биохимический цикл мышечного сокращения состоит из 5 стадий (рис. 20.8):
3) это взаимодействие обеспечивает высвобождение АДФ и Н3РО4 из актин-миозинового комплекса. Актомиозиновая связь имеет наименьшую энергию при величине угла 45°, поэтому изменяется угол миозина с осью фибриллы с 90° на 45° (примерно) и происходит продвижение актина (на 10–15 нм) в направлении центра саркомера (см. рис. 20.8, в);
4) новая молекула АТФ связывается с комплексом миозин–F-актин (см. рис. 20.8, г);
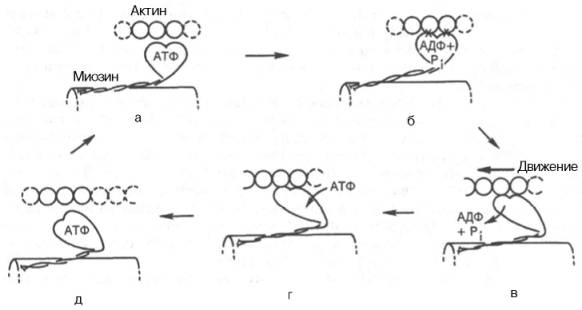
Рис. 20.8. Биохимический цикл мышечного сокращения. Объяснение в тексте.
Регуляция сокращения и расслабления мышц. Сокращение любых мышц происходит по общему механизму, описанному ранее. Мышечные волокна разных органов могут обладать различными молекулярными механизмами регуляции сокращения и расслабления, однако всегда ключевая регулятор-ная роль принадлежит ионам Са 2+ . Установлено, что миофибриллы обладают способностью взаимодействовать с АТФ и сокращаться в его присутствии лишь при наличии в среде определенных концентраций ионов кальция . Наибольшая сократительная активность наблюдается при концентрации ионов Са 2+ около 10 –6 –10 –5 М. При понижении концентрации до 10 –7 М или ниже мышечные волокна теряют способность к укорочению и развитию напряжения в присутствии АТФ.
По современным представлениям, в покоящейся мышце (в миофибрил-лах и межфибриллярном пространстве) концентрация ионов Са 2+ поддерживается ниже пороговой величины в результате связывания их структурами (трубочками и пузырьками) саркоплазматической сети и так называемой Т-системой при участии особого Са 2+ -связывающего белка, получившего название кальсеквестрина, входящего в состав этих структур.
Возможность пребывания живой мышцы в расслабленном состоянии при наличии в ней достаточно высокой концентрации АТФ объясняется снижением в результате действия кальциевой помпы концентрации ионов Са 2+ в среде, окружающей миофибриллы, ниже того предела, при котором еще возможны проявление АТФазной активности и сократимость акто-миозиновых структур волокна. Быстрое сокращение мышечного волокна при его раздражении от нерва (или электрическим током) является результатом внезапного изменения проницаемости мембран и как следствие выхода из цистерн и трубочек саркоплазматической сети и Т-системы некоторого количества ионов Са 2+ в саркоплазму.
Описан процесс сокращения скелетных мышц человека, который состоит из ряда этапов. Первый этап связан с поступлением нервного импульса по аксону мотонейрона к мышечному волокну. Второй этап заключается в возникновении потенциала действия и распространении его вдоль мышечного волокна. В результате третьего этапа в саркоплазму выделяются ионы кальция, что приводит к началу взаимодействия толстого и тонкого филаментов. Четвертый этап представляет собой скольжение тонких филаментов относительно толстых, что составляет собственно сокращение мышцы.
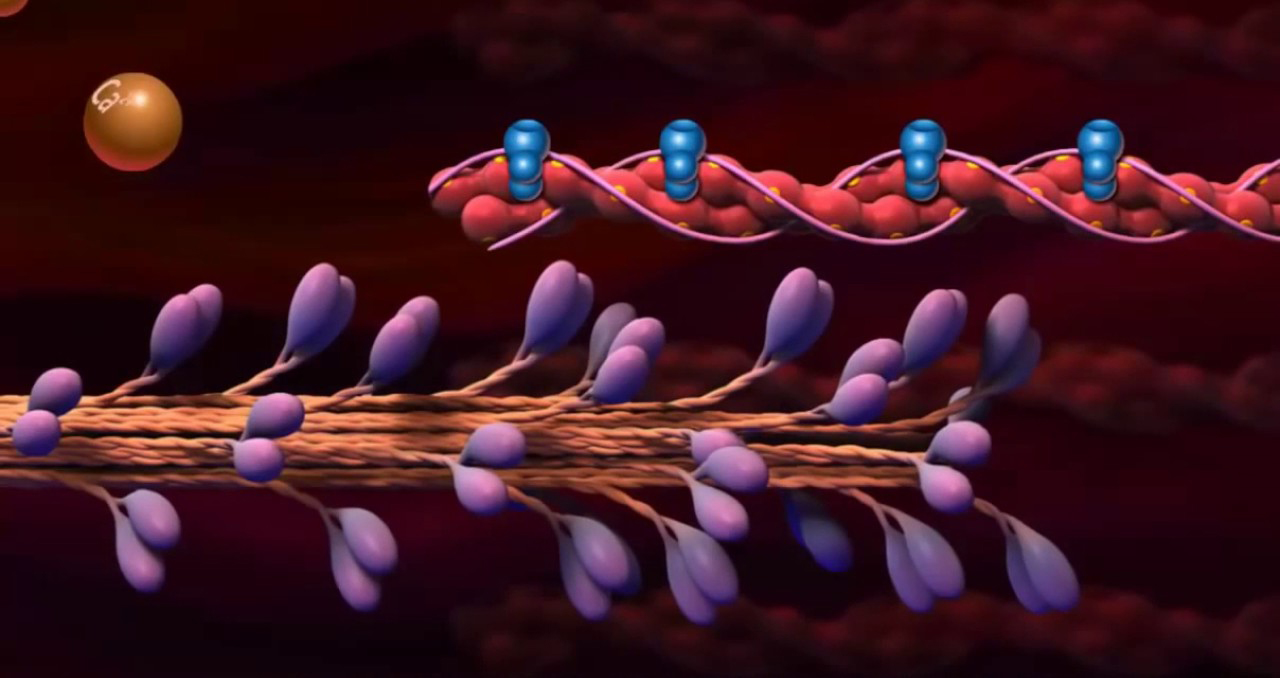
Сокращение скелетных мышц человека
Давайте теперь разберемся в механизме сокращения мышцы, точнее в механизме сокращения мышечных волокон, а еще более точно в механизме сокращения миофибрилл или другими словами, в механизме сокращения саркомера. Этот процесс можно условно разделить на несколько этапов.
Поступление нервного импульса к мышечному волокну
Чтобы сократиться, мышца должна получить сигнал из центральной нервной системы (ЦНС). Такими сигналами являются импульсы, поступающие по мотонейрону к мышце.
Вспомним, что при подходе к мышце аксон мотонейрона ветвится, то есть пускает веточки к мышечным волокнам. Если такого соединения нет, мышечное волокно сокращаться не будет и постепенно атрофируется.
Возникновение потенциала действия
После того, как по аксону мотонейрона к мышечным волокнам приходит импульс, из него в области соединения выделяется ацетилхолин. Выделение этого нейромедиатора (ацетилхолина) приводит к протеканию ряда процессов, в результате которых меняется полярность сарколеммы мышечного волокна. Это называется деполяризацией сарколеммы мышечного волокна. В результате развивается потенциал действия.
Выделение ионов кальция
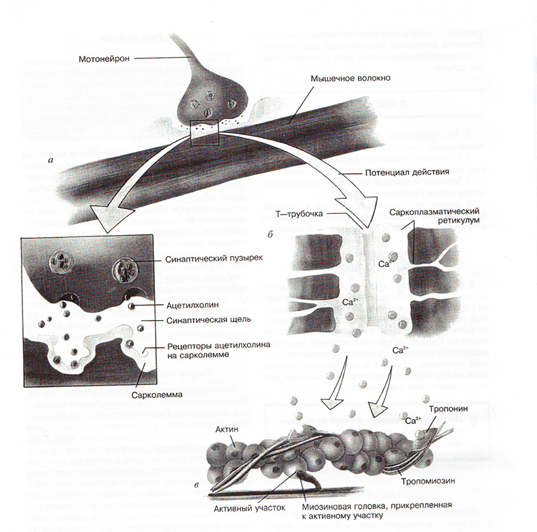
Рис.1. Механизм сокращения скелетных мышц
Затем ионы кальция соединяются с тропонином (тропонин – один из белков тонкого филамента). Этот белок имеет шарообразную форму и расположен в тонком филаменте регулярно через определенные расстояния. После соединения с ионами кальция, тропонин меняет свою конфигурацию и приподнимает длинные тропомиозиновые трубки. Когда мышца неактивна, длинные трубки белка тропомиозина закрывают активные центры на актине. После того как тропомиозиновые трубки приподнимаются, на актине открываются активные центры. К ним теперь могут прикрепляться миозиновые головки.
Сокращение саркомера (гребковая гипотеза, теория скользящих нитей)
Прекращение импульсов, поступающих от мотонейрона к мышечному волокну приводит к расслаблению мышцы.
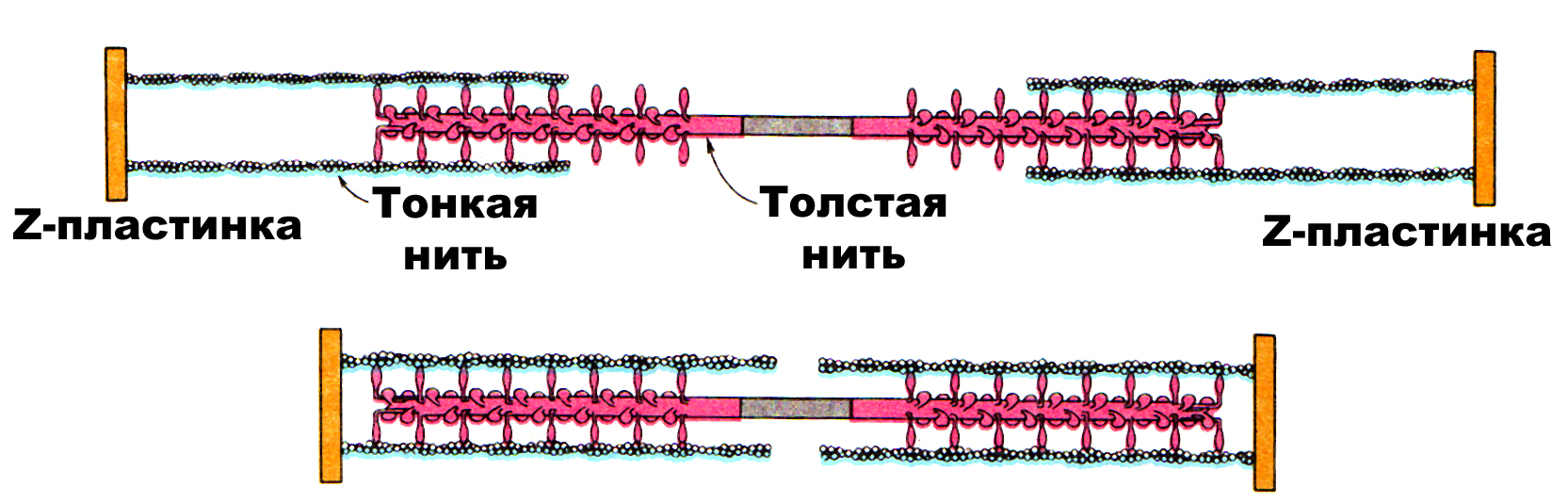
Рис.2. Схема, иллюстрирующая взаимодействие толстого и тонкого филаментов (Л. Страйер, 1985)
Похожие записи:
Effect of KAATSU-training on the maximum voluntary isometric contraction of lower extremity muscles of qualified football players
Изучалось влияние KAATSU-тренинга на изометрическую силу мышц квалифицированных футболистов. Установлено, что интенсивный рост максимальной силы мышц…
Удаление фасции снижает силу мышц
В опытах на диких индейках показано, что удаление фасции снижает силу мышц на 30%. Эти результаты ставят под…
Искусственный интеллект в спортивной тренировке
Описана система комплексного контроля в спорте, построенная на основе искусственного интеллекта. Разработанная система внедрена в подготовку пауэрлифтеров высокой…
Тест времени реакции на сигнал
Представлена программа расчета времени реакции на сигнал, предназначенная для использования в учебных целях, например на занятиях по…
Саркоплазматическая гипертрофия мышц
Дано определение и описаны механизмы саркоплазматической гипертрофии скелетных мышц. Показано, что этот вид гипертрофии мышц широко…
Классификация типов конституции человека М.В. Черноруцкого
Рассмотрена классификация типов конституции человека, разработанная выдающимся терапевтом М.В. Черноруцким в 1925 году. Классификация типов конституции человека М.В.
Типы гипертрофии скелетных мышц человека
В статье дается классификация различных видов гипертрофии скелетных мышц человека на основе ряда классификационных признаков: времени проявления…
Миомейкер: Мембранный активатор слияния миобластов и образования мышц
Ученые установили, что для образования мышечных волокон необходимо слияние клеток-предшественников, которые называются миобластами. Эти клетки имеют только…
Ткани и органы. Мышцы
Регуляция сокращения мышечных волокон
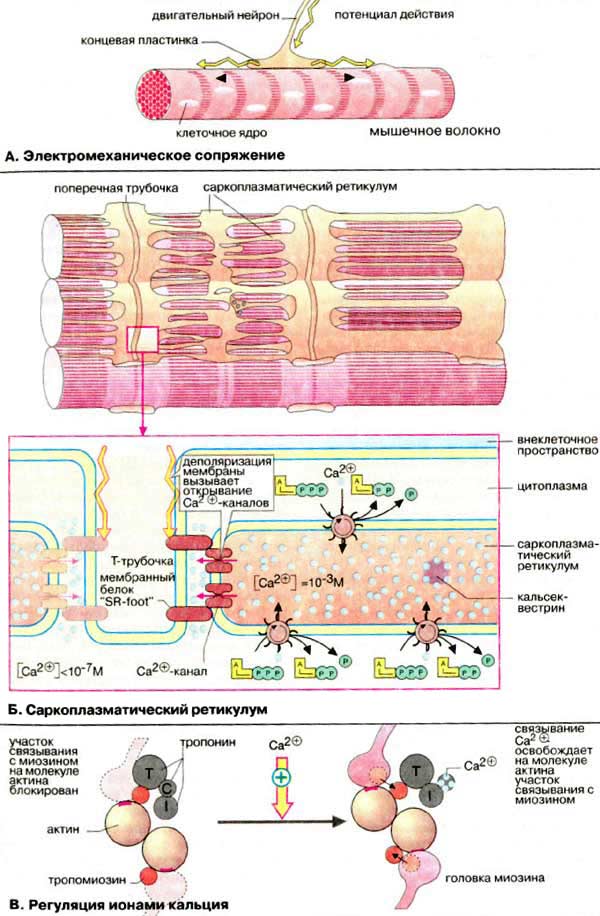
А. Электромеханическое сопряжение
Сокращением мышечного волокна управляют двигательные нейроны , которые выделяют нейромедиатор ацетилхолин в нервно-мышечные соединении (синапсы) (см. рис. 345). Ацетилхолин диффундирует через синаптическую щель и взаимодействуют с ацетилхолиновыми (холинэргическими) рецепторами плазматической мембраны мышечных клеток. Это вызывает открывание трансмембранных ионных каналов и деполяризацию клеточной мембраны (образование потенциала действия). Потенциал действия быстро распространяется по всем направлениям от нервно-мышечного соединения (см. рис. 341, 343), возбуждая все мышечные клетки. В течение нескольких миллисекунд реализуется рассмотренный выше цикл сокращения мышечного волокна.
Б. Саркоплазматический ретикулум
Саркоплазматический ретикулум [СР (SR)] -разветвленная подобная эндоплазматическому ретикулуму органелла, окружающая индивидуальные миофибриллы подобно сетке (в верхней части схемы в качестве примера приведен СР клетки сердечной мышцы). В покоящихся клетках концентрация Са 2+ очень низка (менее 10 -5 М). Однако в саркоплазматическом ретикулуме уровень ионов Са 2+ существенно выше (около 10 -3 М). Высокая концентрация Са 2+ в СР поддерживается Са 2+ -АТФ-азами. Кроме того, в СР имеется специальный белок кальсеквестрин (55 кДа), который благодаря высокому содержанию кислых аминокислот способен прочно связывать ионы Са 2+ .
Переносу потенциала действия на СР индивидуальной миофибриллы способствуют поперечные трубочки Т-системы , представляющие трубчатые впячивания клеточной мембраны и находящиеся в тесном контакте с индивидуальными миофибриллами. Деполяризация плазматической мембраны передается через Т-трубочки на потенци ал-управляемый мембранный белок (так называемый "SR-foot") прилегающей мембраны СР, который открывает Са 2+ -каналы. Результатом является выброс ионов Са 2+ из СР в пространство между филаментами актина и миозина до уровня ≥10 -5 M. В конечном итоге выброс ионов Са 2+ является пусковым механизмом процесса сокращения миофибрилл.
В. Регуляция ионами кальция
В расслабленной скелетной мышце комплекс тропонина (субъединицы = Т, С, I ) с тропомиозином препятствует взаимодействию миозиновых головок с актином .
Быстрое увеличение в цитоплазме концентрации ионов кальция в результате открывания каналов СР приводит к связыванию Са 2+ с С-субъединицей тропонина. Последняя по свойствам близка кальмодулину (см. рис. 375). Связывание ионов Са 2+ вызывает конформационную перестройку в тропонине, тропонинтропомиозиновый комплекс разрушается и освобождает на молекуле актина участок связывания с миозином (на схеме выделен красным цветом). Это инициирует цикл мышечного сокращения (см. с. 324)
В отсутствие последующего стимулирования АТФ-зависимые кальциевые насосы мембраны СР быстро снижают концентрацию ионов Ca 2+ до исходного уровня. Как следствие, комплекс Са 2+ с тропонином С диссоциирует, тропонин восстанавливает исходную конформацию, место связывания миозина на актине блокируется и мышца расслабляется.
Таким образом, при сокращении мышечного волокна скелетных мышц позвоночных происходит следующая последовательность событий. При поступлении сигнала от двигательного нейрона мембрана мышечной клетки деполяризуется, сигнал передается на Сa 2+ -каналы СР. Са 2+ -каналы открываются, внутриклеточный уровень ионов Са 2+ возрастает. Ионы Сa 2+ связывается с тропонином С, вызывая конформационную перестройку в тропонине, что влечет за собой разрушение комплекса тропонин-тропомиозин и дает возможность головкам миозина связываться с актином. Происходит инициация актин-миозинового цикла.
По завершении сокращения уровень ионов Са 2+ снижается за счет активного обратного транспорта Са 2+ в СР, тропонин С отдает Са 2+ , комплекс тропонин-тропомиозин занимает исходное положение на молекуле актина, блокируя актин-миозиновый цикл. Результатом является расслабление мышцы.
Читайте также: