
Что такое окисление в биологии 8 класс кратко
Обновлено: 05.07.2024
Совокупность реакций окисления, протекающих во всех живых клетках. Основная функция О. б. — обеспечение организма энергией в доступной для использования форме. Реакции О. б. в клетках катализируют ферменты, объединяемые в класс оксидоредуктаз (См. Оксидоредуктазы). Изучение окисления в организме было начато в 18 в. А. Лавуазье; в дальнейшем значительный вклад в исследование О. б. (его локализация в живых клетках, связь с др. процессами обмена веществ, механизмы ферментативных окислительно-восстановительных реакций, аккумуляция и превращение энергии и др.) внесли О. Варбург, Г. Виланд (Германия), Д. Кейлин, Х. Кребс, П. Митчелл (Великобритания), Д. Грин, А. Ленинджер, Б. Чанс, Э. Рэкер (США), а в СССР — А. Н. Бах, В. И. Палладин, В. А. Энгельгардт, С. Е. Северин, В. А. Белицер, В. П. Скулачев и др.
О. б. в клетках связано с передачей т. н. восстанавливающих эквивалентов (ВЭ) — атомов водорода или электронов — от одного соединения — донора, к другому — акцептору. У аэробов (См. Аэробы) — большинства животных, растений и многих микроорганизмов — конечным акцептором ВЭ служит кислород. Поставщиками ВЭ могут быть как органические, так и неорганические вещества (см. таблицу).
Классификация организмов по источнику энергии и восстанавливающих эквивалентов
| Тип организмов | Источник энергии | Окисляемое соединение (поставщик восстанавливающих эквивалентов) | Примеры |
|---|---|---|---|
| Фотолитотрофы | Свет | Неорганические соединения | Зелёные клетки высших растений, |
| Фотоорганотрофы | Свет | (Н2О, H2S, S) | синезелёные водоросли, |
| Хемолитотрофы | Реакции | Органические соединения | фотосинтезирующие бактерии |
| Хемоорганотрофы | окисления | Неорганические соединения | Несерные пурпурные бактерии |
| Реакции | (H2, S, H2S, NH3, Fe 2 + ) | Водородные, серные, | |
| окисления | Органические соединения | денитрифицирующие бактерии, | |
| железобактерии | |||
| Животные, большинство | |||
| микроорганизмов, | |||
| нефотосинтезирующие клетки | |||
| растений |
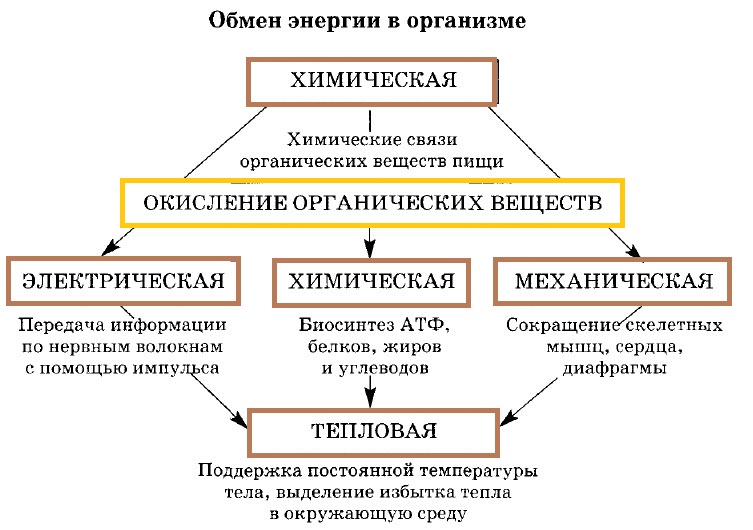
Основной путь использования энергии, освобождающейся при О. б., — накопление её в молекулах аденозинтрифосфорной кислоты (АТФ) и др. макроэргических соединений (См. Макроэргические соединения). О. б., сопровождающееся синтезом АТФ из аденозиндифосфорной кислоты (АДФ) и неорганического фосфата, происходит при Гликолизе, окислении α-кетоглутаровой кислоты и при переносе ВЭ в цепи окислительных (дыхательных) ферментов, обычно называют окислительным фосфорилированием (См. Окислительное фосфорилирование) (см. схему).
В процессе дыхания углеводы, жиры и белки подвергаются многоступенчатому окислению, которое приводит к восстановлению основных поставщиков ВЭ для дыхательных флавинов, Никотинамидадениндинуклеотида (НАД), Никотинамидадениндинуклеотидфосфата (НАДФ) и липоевой кислоты (См. Липоевая кислота). Восстановление этих соединений в значительной мере осуществляется в Трикарбоновых кислот цикле, которым завершаются основные пути окислительного расщепления углеводов (оно начинается с гликолиза), жиров и аминокислот. Помимо цикла трикарбоновых кислот, некоторое количество восстановленных коферментов (См. Коферменты) — ФАД (Флавинадениндинуклеотида) и НАД — образуется при окислении жирных кислот, а также при окислительном дезаминировании глутаминовой кислоты (НАД) и в пентозофосфатном цикле (См. Пентозофосфатный цикл) (восстановленный НАДФ).
Соотношение и локализация различных механизмов О. б. В расчёте на 1 молекулу глюкозы гликолиз даёт 2 молекулы АТФ, а фосфорилирование в дыхательной цепи — 34 молекулы АТФ. Гликолиз, цикл трикарбоновых кислот и дыхательная цепь функционируют, по-видимому, в клетках всех эукариотов (См. Эукариоты). Окисление жирных кислот у позвоночных поставляет половину энергии, потребляемой печенью, почками, мышцей сердца и покоящимися скелетными мышцами; в клетках мозга оно практически не происходит. Окисление по пентозофосфатному пути активно в печени и лактирующих молочных железах, но незначительно в сердечной и скелетных мышцах.
В жидкой фазе цитоплазмы растворены все ферменты гликолиза. Внутренние мембраны митохондрий (См. Митохондрии), мембраны хлоропластов (См. Хлоропласты) (тилакоидов) и клеточные мембраны бактерий содержат фосфорилирующие цепи переноса электронов. В матриксе митохондрий локализовано окисление жирных кислот, ферменты цикла трикарбоновых кислот и глутаматдегидрогеназа. Во внутренней мембране митохондрий находятся ферменты, окисляющие янтарную и β-оксимасляную кислоты, во внешней — ферменты, участвующие в обмене аминокислот: Моноаминоксидаза и кинуренингидроксилаза. В особых органоидах клетки, т. н. пероксисомах, или микротельцах, вклад которых в суммарное поглощение О2 может достигать в печени 20%, находится флавиновая оксидаза, окисляющая аминокислоты, гликолевую кислоту и др. субстраты с образованием перекиси водорода, которая затем разлагается каталазой (См. Каталаза) или используется пероксидазами (См. Пероксидазы) в реакциях окисления. В мембранах эндоплазматической сети клетки локализованы гидроксилазы и оксигеназы, организованные в короткие нефосфорилирующие цепи переноса электронов.
Окислительные реакции не всегда сопровождаются накоплением энергии; в ряде случаев они несут функции превращения веществ (например, окисление при образовании жёлчных кислот, стероидных гормонов, на путях превращения аминокислот и др.). При окислении происходит обезвреживание чужеродных и ядовитых для организма веществ (ароматических соединений, недоокисленных продуктов дыхания и др.). О. б., не сопряжённое с накоплением энергии, называется свободным окислением. Его энергетический эффект — образование тепла. По-видимому, система переноса электронов, осуществляющая окислительное фосфорилирование, способна переключаться на свободное окисление при увеличении потребности организма в тепле (у гомойотермных животных (См. Гомойотермные животные)).
Эволюция энергообеспечения в живой природе. Древнейшие организмы, как полагают, существовали в первичной бескислородной атмосфере Земли и были анаэробами (См. Анаэробы) и гетеротрофными организмами (См. Гетеротрофные организмы). Обеспечение клеток энергией шло за счёт процессов типа гликолиза. Возможно, существовал механизм окисления, известный у некоторых современных микроорганизмов: ВЭ передаются через дыхательную цепь на нитрат (NO – 3) или на сульфат (SO – – 4). Принципиально важным этапом оказалось возникновение у древних одноклеточных организмов механизма фотосинтеза, с которым связывают появление кислорода в атмосфере Земли. В результате стало возможным использование O2, обладающего высоким окислительно-восстановительным потенциалом, в качестве конечного акцептора электронов в дыхательной цепи. Реализация этой возможности произошла при появлении специального фермента — цитохромоксидазы (См. Цитохромоксидаза), восстанавливающей О2, и привела к возникновению биохимического дыхательного аппарата современного типа. Обеспечение энергией у всех аэробов (их клетки содержат митохондрии) основано на таком дыхании. Вместе с тем клетки сохранили ферментный аппарат гликолиза. Образуемая в ходе последнего пировиноградная кислота окисляется далее в цикле трикарбоновых кислот, который, в свою очередь, питает дыхательную цепь электронами. Т. о., эволюция энергетического обмена шла, по-видимому, по пути использования и надстройки уже имевшихся ранее механизмов энергообеспечения. Наличие в клетках ныне существующих организмов биохимических систем гликолиза (в цитоплазме), дыхания (в митохондриях), фотосинтеза (в хлоропластах), а также поразительное сходство механизмов превращения энергии в этих органеллах и в микроорганизмах нередко рассматривают как свидетельство возможного происхождения хлоропластов и митохондрий от древних микроорганизмов-симбионтов. См. также Аденозинфосфорные кислоты, Биоэнергетика, Брожение, Дыхание, Митохондрии, Фотосинтез и лит. при этих статьях.
Лит.: Ленингер А., Превращение энергии в клетке, в кн.: Живая клетка, пер. с англ., М., 1962; Скулачев В. П., Аккумуляция энергии в клетке, М., 1969; его же, Трансформация энергии в биомембранах, М., 1972; Малер Г. и Кордес Ю., Основы биологической химии, пер. с англ., М., 1970, гл. 15; Леви А., Сикевиц Ф., Структура и функции клетки, пер. с англ., М., 1971, гл. 12; Ясайтис А. А., Превращение энергии в митохондриях, М., 1973; Ленинджер А., Биохимия, пер. с англ., М., 1974.

Пути образования АТФ при хемоорганотрофном типе энергетического обмена. ФГА — 3-фосфоглицериновый альдегид; ФГК — 3-фосфоглицериновая кислота; ФЕП — фосфоенолпировиноградная кислота; ПК — пировиноградная кислота; Ацетил-КоА — ацетил-кофермент А. Количественные соотношения отдельных путей биологического окисления показаны одинарными и двойными стрелками.

В результате окисления \(1\) г белка происходит выделение \(17,2\) кДж (\(4,1\) ккал) энергии. Но в качестве источника энергии белки обычно не используются, так как они выполняют другие функции: строительную, защитную, каталитическую и т. д.
В процессе пищеварения белки пищи расщепляются под действием пищеварительных ферментов до аминокислот. Аминокислоты всасываются ворсинками тонкого кишечника и попадают в кровь, которая доставляет их к клеткам. В клетках из аминокислот синтезируются новые белки, свойственные организму человека.
В белковом обмене важную роль играет печень. Она управляет содержанием отдельных аминокислот в крови, осуществляет синтез белков плазмы крови. Одним из продуктов распада аминокислот является ядовитый аммиак. Клетки печени преобразуют аммиак в менее опасную мочевину, которая удаляется из организма с мочой и частично с потом.

Основная функция углеводов в организме — энергетическая. \(1\) г углеводов при окислении даёт \(17,2\) кДж (\(4,1\) ккал) энергии.
С пищей в наш организм поступают разные углеводы. Чаще всего это крахмал (из растительных продуктов), гликоген (из животных продуктов), сахароза, лактоза и др. Эти соединения распадаются в органах пищеварения до глюкозы, которая всасывается стенками тонкого кишечника и попадает в кровь.

Основная часть глюкозы окисляется в клетках до углекислого газа и воды, которые удаляются с выдыхаемым воздухом или с мочой. Неиспользованная глюкоза превращается в гликоген (животный крахмал) и накапливается в клетках печени и в мышцах.
В крови содержание глюкозы поддерживается на уровне \(0,10\)–\(0,15\) % . В регуляции уровня глюкозы участвуют гормоны поджелудочной железы инсулин и глюкагон. Инсулин ускоряет превращение глюкозы в гликоген, а также затормаживает его распад. Глюкагон обладает противоположным действием. Он, наоборот, способствует расщеплению гликогена и повышению уровня глюкозы в крови.
Если поджелудочная железа вырабатывает недостаточное количество инсулина, то содержание глюкозы в крови увеличивается, и это может привести к тяжёлой болезни — сахарному диабету.

Если с пищей в организм поступает слишком много углеводов, они преобразуются в жиры и накапливаются в разных органах.
Окисление жиров в два раза эффективнее окисления углеводов или белков. \(1\) г жира даёт \(38,9\) кДж (\(9,3\) ккал) энергии.
Жиры — это вещества, образованные жирными кислотами и глицерином. В органах пищеварения жиры расщепляются на составные части под влиянием ферментов поджелудочной железы и тонкого кишечника. Образовавшиеся продукты поступают в лимфатические сосуды ворсинок тонкого кишечника, а затем вместе с лимфой попадают в кровеносную систему и доставляются к клеткам.


- Окисление жиров обеспечивает энергией работу внутренних органов.
- Липиды образуют все клеточные мембраны, выполняют функции медиаторов и гормонов.
- Откладываются в запас в подкожной жировой клетчатке и сальнике, защищают органы от механических повреждений.
- Жиры плохо проводят тепло и защищают организм от перегревания и переохлаждения, способствуя поддержанию постоянной температуры тела.
Ежедневно рекомендуется употреблять \(80\)–\(100\) г разных жиров. Лишний жир запасается под кожей, но может откладываться также в печени и в кровеносных сосудах.

Органические вещества могут взаимно превращаться. Из белков образуются жиры и углеводы. Углеводы превращаются в жиры, и наоборот, источником углеводов могут стать жиры. Но заменить белки другими веществами невозможно.
Установлено, что взрослому человеку в сутки необходимо получить с пищей не менее \(1500\)–\(1700\) ккал. Причём на обеспечение процессов жизнедеятельности тратится \(15\)–\(35\) % полученной энергии, а остальная энергия тратится на поддержание постоянной температуры тела.

[Deposit Photos]
Живые организмы не могут существовать без энергии. Ее требует каждый процесс, каждая химическая реакция. Получать энергию многие живые существа, в том числе и человек, могут с пищей. Стоит детально разобраться, откуда появляется энергия, и какие реакции протекают в это время в клетках живых организмов.
Значение биологического окисления и история его исследования
В основе получения энергии лежит процесс биологического окисления. Сейчас он изучен, создана даже целая наука, занимающаяся всеми тонкостями и механизмами процесса — биохимия. Биологическое окисление — это совокупность окислительно-восстановительных превращений веществ в живых организмах. Окислительно-восстановительными называют реакции, которые протекают с изменением степени окисления атомов вследствие перераспределения электронов между ними.

[Deposit Photos]
Первые предположения ученых о том, что внутри каждого живого организма протекают сложные химические реакции, были выдвинуты в XVIII столетии. Изучением проблемы занимался французский химик Антуан Лавуазье, обративший внимание на то, что процессы горения и биологического окисления похожи друг на друга.
Ученый проследил путь кислорода, который поглощается живым организмом в процессе дыхания, и сделал вывод, что в организме происходит процесс окисления, напоминающий процесс горения, но протекающий более медленно. Лавуазье обнаружил, что молекулы кислорода (окислитель) взаимодействуют с органическими соединениями, содержащими углерод и водород. В результате происходит абсолютное превращение, при котором соединения разлагаются.
Некоторые моменты в процессе изучения проблемы оставались для ученых непонятыми:
Чтобы ответить на эти и многие другие вопросы, а также уяснить, что такое биологическое окисление, ученым потребовался не один год. К настоящему времени химиками были изучены: связь дыхания с другими процессами обмена веществ, в т.ч. процесс фосфорилирования. Кроме того, ученые исследовали свойства ферментов, катализирующих реакции биологического окисления; локализацию ферментов в клетке; механизм аккумуляции и преобразования энергии.
Здесь вы найдете безопасные эксперименты на исследование химических свойств белков, жиров и углеводов.
Биологическое окисление и его виды
При разных условиях возможны два вида биологического окисления. Многие грибки и микроорганизмы получают энергию, преобразовывая питательные вещества анаэробным способом. Анаэробное биологическое окисление – это реакция, происходящая без доступа и какого-либо участия в процессе кислорода. Такой способ получения энергии применим живыми организмами в среде, в которую не поступает воздух: в глине, под землей, в иле, на болоте, в гниющих субстанциях. Анаэробное биологическое окисление называют гликолизом.
Более сложный способ преобразования питательных веществ в энергию — аэробное биологическое окисление, или тканевое дыхание. Эта реакция осуществляется во всех аэробных организмах, использующих кислород в процессе дыхания. Аэробный способ биологического окисления невозможен без молекулярного кислорода.
Пути биологического окисления и участники процесса
Чтобы окончательно понять, что собой представляет процесс биологического окисления, следует рассмотреть его стадии.
Гликолиз — это беcкислородное расщепление моносахаридов, предшествующее процессу клеточного дыхания и сопровождающееся выходом энергии. Такая стадия является начальной для каждого организма-гетеротрофа. После гликолиза у анаэробов наступает процесс брожения.
Окисление пирувата заключается в преобразовании пировиноградной кислоты, получаемой в процессе гликолиза, в ацетилкоэнзим. Реакция происходит с помощью ферментного комплекса пируватдегидрогеназы. Локализация – кристы митохондрий.
Распад бета-жирных кислот осуществляется параллельно с окислением пирувата на кристах митохондрий. Цель – переработка всех жирных кислот в ацетилкоэнзим и постановка его в цикл трикарбоновых кислот.
Цикл Кребса: сначала ацетилкоэнзим превращается в лимонную кислоту, затем она подвергается последующим преобразованиям (дегидрированию, декарбоксилированию и регенерации). Все процессы несколько раз повторяются.
Окислительное фосфорилирование — заключительная стадия преобразования в организмах эукариот соединений. Осуществляется преобразование аденозиндифосфата в аденозинтрифосфорную кислоту. Необходимая для этого энергия поступает в процессе окисления молекул фермент-дегидрогеназа и кофермента дегидрогеназа, сформировавшихся в предыдущих стадиях. Затем энергия заключается в макроэргические связи аденозинтрифосфорной кислоты.

АТФ [Wikimedia]
Таким образом, окисление веществ осуществляется такими способами:
- отщеплением водорода от субстрата, который окисляется (процесс дегидрирования);
- отдачей субстратом электрона;
- присоединением кислорода к субстрату.
В клетках живых организмов встречаются все перечисленные типы окислительных реакций, катализируемых соответствующими ферментами — оксидоредуктазами. Процесс окисления происходит неизолированно, он связан с реакцией восстановления: одновременно происходят реакции присоединения водорода или электрона, то есть осуществляются окислительно-восстановительные реакции. Процесс окисления — это каждая химическая реакция, которая сопровождается отдачей электронов с увеличением степеней окисления (окисленный атом имеет большую степень окисления). С окислением вещества может происходить и восстановление — присоединение электронов к атомам другого вещества.
Обмен веществ — совокупность реакций пластического и энергетического обменов.
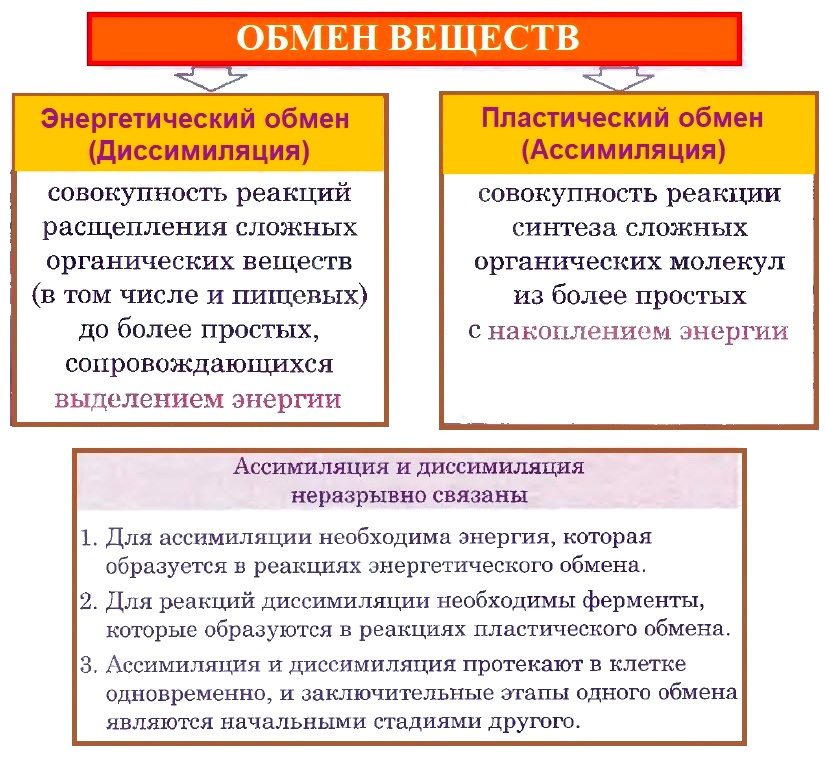
Пластический и энергетический обмен, их взаимосвязь.
Пластический обмен (ассимиляция) — совокупность реакций синтеза сложных органических веществ (белков, жиров, углеводов и нуклеиновых кислот) из более простых. Энергетический обмен (диссимиляция) — совокупность реакций, обеспечивающих клетку энергией, в ходе которых происходит расщепление и окисление сложных органических веществ.
Этапы обмена веществ:
- поступление веществ в организм;
- изменение веществ в ходе ассимиляции и диссимиляции;
- выведение конечных продуктов обмена.
Водно-минеральный обмен в организме.
Суточная потребность организма в воде в среднем составляет 2-2,5 л. Вода поступает в организм при питье (около 1 л), с пищей (около 1 л), небольшое количество (300— 350 мл) ее образуется в результате окисления органических веществ. Вода всасывается в кишечнике (тонком и толстом), ротовой полости и желудке. Из организма вода выводится с мочой (1,2-1,5 л), с потом (500-700 мл), выдыхаемым воздухом (350-800 мл), калом (100-150 мл).

Минеральные соли в организме могут быть в твердом состоянии в виде кристаллов — Са3(Р04)2 и СаСО3 в костной ткани; в диссоциированном состоянии в виде катионов и анионов. Анионы создают фосфатную буферную систему, поддерживающую внутри клеток слабокислую среду (pH 6,9), и бикарбонатную буферную систему, поддерживающую слабощелочную реакцию внеклеточной среды (pH 7,4). Общее количество минеральных солей около 4,5%. Потребности организма в них удовлетворяются продуктами питания. Железа много в яблоках, йода — в морской капусте, кальция — в молочных продуктах. Человеку необходимо постоянное поступление натрия и хлора (до 10 г поваренной соли в сутки). Всасывание солей происходит вместе с водой в толстом кишечнике. Попавшие в кровь минеральные соли доставляются клеткам. Излишки минеральных солей выводятся с мочой, потом и калом.

Обмен белков.
Суточная потребность организма в белках составляет 72-92 г. Источником белков являются преимущественно продукты животного происхождения. По содержанию аминокислоты белки делятся на полноценные (белки молока, мяса, рыбы и др.) и неполноценные, которые не содержат ни одной из незаменимых аминокислот. Особенно важны десять незаменимых аминокислот, не синтезируемых в организме (лизин, валин, лейцин, изолейцин, треонин, фенилаланин, триптофан, метионин, аргинин, гистидин).
Протеолитические ферменты расщепляют белки до полипептидов и аминокислот. Аминокислоты всасываются в кровеносные капилляры ворсинок тонкого кишечника и разносятся кровью по организму. В клетках из них образуются белки, свойственные организму. При избытке белки преобразуются в углеводы и жиры. Часть аминокислот, не использованных в синтезе белка, окисляется с освобождением энергии (17,6 кДж на 1 г вещества) и образованием воды, диоксида углерода, аммиака и др. Аммиак в печени превращается в мочевину. Продукты диссимиляции белков выводятся с мочой, потом и частично с выдыхаемым воздухом.
Обмен жиров.
Суточная потребность организма в жирах составляет 81-110 г. Животные жиры поступают в организм в виде сливочного масла, сыра, сметаны, свиного сала; растительные — в виде растительного масла. Липолитические ферменты расщепляют жиры до глицерола и жирных кислот. Жиры всасываются в лимфу, затем поступают в кровь и разносятся по всем клеткам. Часть жира, попавшего в клетки, является строительным материалом. Большая же его часть откладывается в подкожной клетчатке. При окислении 1 г жира выделяется 38,9 кДж энергии. Жиры могут синтезироваться из углеводов и белков. Конечные продукты окисления жиров — диоксид углерода и вода, удаляются с выдыхаемым воздухом, мочой, потом.
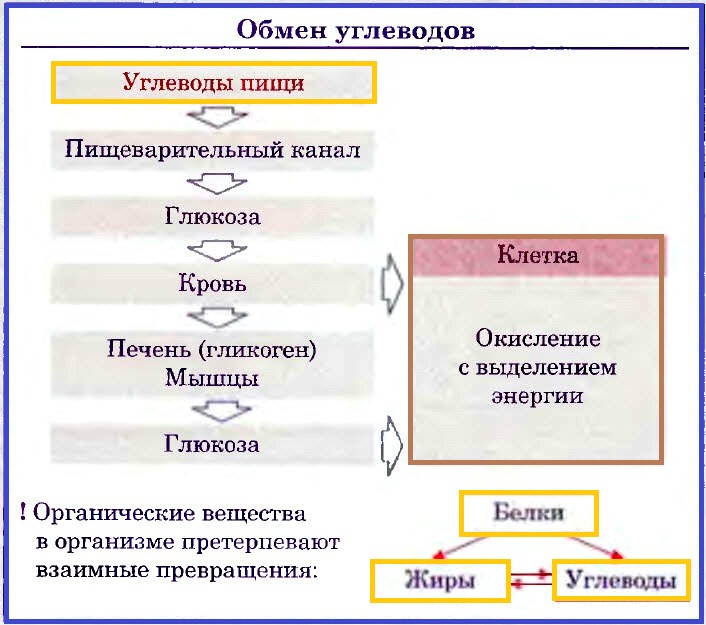
Обмен углеводов.
В сутки человек должен получать 358—484 г углеводов. Основной их источник — продукты растительного происхождения (картофель, хлеб). Углеводы в организме могут образовываться из белков и жиров. Амилолитические ферменты расщепляют углеводы до дисахаридов и моносахаридов. Моносахариды всасываются в кровеносные капилляры ворсинок кишечника и разносятся кровью по организму. Избыток глюкозы превращается в печени в гликоген. При чрезмерном поступлении углеводов они превращаются в жиры. В клетках глюкоза окисляется до диоксида углерода и воды, которые удаляются с выдыхаемым воздухом, мочой, потом, при этом выделяется энергия (17,6 кДж на 1 г глюкозы).
Читайте также: