
Что представляет собой поверхностный аппарат клетки кратко
Обновлено: 30.06.2024
Поверхностный аппарат клеток состоит из трех субсистем: плазматической мембраны, субмембранного опорносократительного аппарата гиалоплазмы и надмембранного комплекса.
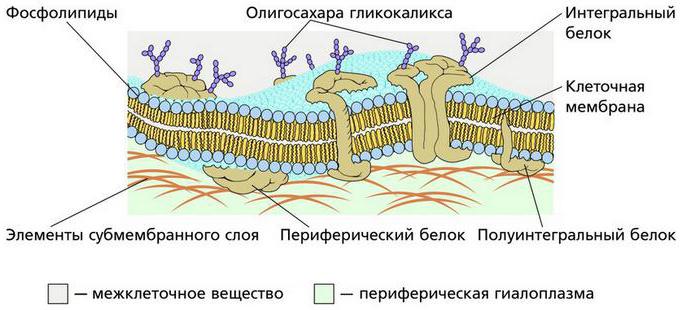
Плазматическая мембрана (рис. 1.4). Среди многочисленных моделей мембран, наиболее универсальной оказалась так называемая “жидкостно-мозаичная” модель, которая постулирует наличие гидрофобно-гидрофильных взаимодействий не только между липидными молекулами, но и между липидами и белками. В этом случае основой является жидкостный билипидный слой, образованный строго ориентированными липидными молекулами. Двойной слой липидных молекул обращен друг к другу гидрофобными участками, а внешняя и внутренняя поверхности билипидного слоя образованы гидрофильными участками молекул. Белки, входящие в мембрану, не составляют сплошного слоя на внутренней и внешней поверхности билипидного слоя. Мембранные белки представлены тремя разновидностями: периферические белки располагаются на поверхности би- липидного слоя и связаны с полярными головками липидных молекул электростатическими взаимодействиями; интегральные белки пронизывают всю толщу мембраны, причем их гидрофобная часть погружена в гидрофобную зону билипидного слоя; полуинтегральные белки погружены в мембрану лишь наполовину, выступая наружу с какой-то одной (внешней или внутренней) поверхности мембраны. Гидрофобные части белковых глобул сосредоточены на одном из полюсов.
Рис. 1.4. Мозаичная модель клеточной мембраны

Из этой модели организации мембраны вытекает важное следствие, а именно: возможность латерального и отчасти вертикального смещения интегральных и полуинтегральных белковых глобул, то есть динамичность и лабильность такой системы. Более того, мембранные белки обладают разнообразными специфическими функциями. Они выполняют рецепторные функции, являются активными и пассивными переносчиками различных соединений, наконец, входят в состав многочисленных ферментных систем.
Субмембранная система клетки. Представляет собой специализированную периферическую часть цитоплазмы и занимает, следовательно, пограничное положение между рабочим метаболическим аппаратом клетки и плазматической мембраной. В субмембранной системе поверхностного аппарата можно выделить две части: периферическую гиалоплазму, где сосредоточены ферментативные системы, связанные с процессами трансмембранного транспорта и рецепции, и структурно оформленную опорно-сократимую систему. Опорно-сократимая система состоит из микрофибрилл, микротрубочек и скелетных фибриллярных структур.
Надмембранные структуры поверхностного аппарата эукариотных клеток можно разделить на две большие категории.
1. Собственно надмембранныи комплекс, или гликокаликс. В его состав входят периферические белки мембраны, углеводные части гликолипидов и гликопротеинов. Гликокаликс играет важную роль в рецепторной функции, обеспечивает “индивидуализацию” клетки — в его составе сосредоточены рецепторы тканевой совместимости.
2. Производные надмембранных структур. К ним относятся специфические химические соединения, не производящиеся самой клеткой. Наиболее изучены они на микроворсинках клеток кишечного эпителия млекопитающих. Здесь ими являются гидролитические ферменты, адсорбирующиеся из полости кишки. Их переход из взвешенного и фиксированное состояние создает базу для качественно иного типа пищеварения, так называемого пристеночного пищеварения. Последнее по своей сути занимает промежуточное положение между полостным и внутриклеточным.
Основные функции поверхностного аппарата.
1. Трансмембранный транспорт. Представлен тремя категориями процессов:
б) пассивный транспорт, или облегченная диффузия;
в) активный транспорт, или избирательный (при участии АТФ и ферментов).
2. Рецепторная функция.
3.Транспорт в мембранной упаковке. Делится на экзоцитоз и эндоцитоз (фагоцитоз, пиноцитоз) в зависимости от того, в каком направлении переносятся вещества (в клетку или из клетки).
Библиотека образовательных материалов для студентов, учителей, учеников и их родителей.
Наш сайт не претендует на авторство размещенных материалов. Мы только конвертируем в удобный формат материалы из сети Интернет, которые находятся в открытом доступе и присланные нашими посетителями.
Если вы являетесь обладателем авторского права на любой размещенный у нас материал и намерены удалить его или получить ссылки на место коммерческого размещения материалов, обратитесь для согласования к администратору сайта.
Разрешается копировать материалы с обязательной гипертекстовой ссылкой на сайт, будьте благодарными мы затратили много усилий чтобы привести информацию в удобный вид.
Поверхностный аппарат клетки имеет сложное строение. В основе его лежит плазматическая мембрана, с которой снаружи связан надмембранный комплекс – гликокаликс, а изнутри – опорно-сократительный аппарат гиалоплазмы. Плазмолемма (plasmolemma), или внешняя клеточная мембрана, – самая толстая из цитомембран: её толщина 10 нм. Плазмолемма состоит из билипидного слоя, встроенных в него белковых молекул и гликокаликса (рисунок 3.2).
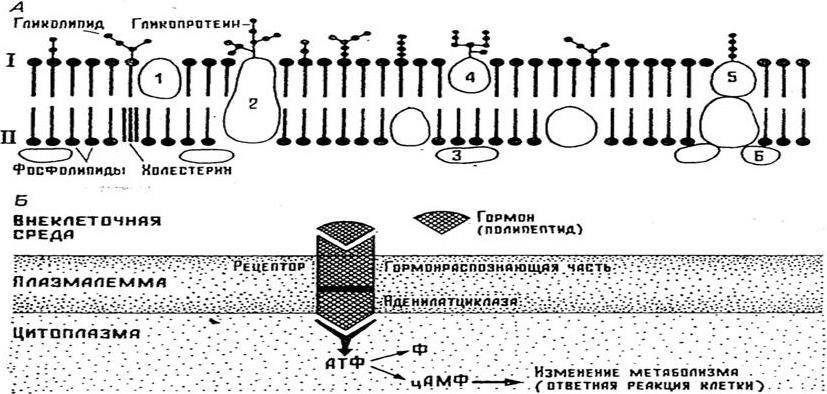
А – строение; Б – участие в рецепции: I надмембранный слой (гликокаликс); II – липопротеиновая мембрана; 1 – 6 – белки.
Рисунок 3.2 – Плазмалемма
Лежащий в основе плазмолеммы билипидный слой образуют полярные молекулы фосфолипидов (с гидрофильной головкой и гидрофобными хвостиками), а также молекулы холестерина. Билипидный слой асимметричен, почти все гликолипиды сконцентрированы в наружном монослое, в котором, кроме того, сосредоточены высокомолекулярные, более насыщенные жирные кислоты, в отличие от внутреннего слоя, в состав которого входят ненасыщенные жирные кислоты. Внутренняя сторона мембраны по отношению к наружной заряжена более отрицательно. В билипидном слое находятся различные белки: интегральные, полуинтегральные и субповерхностные. Белки обеспечивают такие функции клетки как рецепцию, регулируемый транспорт, структурную организацию процессов метаболизма и др. Интегральные белковые молекулы, прочно ассоциированные липидами, нельзя выделить из мембран, не разрушив последних, в отличие от легкоэстрагируемых периферических белков, расположенных вне билипидного слоя, но либо ковалентно связанных непосредственно с липидами, либо через олигосахарид – с фосфатидилинозитолом наружного монослоя. Интегральные белки могут быть соединены с многочисленными углеводными остатками и, по существу являться гликопротеинами. От консистенции билипидного слоя во многом зависит активность мембраны.
Среди белковых молекул плазмолеммы встречаются структурные, транспортные белки – переносчики тех или иных веществ, белки, образующие поры, или гидрофильные каналы и ферменты. Белки – переносчики электронов. Состав гликолипидов гликокаликса выделяют класс ганглиозидов, участвующих в работе химических синапсов нервных клеток. Гликолипидам принадлежит важнейшая роль в рецепторной функции мембраны. Состав гликолипидов меняется в малигнизированных клетках (клетках злокачественной опухоли). Гликолипиды эритроцитов определяют группу крови.
Важный компонент мембран животной клетки – стероидный липид холестерол, определяющий их консистенцию. Несмотря на то, что мембраны различаются по химическому составу, все они выполняют барьерную функцию и ограничивают свободную диффузию веществ.
Плазмолемма выполняет следующие функции:
1) разграничительную – отделяет содержимое клетки от внешней среды;
2) рецепторную – воспринимает из окружающей среды раздражения различной природы;
3) транспортную – регулирует обмен веществ между клеткой и окружающей средой, обладая уникальной избирательной проницаемостью.
Транспортная функция обусловлена необходимостью обеспечить в клетке оптимальное значение рН и соответствующую ионную концентрацию для эффективной работы ферментов; доставить питательные вещества, которые служат источником энергии и сырьем для синтеза собственных белков; вывести вредные продукты метаболизма (диоксид углерода, пероксид водорода, аммиак, нерастворимые соли), а также гормоны, медиаторы и другие, биологически активные вещества; создать ионный градиент для электропроводимости поверхностной мембраны, осуществления нервной деятельности. Вещества поступают в клетку и выводятся из нее различными способами: диффузия, осмос, фагоцитоз и другие (рисунок 3.3).
1 Диффузия – поступление в клетку через мембрану веществ по диффузионному градиенту из области с высокой концентрацией в область с низкой. Некоторые газы диффундируют быстро; ионы и полярные молекулы (глюкоза, аминокислоты, жирные кислоты и глицерол) – медленно. Значительно быстрее через мембрану проходят незаряженные и жирорастворимые (липофильные) молекулы.
2 Осмос – переход молекул воды по градиенту концентрации из гипотонического раствора в гипертонический. Гипо- или гипертонический раствор приводит в одном случае к пикнозу эритроцитов, а в другом – к их гемолизу, но в обоих случаях вызывает шок. Поэтому внутривенно можно вводить только изотонические растворы (0,86 % NaС1).
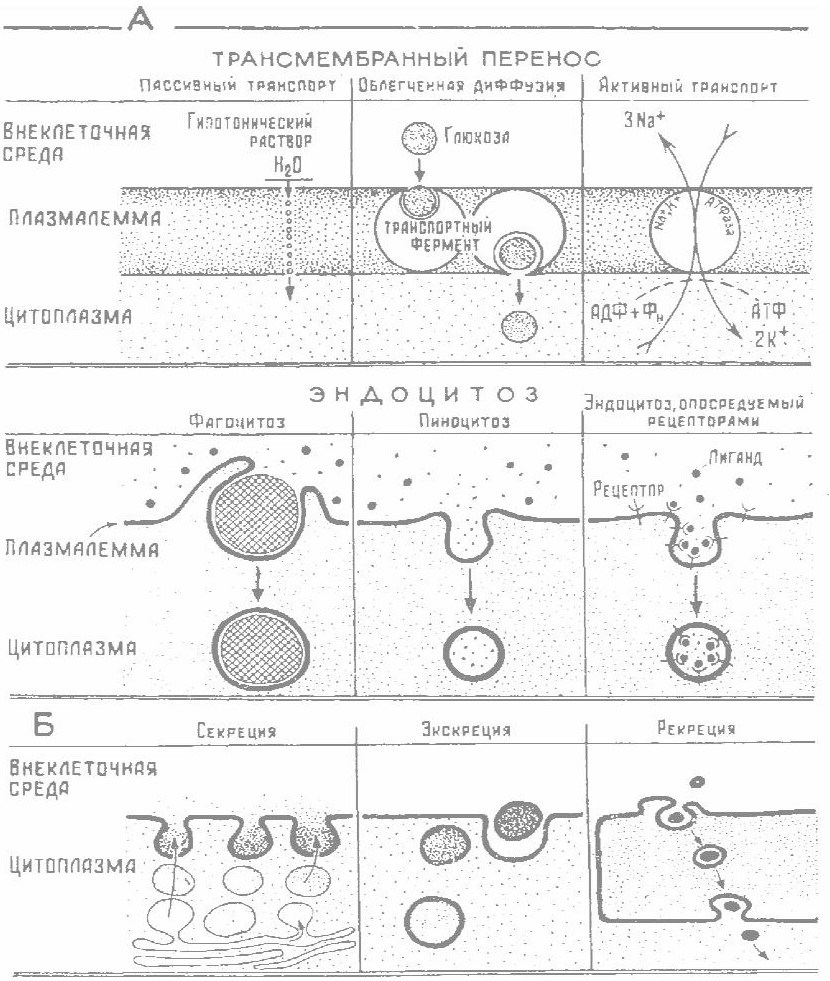
Рисунок 3.3 – Участие плазмалеммы в поступлении (А) и выведении (Б) веществ
3 Активный транспорт (связанный с затратами энергии) – это перенос молекул или ионов через мембрану по электрохимическому градиенту. Так как содержимое всех клеток заряжено отрицательно, то катионы всегда стремятся внутрь клетки, тогда как анионы отталкиваются клеткой. Внутри клеток и внеклеточной жидкости, как известно, преобладают ионы: Na + , К + и СI - , причем натрий активно выкачивается из клетки, а калий активно в нее поступает. Этот процесс называется калийнатриевым насосом (>К + Na + насосом). Для обеспечения активного транспорта организм расходует энергию, запасенную в аденозинтрифосфорную кислоту (АТФ). Работу калий-натриевого насоса, которую запускает определенный раздражитель из внешней среды, например медиатор ацетилхолин (АЦХ) и адреналин (А), можно сравнить с ключом, открывающим только определенный замок: в данном случае замком будет служить рецептор на интегральном белке. После возбуждения АЦХ домены интегрального белка раскручиваются. В нем появляется канал, ионы с внутренней стороны мембраны перераспределяются на наружную, и знак потенциала изменяется. На мембране появляется электрический ток.
4 Фагоцитоз, эндоцитоз и экзоцитоз – это активные процессы, посредством которых различные вещества транспортируются через мембрану либо в клетку, либо из нее.
Фагоцитоз – это процесс поглощения твердых частиц или крупных одноклеточных организмов, когда захват осуществляется с образованием псевдоподий. При этом в месте контакта с клеткой поглощаемого субстрата образуется крупная пищеварительная вакуоль – фагосома, которая отшнуровывается от плазматической мембраны и поступает в клетку, где сливается с первичной лизосомой. Мембраны фагосомы и лизосомы сливаются за счет слипания липидных бислоев. В результате образуются фаголизосомы, представляющие собой компартменты – специализированные структуры, предназначенные для внутриклеточного переваривания.
Эндоцитоз – процесс поглощения более мелких органических частиц. В этом случае в месте контакта поглощаемого субстрата с клеткой образуется эндоцитозный пузырек, который затем отшнуровывается от плазматической мембраны и поступает в цитоплазму клетки, где сливается с первичной лизосомой. В результате слияния образуется вторичная лизосома, в которой и переваривается субстрат.
Пиноцитоз – это процесс поглощения жидких веществ (воды, коллоидов, суспензий), протекающий на молекулярном уровне. Этим способом клетки дышат кислородом, растворенным в воде, и выделяют диоксид углерода, медиаторы и иные вещества.
Экзоцитоз – процесс, обусловливающий транспортировку веществ, заключенных в мембранную упаковку, из клетки во внешнюю среду. Таким образом, транспортируются не только продукты экскреции, но и секреторные гранулы, продуцируемые клеткой; удаляются продукты непереваренной пищи из пищеварительных вакуолей, остатки фаголизосом, миелиновые тельца.
Под плазмолеммой в периферическом слое гиалоплазмы расположена субмембранная часть поверхностного аппарата, связывающая плазматическую мембрану с цитоскелетом, представленным, как было сказано ранее, системой микрофиламентов, микротрубочек и промежуточными филаментами.
Кроме перечисленных функций плазмолемма участвует в формировании межклеточных контактов, в частности, при развитии тканевых систем.
По функциональному значению межклеточные контакты можно разделить на следующие типы: изолирующие; механические; химические; электрические;
По морфологическим признакам (по строению): простые, плотные, адгезивные пояски, десмосомы, щелевидные и контакты по типу замка, межнейронные синапсы.
Структурно-функциональная организация эукариотической клетки
I. Определение клетки, ее компоненты
Клетка - целостная элементарная система, способная к самовоспроизведению и саморегуляции метаболических процессов. Эукариотическая клетка состоит из 3-х частей: поверхностного аппарата, цитоплазмы и ядра.
II. Поверхностный аппарат клетки (рис. 1) состоит из плазмалеммы (1), надмембраннного (2) и субмембранного комплексов (3).
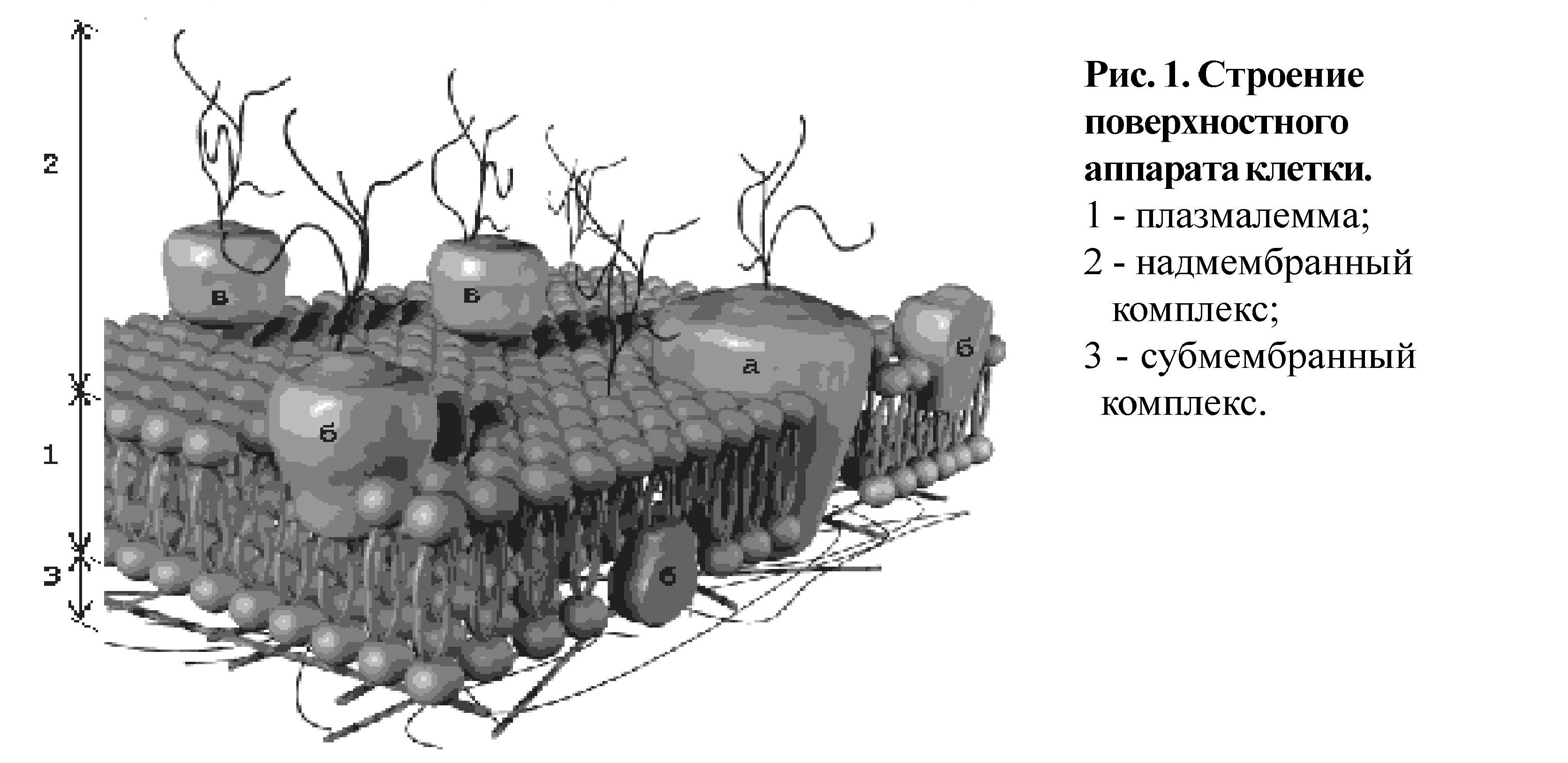
Плазмалемма (плазматическая мембрана) образована белками (-60%) и липидами (-40%). В состав липидов входят гликолипиды, стеролы, но наиболее распространены фосфолипиды. Фосфолипиды состоят из гидрофильной головки (глицерин) и гидрофобного хвоста (жирные кислоты); данная полярность липидов и позволяет образовывать в водной среде билипидный слой. По расположению в плазмалемме выделяют три вида белков: интегрГальные (а), полуинтегральные (б), периферические (в). "Белки как айсберги плавают в липидном море" по¬одиночке, но чаще сцепленными группами. Надмембранный комплекс (2), выполнен гликолипидами и гликопептидами, в животной клетке представлен гликокаликсом, в растительной - клеточной стенкой. Субмембранный слой (3) образуется путем скопления микротрубочек и микрофиламентов цитоскелета под плазмалеммой. Функции поверхностного аппарата клетки: барьерная (защитная, пограничная), обменно-транспортная, рецепторная, метаболическая, контактная (в многоклеточном организме).
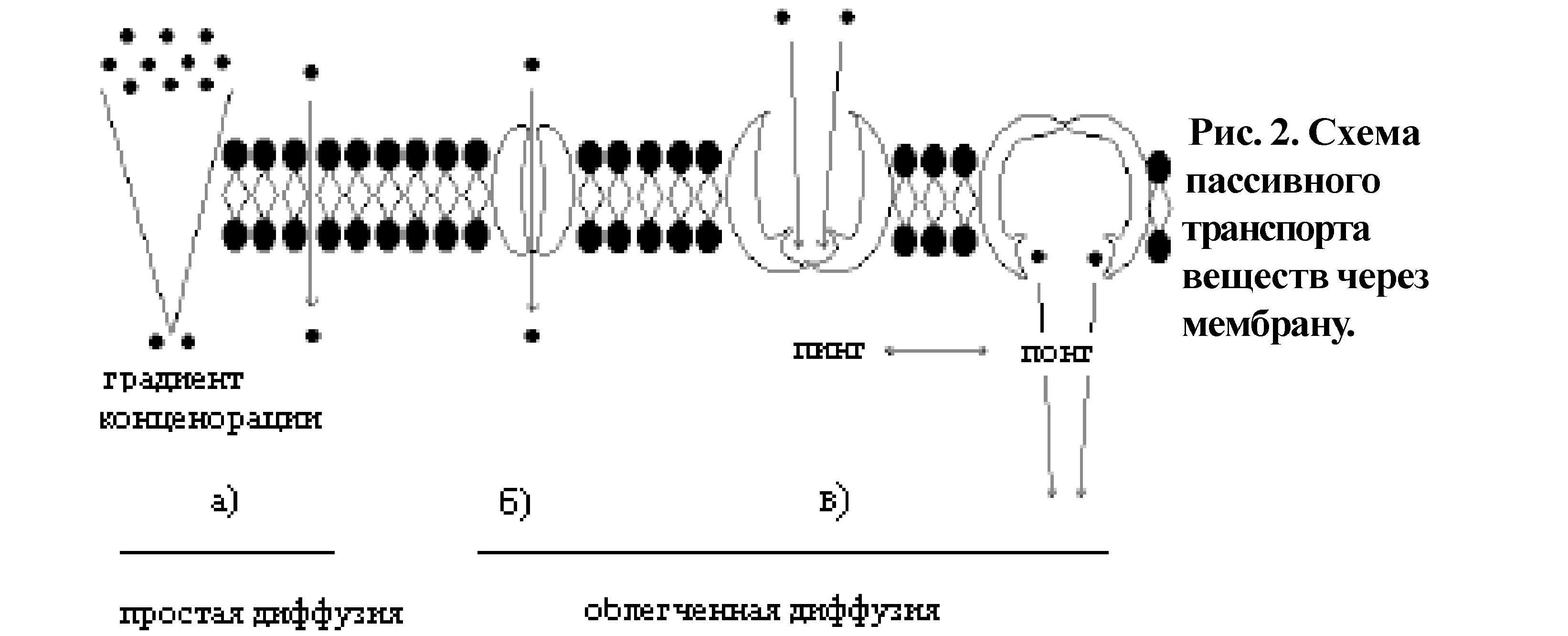
Обменно-транспортная функция. Выделяют 2 вида клеточного транспорта: пассивный и активный. Пассивный транспорт (рис.2) идет по градиенту концентрации веществ, без затраты энергии. Если вещество проходит непосредственно через билипидный слой, такой вид пассивного транспорта называется простой диффузией (так транспортируются малые неполярные молекулы - 02, С02, N2, бензол, мочевина и др.) (рис.2а), если через специфические белки - это облегченная диффузия (так транспортируются полярные молекулы. Пример: глюкоза) (рис.2б,в).
Активный транспорт (рис.З) идет против градиента концентрации веществ, с затратой энергии. Активный транспорт можно разделить на 2 вида: активный транспорт низкомолекулярных соединений (рис.З-1) /Пример: (Na++K+)-Hacoc/ и активный транспорт высокомолекулярных соединений (транспорт веществ в клетку
1. эндоцитоз, из клетки - экзоцитоз) или транспорт в мембранной упаковке;
2, на примере микропиноцитоза;
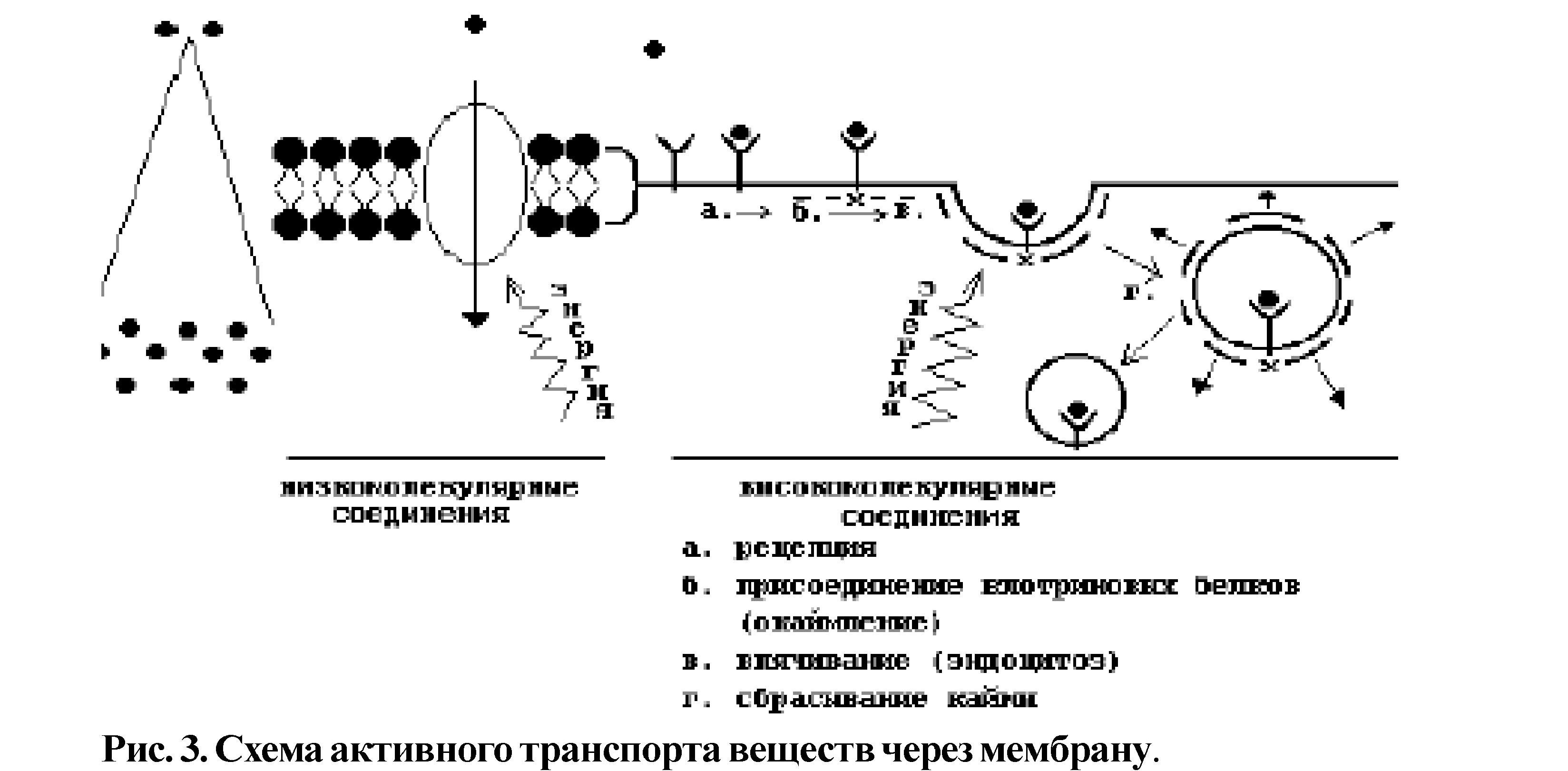
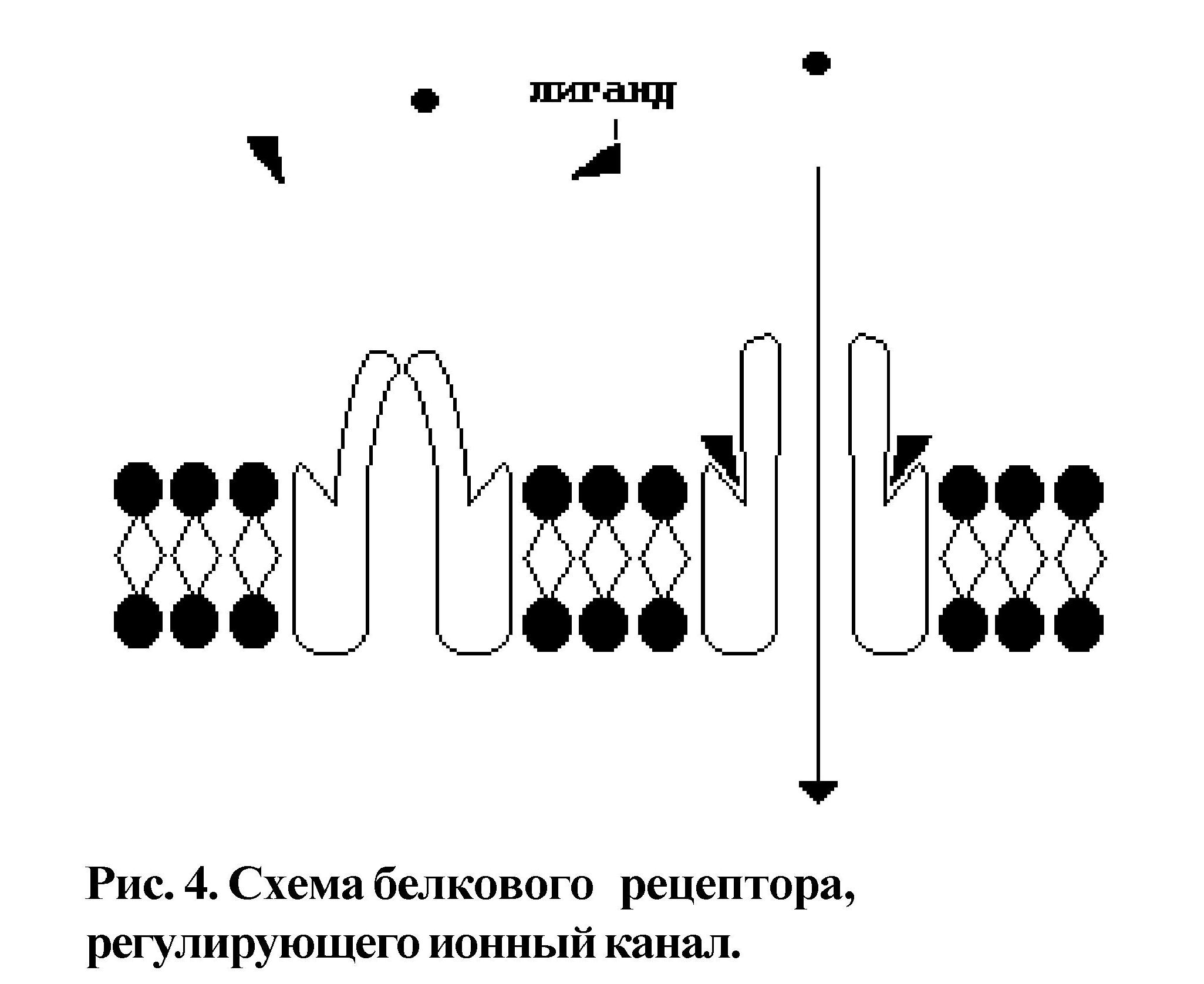
Рецепторная функция поверхностного аппарата связана с идентификацией информационного вещества - лиганда (как правило, это гормон) при помощи рецептора и адекватного ответа на данный стимул (это может быть запуск химического процесса в клетке или открытие транспортного канала) (рис.4).
III. Цитоплазма состоит из гиалоплазмы, органоидов и включений.
Гиалоплазма (цитоплазматический матрикс, цитозоль) - это внутренняя среда клетки, занимающая около 50% ее объема. По своим физико-химическим свойствам это коллоид, способный переходить из состояния геля в золь.
Гиалоплазма состоит на 90% из воды, коллоидные свойства определяются разнообразными белками. В ней содержатся также аминокислоты, полисахариды,
нуклеотиды, АТФ, жирные кислоты, витамины, растворенные газы и т.д., то есть в гиалоплазме присутствует весь разнообразный спектр веществ, необходимый клетке для процессов ее жизнедеятельности.
Органоиды - это постоянные структуры клетки. По строению можно выделить немембранные органоиды (рибосомы, центриоли, микротрубочки, филаменты. ) и мембранные, среди которых различают одномембранные (эндоплазматическая сеть, комплекс Гольджи, лизосомы, пероксисомы. ) и двухмембранные (митохондрии, пластиды).
Рибосомы. Диаметр рибосом около 20 нм. Состоят из двух субъединиц: малой и большой. В клетке эукариот два вида рибосом - 80 и 70 S (S - единицы седиментации) (табл. 1). В состав рибосом входят рРНК, рибосомальные белки, Синтез рРНК и сборка субъединиц рибосом осуществляется в ядрышке. Функции рибосом - синтез белка.
Эндоплазматическая сеть (ЭПС) - система соединенных сплющенных цистерн. Выделяют две структурно взаимосвязанные разновидности ЭПС: гладкую и гранулярную (шероховатую). Гладкая ЭПС имеет трубчатое строение, ее мембраны более контрастны (при электронной микроскопии), т. к. содержат рабочие ферменты. Функции гладкой ЭПС: компартментализация, первичный синтез липидов, синтез олигосахаридов, синтез предшественников стероидов, транспорт синтезированных веществ, детоксикация. Гранулярная ЭПС представлена уплощенными цистернами с рибосомами. Мембрана менее контрастна в сравнении с гладкой ЭПС. Функции гранулярной ЭПС: компартментализация, синтез экспортного белка, созревание белка, транспорт синтезированного белка и др.
КомплексГолъджи (КГ) состоит из дискоидных цистерн, собранных в стопки, и пузырьков по периферии. Пузырьки представляют собой формирующиеся первичные лизосомы или секреторные гранулы. При митозе КГ делится пополам, т. е. имеет преемственное строение. Функции КГ: созревание, сортировка и упаковка экспортного белка; формирование первичных лизосом и секреторных гранул; синтез полисахаридов и липидов; детоксикация; компартментализация.
Лизосомы - округлые тельца с гомогенным содержимым, окруженные мембраной. Размер лизосом 0,2-1 мкм. Содержат около 60 гидролитических ферментов (20% в мембране, 80% внутри). Функции лизосом: ауто- и гетерофагия. Пероксисомы - округлые тельца с кристаллоподобной сердцевиной. Содержат разнообразные ферменты, большинство из которых относятся к группе каталаз. Выделяют два вида пероксисом: 0,15 - 0,25 мкм - универсальные мелкие, локализуются во всех клетках; 0,3 -1,5 мкм - крупные (в клетках печени, почек). Пероксисомы участвуют в метаболизме Н202, которая используется для последующего окисления разнообразных веществ.
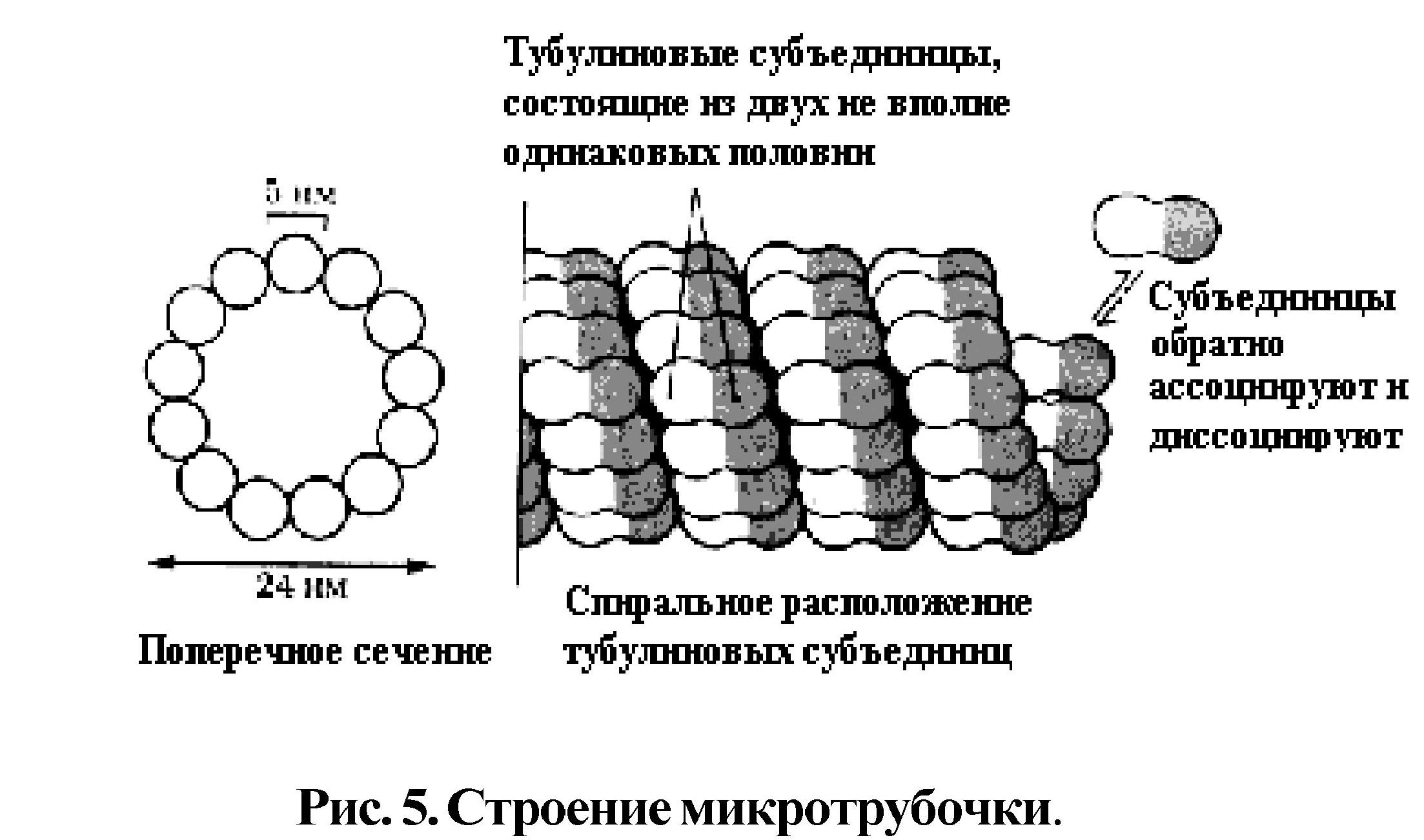
Цитоскелет включает опорные органоиды - микротрубочки, микрофиламенты, промежуточные филаменты. Микротрубочки (рис. 5) - полый цилиндр диаметром 24 нм, стенка которого построена из спирально упакованных субъединиц белка тубулина. Растут микротрубочки путем добавления с одного конца тубулиновых субъединиц. Кроме того, микротрубочки являются структурными компо-нентами центриолей, ресничек, жгутиков, базальных телец, митотического веретена.
Микрофиламенты - белковые нити диаметром 5 - 7 нм состоят из актина и миозина.
Микрофиламенты обеспечивают двигательные функции
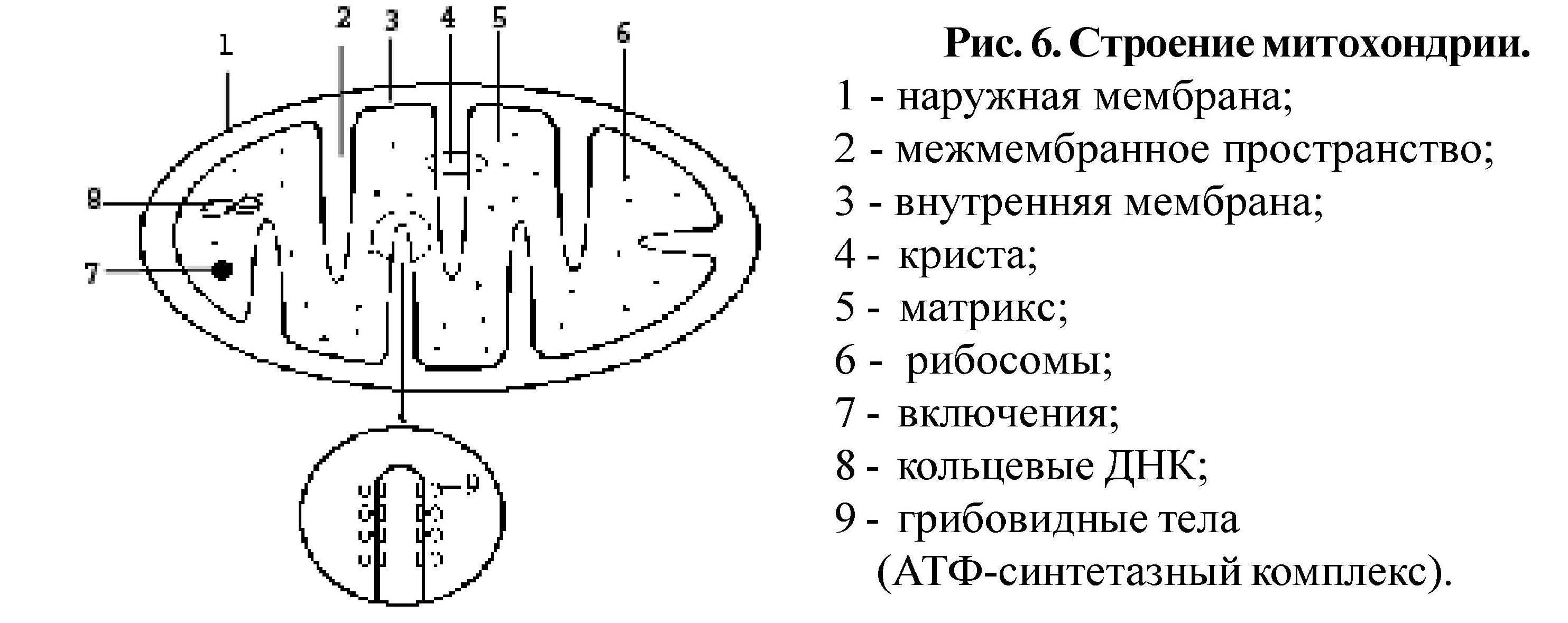
Существуют и другие органоиды, имеющие свое специфическое строение и функции.
Литическая система клетки – расщепление макромолекул внутри клетки (рис.7).
Выделяют 2 вида литических циклов: аутофагический и гетерофагический. Гетерофагический цикл - осуществляется гидролиз макро-молекул, поступивших в клетку путём эндоцитоза (фаго- и пиноцитоз).
Аутофагический цикл - расщепляются собственные клеточ-ные макромолекулы; является важнейшим элементом внутри-клеточной физио-логической регене-рации.
Включения, в отличие от органоидов, непостоянные структуры; их наличие зависит от метаболического состояния клетки. Можно выделить следующие группы включений: трофические - выполняют функцию запаса питательных веществ; секреторные - содержат вещества, выделяемые клеткой; специфические (пигментные) - характерны для специализированных клеток.
IV. Ядро (нуклеус) - наследственный аппарат эукариотической клетки содержит генетическую информацию. Форма ядра, как правило,
округлая, но может быть разнообразной, что зависит от формы клетки и ее функционального состояния. В структуре ядра выделяют следующие компоненты: поверхностный аппарат, кариоплазму, ядерный матрикс, хроматин, ядрышко. Поверхностный аппарат ядра, его строение см. рис. 8. Поровые комплексы занимают площадь от 10 -12% поверхности ядра и более, что зависит от его активности, и состоят из 3-х рядов глобулярных белков, часто встречается центральная глобула. Глобулярные белки соединены фибриллярными. Функция поровых комплексов: вывод из ядра в цитоплазму мРНК, а также ее созревание; выход субъединиц рибосом; проведение в ядро из цитоплазмы рибосомальных, гистоновых белков, ферментов репликации и транскрипции, а также нуклеотидов. Ламина (плотная пластинка) тесно связана с конденсированным хроматином, в связи с чем, кроме поддержания “архитектуры ядра", участвует в пространственной организации хроматина
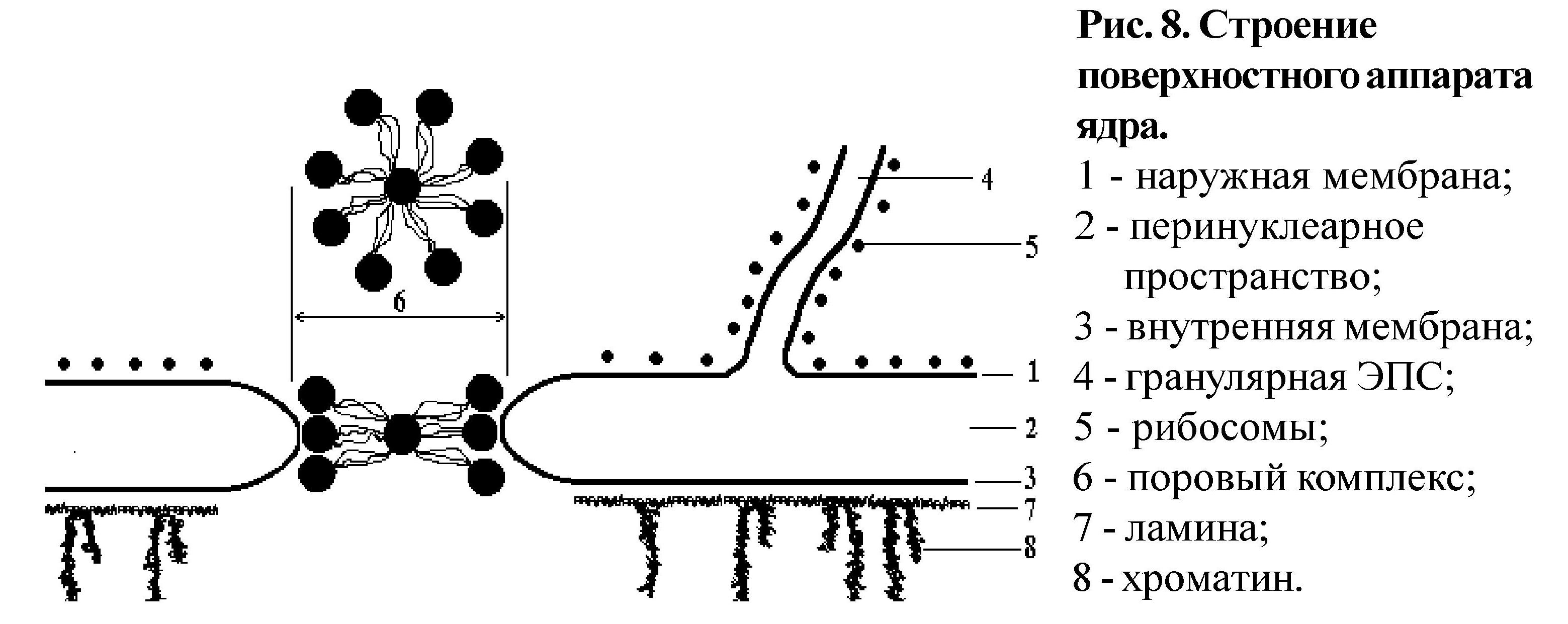
Кариоплазма (ядерный сок) - внутренняя бесструктурная среда ядра, по своим физико-химическим свойствам это белковый коллоид, в котором располагаются все структуры ядра. Основу ядерного матрикса составляют фибриллярные белки, создающие "скелет" ядра или участвующие в репликации и транскрипции.
Хроматин по химической структуре - это дезоксинуклеопротеид, состоящий из ~ 40% ДНК, ~ 60% белка (~85% гистоновые, ~15% негистоновые) и ~1% РНК. В интерфазном ядре выделяют два вида хроматина: эу- и гетерохроматин. Эухроматин - деспирализованный хроматин, активный, т. е. обладает высоким уровнем синтетических процессов. Гетрохроматин спирализован (конденсированный), неактивный.
Ядрышко - несамостоятельная и непостоянная структура ядра, которая формируется в области вторичных перетяжек хромосом (у человека 13, 14, 15, 21, 22 хромосомы), где располагаются гены, кодирующие рРНК. Функция ядрышка: синтез рибосомальных РНК и сборка субъединиц рибосом.
V. Концепция мембранного строения клетки. Мембраны клетки представлены плазмалеммой поверхностного аппарата и цитоплазматическими (внутриклеточными) мембранами. Все мембраны имеют принципиально одинаковое строение. На мембранах протекают все основные процессы в клетке, они структурируют биохимические процессы. Мембраны выполняют функцию компартментализации (компартмент – отсек, окруженная мембраной область) - пространственное разграничение внутриклеточного содержимого на обособленные отсеки, что создает условия для одновременного протекания различных ферментативных реакций. Мембранный конвейер или поток дифференцирующихся мембран - система рециклирования мембран. Мембраны формируются на основе предшествующих структур и возможен взаимопереход одних мембран в другие. Главное место сборки мембран - наружная мембрана поверхностного аппарата ядра и ЭПС в дифференцированных клетках. Перестройка мембран происходит в очень короткие сроки путем встраивания специфических белков соответствующих органоидов и изменения липидного состава.
Поверхностный аппарат клетки представляет собой универсальную субсистему. Им определяется граница между внешней средой и цитоплазмой. ПАК обеспечивает регуляцию их взаимодействия. Рассмотрим далее особенности структурно-функциональной организации поверхностного аппарата клетки.
Компоненты
Выделяют следующие составляющие поверхностного аппарата клеток эукариот: плазматическую мембрану, надмембранный и субмемранный комплексы. Первая представлена в виде сферически замкнутого элемента. Плазмолемма считается основой поверхностного клеточного аппарата. Надмембранный комплекс (его именуют также гликокаликсом) – это наружный элемент, расположенный над плазматической мембраной. В его состав входят различные компонеты. В частности, к ним относятся:
- Углеводные части гликопротеидов и гликолипидов.
- Мембранные периферические белки.
- Специфические углеводы.
- Полуинтегральные и интегральные белки.
Субмембранный комплекс расположен под плазмолеммой. В его составе выделяют опорно-сократительную систему и периферическую гиалоплазму.
Элементы субмембранного комплекса
Рассматривая строение поверхностного аппарата клетки, следует отдельно остановиться на периферической гиалоплазме. Она является специализированной цитоплазматической частью и располагается над плазмолеммой. Периферическая гиалоплазма представлена в виде жидкого высоко дифференцированного гетерогенного вещества. В нем содержатся разнообразные высоко- и низкомолекулярные элементы в растворе. Фактически она представляет собой микросреду, в которой протекают специфические и общие метаболические процессы. Периферическая гиалоплазма обеспечивает выполнение множества функций поверхностного аппарата.
Опорно-сократительная система
Она располагается в периферической гиалоплазме. В опорно-сократительной системе выделяют:
- Микрофибриллы.
- Скелетные фибриллы (промежуточные филамента).
- Микротрубочки.
Микрофибриллы представляют собой нитевидные структуры. Скелетные фибриллы формируются вследствие полимеризации ряда белковых молекул. Их количество и длина регулируется специальными механизмами. При их изменении возникают аномалии клеточных функций. Наиболее удалены от плазмалеммы микротрубочки. Их стенки образованы белками тубулинами.
Строение и функции поверхностного аппарата клетки
Обмен веществ осуществляется за счет наличия транспортных механизмов. Строение поверхностного аппарата клетки обеспечивает возможность осуществлять перемещение соединений несколькими способами. В частности, осуществляются следующие виды транспорта:
- Простая диффузия.
- Пассивный транспорт.
- Активное перемещение.
- Цитоз (обмен в мембранной упаковке).
Кроме транспортной, выявлены такие функции поверхностного аппарата клетки, как:
- Барьерная (разграничительная).
- Рецепторная.
- Опознавательная.
- Функция движения клетки с помощью образования фило-, псевдо- и ламеллоподий.
Свободное перемещение
Простая диффузия через поверхностный аппарат клетки осуществляется исключительно при наличии по обеим сторонам мембраны электрического градиента. Его размер определяет скорость и направление перемещения. Билипидный слой может пропускать любые молекулы гидрофобного типа. Однако большая часть биологически активных элементов гидрофильны. Соответственно, их свободное перемещение затруднено.
Пассивный транспорт
Этот вид перемещения соединения называют также облегченной диффузией. Она также осуществляется через поверхностный аппарат клетки при наличии градиента и без расхода АТФ. Пассивный транспорт идет быстрее, чем свободный. В процессе увеличения разности концентраций в градиенте наступает момент, в который скорость перемещения становится постоянной.
Переносчики
Транспорт через поверхностный аппарат клетки обеспечивают специальные молекулы. С помощью этих переносчиков по градиенту концентрации проходят крупные молекулы гидрофильного типа (аминокислоты, в частности). Поверхностный аппарат клетки эукариот включает в себя пассивных переносчиков для разнообразных ионов: К+, Na+, Са+, Cl-, НСО3-. Эти специальные молекулы отличаются высокой избирательностью относительно транспортируемых элементов. Кроме этого, важным их свойством является большая скорость перемещения. Она может достигать 104 и более молекул в секунду.
Активный транспорт
Он характеризуется перемещением элементов против градиента. Молекулы транспортируются из области с низкой концентрацией в участки с более высокой. Такое перемещение предполагает определенные затраты АТФ. Для осуществления активного транспорта в строение поверхностного аппарата животной клетки включены специфические переносчики. Они получили название "помп" или "насосов". Многие из этих переносчиков отличаются АТФ-азной активностью. Это означает, что они способны расщеплять аденозинтрифосфат и извлекать энергию для своей деятельности. Активный транспорт обеспечивает создание градиентов ионов.
Цитоз
Этот метод используется для перемещения частиц разных веществ либо крупных молекул. В процессе цитоза транспортируемый элемент окружается мембранным пузырьком. Если перемещение осуществляется в клетку, то его именуют эндоцитозом. Соответственно, обратное направление называется экзоцитозом. В некоторых клетках элементы проходят сквозь. Такой вид транспорта называется трансцитозом или диациозом.
Плазмолемма
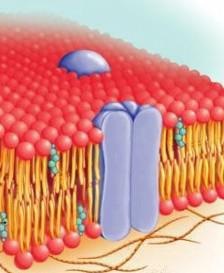
Структура поверхностного аппарата клетки включает в себя плазматическую мембрану, образованную преимущественно липидами и белками в соотношении приблизительно 1:1. Первая "бутербродная модель" этого элемента была предложена в 1935 г. В соответствии с теорией, основу плазмолеммы формируют липидные молекулы, уложенные в два слоя (билипидный слой). Они обращены хвостами (гидрофобными участками) друг к другу, а наружу и внутрь – гидрофильными головками. Эти поверхности билипидного слоя покрывают белковые молекулы. Данная модель была подтверждена в 50-х годах пошлого столетия ультраструктурными исследованиями, проведенными с использованием электронного микроскопа. Было , в частности, установлено, что поверхностный аппарат животной клетки содержит трехслойную мембрану. Ее толщина составляет 7.5-11 нм. В ней присутствует средний светлый и два темных периферических слоя. Первый соответствует гидрофобной области липидных молекул. Темные участки, в свою очередь, представляют собой сплошные поверхностные слои белка и гидрофильные головки.
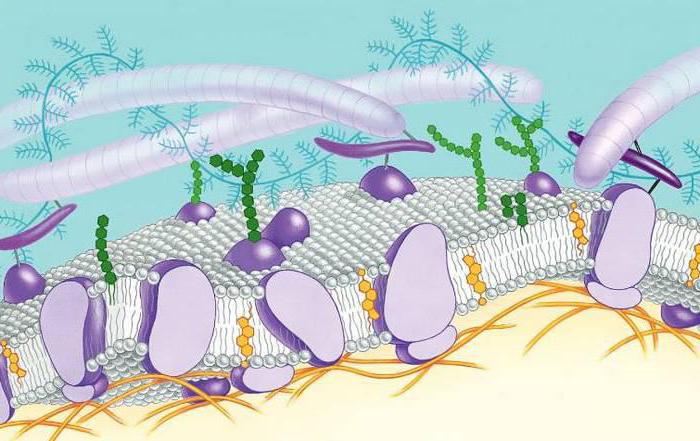
Другие теории
Разнообразные электронно-микроскопические исследования, проведенные в конце 50-х – начале 60-х гг. указывали на универсальность трехслойной организации мембран. Это нашло отражение в теории Дж. Робертсона. Между тем, к концу 60-х гг. накопилось довольно много фактов, которые не были объяснены с точки зрения существовавшей "бутербродной модели". Это дало толчок к разработке новых схем, в числе которых были модели, базирующиеся на наличии гидрофобно-гидрофильных связях белковых и липидных молекул. Среди одной из них была теория "липопротеинового коврика". В соответствии с ней, в составе мембраны присутствуют белки двух видов: интегральные и периферические. Последние связываются электростатическими взаимодействиями с полярными головками на липидных молекулах. Однако при этом они никогда не формируют сплошного слоя. Ключевая роль в формировании мембраны принадлежит глобулярным белкам. Они погружаются в нее частично и именуются полуинтегральными. Перемещение этих белков осуществляется в липидной жидкой фазе. За счет этого обеспечивается лабильность и динамичность всей мембранной системы. В настоящее время эта модель считается наиболее распространенной.
Липиды
Ключевые физико-химические характеристики мембраны обеспечиваются слоем, представленным элементами - фосфолипидами, состоящими из неполярного (гидрофобного) хвоста и полярной (гидрофильной) головки. Наиболее распространенными из них считаются фосфоглицериды и сфинголипиды. Последние сосредотачиваются главным образом в наружном монослое. Они имеют связь с олигосахаридными цепями. За счет того, что звенья выступают за пределы наружной части плазмолеммы, она приобретает асимметричную форму. Гликолипиды выполняют важную роль при осуществлении рецепторной функции поверхностного аппарата. В составе большинства мембран также находится холестерол (холестерин) - стероидный липид. Его количество различно, что, в значительной степени определяет жидкостность мембраны. Чем больше будет холестерола, тем она выше. Уровень жидкостности также зависит от соотношения ненасыщенных и насыщенных остатков от жирных кислот. Чем их больше, тем она выше. Жидкостность влияет на активность ферментов в мембране.
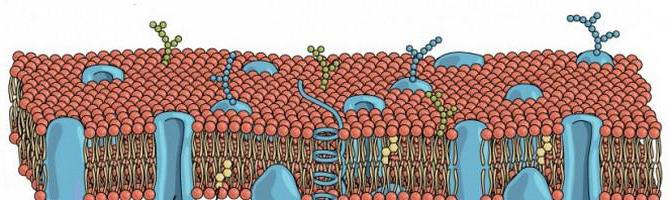
Белки
Липиды определяют главным образом барьерные свойства. Белки, в отличие от них, способствуют выполнению ключевых функций клетки. В частности, речь о регулируемом транспорте соединений, регуляции метаболизма, рецепции и так далее. Белковые молекулы распределяются в липидном бислое мозаично. Они могут перемещаться в толще. Это движение контролируется, по всей видимости, самой клеткой. В механизме перемещения задействованы микрофиламенты. Они прикрепляются к отдельным интегральным белкам. Мембранные элементы различаются в зависимости от своего расположения по отношению к билипидному слою. Белки, таким образом, могут быть периферическими и интегральными. Первые локализуются вне слоя. Они имеют непрочную связь с мембранной поверхностью. Интегральные белки полностью в нее погружены. Они имеют прочную связь с липидами и не выделяются из мембраны без повреждения билипидного слоя. Белки, которые пронизывают ее насквозь, именуются трансмембранными. Взаимодействие между белковыми молекулами и липидами разной природы обеспечивает устойчивость плазмалеммы.
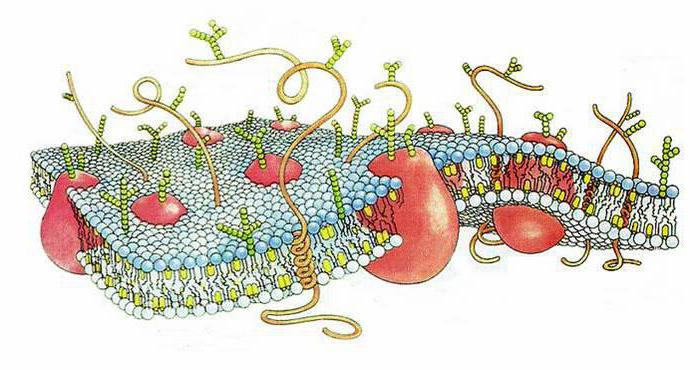
Гликокаликс
Липопротеины имеют боковые цепи. Олигосахаридные молекулы могут связываться с липидами и образовывать гликолипиды. Их углеводные части совместно с аналогичными элементами гликопротеинов придают клеточной поверхности отрицательный заряд и формируют основу гликокаликса. Он представлен рыхлым слоем с электронной умеренной плотностью. Гликокаликс покрывает наружную часть плазмолеммы. Его углеводные участки способствуют распознаванию соседних клеток и вещества между ними, а также обеспечивает адгезивные связи с ними. В гликокаликсе присутствуют также рецепторы гормонов и гитосовместимости, ферменты.
Дополнительно
Мембранные рецепторы представлены преимущественно гликопротеинами. Они обладают способностью устанавливать высокоспецифические связи с лигандами. Рецепторы, присутствующие в мембране, кроме этого, могут регулировать движение некоторых молекул внутрь клетки, проницаемость плазмалеммы. Они способны превращать сигналы внешней среды во внутренние, связывать элементы межклеточного матрикса и цитоскелет. Некоторые исследователи считают, что в состав гликокаликса также включаются полуинтегральные белковые молекулы. Их функциональные участки располагаются в надмембранной области поверхностного клеточного аппарата.
Читайте также: