
Генетика количественных признаков реферат
Обновлено: 06.07.2024
Генетика количественных признаков предоставляет психогенетике общую теорию, на базе которой строится методологический аппарат изучения природы индивидуальных психологических различий.
В самом общем виде генетика количественных признаков — применительно к психологическим задачам — исходит из того, что люди отличаются друг от друга по ряду сложных психологических признаков, и предлагает модель, в рамках которой межиндивидуальные различия по этим признакам могут быть описаны в терминах фенотипи-ческой дисперсии признака в популяции, а сама фенотипическая дисперсия может быть разложена на составляющие ее генетические и средовые компоненты.
МОДЕЛЬ ОДНОГО ГЕНА
Количественные генетические модели позволяют описать измеряемые эффекты различных генотипов, возможных в отдельно взятом локусе, и суммировать эффекты всех локусов, контролирующих тот или иной поведенческий признак. Причем количество локусов, контролирующих данный признак, обычно неизвестно, и чаще всего ученые делают допущение о том, что генетический контроль большинства поведенческих признаков осуществляется большим количеством генов, вклад которых в дисперсию изучаемого признака примерно одинаков. Одним из характерных признаков количественных генетических моделей является то, что они предполагают существование нормально распределенных фенотипических значений признаков, контролируемых множеством генов, эффекты которых, в свою очередь, опосредованы средовыми влияниями. Многолетние психологические исследования показали, что распределение большинства поведенческих признаков действительно соответствует нормальной кривой. Поэтому допущение о нормальности распределения признака, контролируемого большим количеством генов и значимых средовых влияний (т.е. являющегося мультифакторным), — психологически адекватная и статистически удобная модель для психогенетики. Важно заметить, что ожидаемая от полигенной системы нормальность распределения, будучи статистически удобной и эмпирически оправданной, не зависит от количества генов, контролирующих эту систему. Как статистические характеристики, так и теоретические положения, лежащие в основе моделей количественной генетики, одинаково правомерны для моделей, содержащих 1, 2, 25 или более генов. Именно поэтому мы начнем изложение основных признаков количественной генетики с рассмотрения модели одного гена и только потом перейдем к модели множественных генов (так называемой полигенной модели).
Конец страницы №184
Начало страницы №185

Рис. 8.1. Соотношение трех генотипов, возможных для двуаллельной системы.
Параметры dnh представляют собой разницы эффектов генов А-а. Аа может находиться на любой из сторон от т, что, соответственно, будет влиять на знак h. В примере, показанном на рисунке, значение h положительно [253, 342].
Генотипическое значение. Генотипическим значением называется некоторое количественное значение, приписываемое определенному генотипу. Так, в рамках простейшей двуаллельной системы (А и а) существуют два параметра, определяющие измеряемые эффекты трех возможных генотипов (АА, Аа и аа). Этими параметрами являются параметр d, представляющий собой удвоенную разницу между гомозиготами АА и аа, и параметр И, определяющий измеряемый эффект гетерозиготы Аа таким образом, что он не является точным усредненным эффектом двух гомозигот. Средняя точка между двумя гомозиготами, точка т, отражает среднее эффектов двух гомозиготных генотипов. Параметры d и И называются эффектами генотипов. Графически соотношение трех генотипов показано на рис. 8.1. Если в локусе отсутствует доминантность, то h будет равняться нулю, а значение генотипа Аа будет соответствовать значению в точке т. При полной доминантности значение Аа будет равняться значению АА. Если же доминантность А частична, то Аа будет находиться ближе к точке АА (или аа, в зависимости от направления доминантности) и значение h будет положительным.
Конец страницы №185
Начало страницы №186
Модели генетики количественных признаков, во всяком случае в их классическом варианте, не являются ни средством идентификации конкретных генов, контролирующих вариативность признака, ни средством точного определения вклада каждого генотипа. Эти модели решают другую задачу, а именно задачу определения общего вклада генотипа в вариативность изучаемого признака в популяции.
Теперь допустим, что каждый аллель генотипа имеет некоторый средний эффект. В этом смысле аддитивное генотипическое значение
 |
представляет собой сумму средних эффектов каждого аллеля для всех аллелей, входящих в генотип. Каждый аллель характеризуется определенным аддитивным эффектом, соответственно, при унаследовании определенного аллеля от родителя ребенок наследует и аддитивный эффект этого аллеля, т.е. вклад аллеля в генотип ребенка будет таким же, ка-
Рис. 8.2. При отсутствии доминант- ким был ег0 (g^e^) вклад в ге_
ности (п = 0) аддитивное генотипи- нотип родителя. И неважно,
ческое значение определяется генной сколько (много или мало) ajme.
дозои- лей присутствует в данном локу-
Конец страницы №186
Начало страницы №187

Рис. 8.3. Графическое изображение доминантных отклонений.
Наличие доминантности вносит разного рода изменения в аддитивное генотипическое значение.
Обозначения: • — аддитивные генотипические значения при условии полной доминантности; * — наблюдаемые аддитивные значения, определяемые на основе генных доз; D — доминантные отклонения, которые представляют собой разницу между ожидаемыми аддитивными значениями, соответствующими ситуации полной доминантности, и значениями, наблюдаемыми для определенных генных доз.
се или сколько локусов вовлечено в контроль вариативности по тому или другому признаку. Иными словами, аддитивное генотипическое значение представляет собой не что иное, как сумму вкладов каждого аллеля в генотип.
Доминантные отклонения. Доминантные отклонения есть мера того, насколько генотип отличается от своего ожидаемого аддитивного значения.
Доминантные отклонения (рис. 8.3) — это разница между ожидаемыми и наблюдаемыми значениями генотипов. Феномен доминантности допускает, что два аллеля одного локуса могут взаимодействовать друг с другом и тем самым менять генотипическое значение, которое наблюдалось бы в том случае, если бы они были независимы друг от друга и делали независимые вклады в генотипическое значение. Так, в результате взаимодействия аллелей Аа наблюдаемое генотипическое значение меньше того, которое ожидалось бы при условии полной доминантности. Напротив, значения ААи аа выше ожидаемых при допущении, что аллель А полностью доминантен по отношению к аллелю а.
Доминантность обязана своим возникновением уникальному сочетанию аллелей в данном локусе. Очевидно, что генотип потомка, наследующего только один аллель от каждого из родителей, в подавляющем большинстве случаев не может воспроизвести уникальность генотипа одного из них. Поэтому потомки будут отличаться от своих родителей в той мере, в какой аллели данного локуса не суммируются линейным образом при определении генотипического значения.
Конец страницы №187
Начало страницы №188
Рассмотрев типы генетических влияний, определим, как частоты встречаемости аллелей, определяющие эти типы, задают среднее значение генотипа в популяции. Допустим, что в популяции аллели А и а встречаются с частотами р и д, соответственно. Тогда первая колонка в табл. 8.1 показывает три возможных генотипа, вторая — частоты их встречаемости в популяции (при допущении, что особи в этой популяции образуют родительские пары случайным образом) и третья — значение генотипа. Популяционное среднее получается путем умножения значений генотипа на частоту встречаемости тех аллелей, которые этот генотип составляют, и последующего суммирования значений все трех генотипов.
Таблица 8.1 Определение среднего значения генотипа в популяции
| Генотип | Частота | Значение | Частота х значение |
| АА Аа Аа | Р 2 2pq q 2 | +d h -d | рЧ 2pqh -q4 |
| 2 =d(p 2 -q 2 ) +2hpq |

Организация стока поверхностных вод: Наибольшее количество влаги на земном шаре испаряется с поверхности морей и океанов (88‰).
Механическое удерживание земляных масс: Механическое удерживание земляных масс на склоне обеспечивают контрфорсными сооружениями различных конструкций.
Полимерное и полигенное действие генов. Понятие об аддитивных генах. Практическое значение коэффициентов наследуемости и повторяемости, методы их вычисления с помощью коэффициентов связи и дисперсионного анализа. Взаимодействие генотипа и среды.
| Рубрика | Биология и естествознание |
| Вид | учебное пособие |
| Язык | русский |
| Дата добавления | 03.07.2013 |
| Размер файла | 595,6 K |
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
HTML-версии работы пока нет.
Cкачать архив работы можно перейдя по ссылке, которая находятся ниже.
Подобные документы
Формы взаимодействия аллельных генов: полное и неполное доминирование; кодоминирование. Основные типы взаимодействия неаллельных генов: комплементарность; эпистаз; полимерия; гены-модификаторы. Особенности влияния факторов внешней среды на действие генов.
курсовая работа [601,5 K], добавлен 21.09.2010
Понятие и принципы биологии как научного направления, история ее развития и значение. Значение в организме ДНК и РНК, описание их свойств и структуры. Исследование свойств генов и развитие генетики, сферы практического применения современных достижений.
контрольная работа [26,7 K], добавлен 16.06.2014
Описание комплементарного взаимодействия генов. Рассмотрение характерных особенностей модификационной и наследственной (комбинативной, мутационной) закономерностей изменчивости организма. Задачи и методы селекции растений, животных и микроорганизмов.
реферат [20,8 K], добавлен 06.07.2010
Менделевская генетика. Гибридологический метод. Моногибридное и поли- схрещивание. Типы межаллельных взаимодействий. Наследование групп крови. Взаимодействие генов. Неменделевская генетика. Хромосомные аберрации. Наследование сцепленное с полом.
курсовая работа [1,9 M], добавлен 17.05.2004
Понятие "неаллельные гены". Исследование фенотипического проявления признаков при дигибридном скрещивании особей в случае различных форм взаимодействия неалельных генов – комплементарности, доминантного и рецессивного эпистаза, гипостаза и криптомерии.
Большинство признаков и свойств организмов характеризуются количественным типом индивидуальной изменчивости, для которой типично непрерывное изменение величины признака у особей какой-либо группы. Величина количественного признака варьирует от минимального уровня у части особей к среднему — у других и далее к максимальному уровню у остальных. Даже в пределах достаточно однородной по полу, возрасту, породе группы животных у близкородственных особей наблюдается индивидуальная изменчивость признака, величину которого можно измерить. К количественным признакам относят хозяйственно ценные (живая масса, величина, удой, настриг шерсти) и физиологические признаки. Они характеризуются типичным непрерывным изменением уровня у особей конкретной группы. К количественным признакам относят также и те, которые имеют прерывистое выражение, например яйценоскость, плодовитость, а также ряд физиологических отличий.
Количественные признаки непрерывного и прерывистого типов изменчивости имеют важное значение в практике животноводства и ветеринарии, в научных исследованиях, поэтому необходимо изучать генетические особенности и закономерности их изменчивости.
Генетические основы наследования количественных признаков. Наследование количественных признаков обусловлено одинаковым или сходным действием многих доминантных неаллельных генов на признак (полимерия) либо многими однозначными генами (полигения). На наличие двух или трех пар однозначно действующих полимерных генов, определяющих степень выраженности признака, указывает тип расщепления признака у особей второго поколения. Так, при трех доминантных генах А1; А2 и А3 и их рецессивных аллелях а1, а2, а3 во II поколении будут выявлены 64 варианта генотипов в соотношении 1 : 6: 15:20: 15:6: 1.
Если общая возможность развития признака связана с действием одного гена, то его принято называть главным (менделирующим) геном (олигогеном) и тогда признак наследуется в соответствии с законом Менделя. Полигены могут проявлять модифицирующее влияние на количественные признаки и составлять группу генов-модификаторов, то есть генов, которые, действуя каждый отдельно, проявляют слабое влияние на изменение в фенотипе, вызванное действием главного гена. Гены-модификаторы могут оказывать влияние и при отсутствии главного гена.
Полимерные гены способствуют увеличению изменчивости и формированию различных подгрупп (экотипов) внутри вида, так как они обеспечивают многообразные рекомбинации генотипов. Влияние рекомбинации и отбора в разных условиях среды способствует образованию несходных экотипов и повышению приспособленности вида к многообразию факторов внешней среды, формированию наследственной адаптации.
Гаметическая интеграция способствует формированию устойчивости разнообразных признаков и свойств, благоприятствующих повышению приспособленности, так как образуются адаптивные комплексы генов, которые повышают индивидуальную изменчивость организмов по степени их приспособленности к условиям среды. Гаметическая интеграция межлокусных корреляций обусловливает на фенотипическом уровне взаимодействие между локусэми неаллельных полигенов. Она может быть выявлена при помощи статистических параметров (σ, σ2, Cv).
Приспособленность организмов могут характеризовать величины среднего уровня жизнеспособности особей, их плодовитость, интенсивность развития всех особей данной группы (популяции).
Обобщение теории роли гаметической интеграции было сделано в трудах Майра, 1974; Левонтина, 1978; Животовского, 1984; и др.
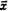
Методы изучения изменчивости и наследуемости количественных признаков. Фенотипическую изменчивость количественных признаков определяют с помощью статистических параметров (, σ, Cv), фенотипическую связь между признаками — применением r, b, rs и др. Для генетического анализа изменчивости количественных признаков требуется разложить фенотипическую изменчивость, выраженную через вариансу σр2, на составляющие вариансы: генотипическую (σg2) и паратипическую (σе2). Это позволяет установить в группе особей долю изменчивости, обусловленную их генетическим разнообразием, и долю изменчивости, связанную с влиянием факторов среды.
Возникает вопрос: существуют ли тупики отбора, когда его действие прекращается? Оказывается, что постоянный процесс мутирования создает новый источник изменчивости и тупика отбора не возникает.
Для выявления наследуемости признака широко применяют методы корреляционного и дисперсионного анализа. Тем самым выявляется доля генетического влияния на признак в фенотипической изменчивости.
Наследуемость. В генетических исследованиях необходимо различать три близких, но разных понятия: наследственность, наследование и наследуемость. Наследственность — это биологическое свойство, проявляющееся в сходстве родителей и потомков. Наследование — это способ передачи наследственности родителей потомкам с помощью гамет и их хромосомного аппарата.
Наследуемость — это статистический термин, который применяют для обозначения доли общей фенотипической изменчивости, обусловленной генетическими факторами. Наследуемость характеризует количественный признак у группы животных и служит показателем для прогнозирования эффективности селекции по фенотипическим показателям признака.
Величина коэффициента наследуемости (h2) служит мерой выражения генетической детерминации изменчивости признака. Величина Л2 не может быть больше единицы и меньше нуля (то есть отрицательной). Формула для определения h2 выражена следующим образом:
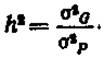
Чем больше величина h2, тем больше изменчивость признака обусловлена генетическими факторами и тем меньше изменчивость, вызываемая факторами среды (σЕ2). При h2 менее 0,05 (то есть менее 5%) улучшение признака за счет массовой селекции малоэффективно. При h2>0,3 и не менее 0,7 селекция достаточно эффективна.
Особенности факторов среды и ряд характеристик популяции (направление отбора в ней, наличие инбридинга, миграции особей, интенсивность браковки в стаде и т. п.) влияют на величину коэффициента наследуемости. Большая изменчивость факторов среды уменьшает h2, а однородность условий увеличивает или стабилизирует его величину.
Вычисленные коэффициенты наследуемости могут характеризовать только данную популяцию и в данных условиях среды и отбора.
При рассмотрений величины h2 для разных признаков выявлена следующая тенденция: признаки, которые связаны с репродукцией и приспособленностью к процессу размножения, характеризуются невысоким коэффициентом наследуемости (плодовитость у свиней: h2 = 5%; яйценоскость у кур — 10%), а признаки, имеющие меньшее значение для приспособленности, характеризуются более высоким уровнем h2 (масса животных — 55—65%; толщина шпига у свиней —70%, молочность и жирномолочность — 35—40 %).
Определение коэффициента наследуемости с использованием коэффициентов корреляции (r) и регрессии (b). Метод коэффициентов путей, предложенный С. Райтом, позволяет выявить величину связи между фенотипической изменчивостью (следствие) и влиянием генотипа (причина) или среды (причина). Этот метод нашел применение для определения величины связи между генотипами родственных особей.
Для определения доли влияния генотипа на изменчивость признака используют коэффициенты путей между родственными животными, например между матерями и дочерьми или между полусибсами, отцами и сыновьями и т. п. Графическое изображение коэффициентов путей представлено на рис. 61,

где направление влияния причин указано стрелками. Из схемы следует, что генотип дочерей определяется генотипом матерей, величина влияния которого (а) равна 0,5. Остальная доля наследственности дочерей обусловлена генотипом отцов. Вместе с тем генотипы матерей оказывают влияние на их фенотипы (на схеме это обозначено h). Соответственно генотипы дочерей определяют их фенотип. Связь между фенотипами матерей и Дочерей может быть определена с помощью обычного коэффициента корреляции rмд. Из схемы видно, что звенья rмд, h, a, h образуют замкнутую цепь. В ней известны два звена:
и rмд, который вычисляют по конкретным данным. Так как связь между звеньями выражается произведением коэффициентов путей, то можно записать равенство: 0,5·h·h=rмд, или 0,5 h2 =rмд , тогда h2 =2rмд. При использовании Коэффициента регрессии bмд формула примет следующий вид: h2=2bмд. Полученная формула удвоенного коэффициента корреляций (или регрессии) между фенотипическими показателями матерей и их дочерей выражает величину коэффициента наследуемости h2, который определяет долю генетического влияния на фенотипическую изменчивость признака,
Если полученный коэффициент наследуемости — величина отрицательная или больше единицы, то причину этого можно объяснить следующим образом:
- группы животных, подвергнутых дисперсионному анализу, малочисленны, то есть объем выборки недостаточен;
- неоднозначное реагирование на изменений факторов среды материнских и дочерних особей и смена рангов тех и других. Нет смысла определять величину h2, если имеют место резкие различия в условиях между поколениями родителей и потомков:
- проявление межаллельного взаимодействий генов (доминирование, сверхдоминирование), влияние инбридинга.
Возможно также определение h2, основанное на сопоставлении признаков полусибсов, то есть у потомства от одного из родителей, по формуле h2=4rп/С или (реже) у полных сибсов: h2=2rc.
При сопоставлении результатов использования указанных формул выясняется, что они дают несколько различные величины h2 для одного и того же выборочного материала.
Для вычисления h2 можно применять дисперсионный анализ с разными типами статистических комплексов, используя формулу h2 =η2=Сx:Су (по Н. А. Плохинскому) или формулу Д. Снедекора: h2= σx2: (σx2+ σz2).
Коэффициент повторяемости. При проведении генетического анализа количественных признаков выявляют популяционный показатель, тесно связанный с h2 и имеющий важное значение для оценки наследственности. Этим показателем является коэффициент повторяемости (rw), Его свойства заключаются в следующем: a) rw — показатель генетического разнообразия; б) он является мерой верхнего предела коэффициента наследуемости; в) определяет надежность вносимых поправок в варьирующий признак с учетом изменения средовых факторов; г) служит мерой определения ошибки измеряемого признака.
При высокой величине повторяемости уровня признака особи сохраняют определенный его уровень, что дает возможность более эффективно вести селекцию.
Для вычисления rw чаще всего используют однофакторный дисперсионный комплекс. Коэффициент повторяемости находится в границах от 0 до +1. Для целей селекции предпочтительнее, если rw имеет большую величину, Считают, что если rw 8
В своем реферате я рассмотрю такие вопросы, как законы наследования, генную инженерию и биотехнологии.
Генетика является одной из самых прогрессивных наук естествознания. Ее достижения изменили естественнонаучное и во многом философское понимание явлений жизни. Роль генетики для практики селекции и медицины очень велика. Значение генетики для медицины будет возрастать с каждым годом, ибо генетика касается самых сокровенных сторон биологии и физиологии человека. Благодаря генетике, ее знаниям, разрабатываются методы лечения ряда наследственных заболеваний, таких, как фенилкетонурия, сахарный диабет и другие. Здесь медико-генетическая работа призвана облегчить страдания людей от действия дефектных генов, полученных ими от родителей. Внедряются в практику приемы медико-генетического консультирования и прентальной диагностики, что позволяет предупредить развитие наследственных заболеваний.
1. ГЕНЕТИКА ПОЛА
Пол — совокупность признаков, по которым производится специфическое разделение особей или клеток, основанное на морфологических и физиологических особенностях, позволяющее осуществлять в процессе полового размножения комбинирование в потомках наследственных задатков родителей.
Морфологические и физиологические признаки, по которым производится специфическое разделение особей, называется половым.
Признаки, связанные с формированием и функционированием половых клеток, называется первичными половыми признаками. Это гонады (яичники или семенники), их выводные протоки, добавочные железы полового аппарата, копулятивные органы. Все другие признаки, по которым один пол отличается од другого, получили название вторичных половых признаков. К ним относят: характер волосяного покрова, наличие и развитие молочных желез, строение скелета, тип развития подкожной жировой клетчатки, строение трубчатых костей и др.
1.1. Генетические механизмы формирования пола
Начало изучению генотипического определения пола было положено открытием американскими цитологами у насекомых различия в форме, а иногда и в числе хромосом у особей разного пола (Мак-Кланг, 1906, Уилсон, 1906) и классическими опытами немецкого генетика Корренса по скрещиванию однодомного и двудомного видов брионии. Уилсон обнаружил, что у клопа Lydaeus turucus самки имеют 7 пар хромосом, у самцов же 6 пар одинаковых с самкой хромосом, а в седьмой паре одна хромосома такая же, как соответствующая хромосома самки, а другая маленькая.
Пара хромосом, которые у самца и самки разные, получила название идио, или гетерохромосомы, или половые хромосомы. У самки две одинаковые половые хромосомы, обозначаемые как Х-хромосомы, у самца одна Х-хромосома, другая — Y-хромосома. Остальные хромосомы одинаковые у самца и у самки, были названы аутосомами. Таким образом, хромосомная формула у самки названного клопа запишется 12A + XX, у самца 2A + XY. У ряда других организмов, хотя и существует в принципе тот же аппарат для определения пола, однако гетерозиготны в отношении реализаторов пола не мужские, а женские организмы. Особи мужского пола имеют две одинаковые половые хромосомы ZZ, а особи женского пола — ZO или ZW. ZZ-ZW тип определения пола наблюдается у бабочек, птиц, ZZ-ZO — ящериц, некоторых птиц.
Совершенно другой механизм определения пола, называемый гаплодиплоидный, широко распространен у пчел и муравьев. У этих организмов нет половых хромосом: самки — это диплоидные особи, а самцы (трутни) — гаплоидные. Самки развиваются из оплодотворенных яиц, а из неоплодотворенных развиваются трутни.
Человек в отношении определения пола относится к типу XX-XY. При гаметогенезе наблюдается типичное менделевское расщепление по половым хромосомам. каждая яйцеклетка содержит одну Х-хромосому, а другая половина — одну Y-хромосому. Пол потомка зависит от того, какой спермий оплодотворит яйцеклетку. Пол с генотипом ХХ называют гомогаметным, так как у него образуются одинаковые гаметы, содержащие только Х-хромосомы, а пол с генотипом XY-гетерогаметным, так как половина гамет содержит Х-, а половина — Y-хромосому. У человека генотипический пол данного индивидума определяют, изучая неделящиеся клетки. Одна Х-хромосома всегда оказывается в активном состоянии и имеет обычный вид. Другая, если она имеется, бывает в покоящемся состоянии в виде плотного темно-окрашенного тельца, называемого тельцем Барра (факультативный гетерохроматин). Число телец Барра всегда на единицу меньше числа наличных х-хромосом, т.е. в мужском организме их нет вовсе, у женщин (ХХ) — одно. У человека Y-хромосома является генетически инертной, так как в ней очень мало генов. Однако влияние Y-хромосомы на детерминацию пола у человека очень сильное. Хромосомная структура мужчины 44A+XY и женщины 44A+XX такая же, как и у дрозофины, однако у человека особь кариотипом 44A+XD оказалась женщиной, а особь 44A+XXY мужчиной. В обоих случаях они проявляли дефекты развития, но все же пол определялся наличием или отсутствием y-хромосомы. Люди генотипа XXX2A представляют собой бесплодную женщину, с генотипом XXXY2A — бесплодных умственно отстающих мужчин. Такие генотипы возникают в результате нерасхождения половых хромосом, что приводит к нарушению развития (например, синдром Клайнфельтера (XXY). Нерасхождение хромосом изучаются как в мейозе, так и в нитозе. Нерасхождение может быть следствием физического сцепления Х-хромосом, в таком случае нерасхождение имеет место в 100% случаев.
Всем млекопитающим мужского пола, включая человека, свойственен так называемый H-Y антиген, находящийся на поверхности клеток, несущих Y-хромосому. Единственной функцией его считается дифференцировка гонад. Вторичные половые признаки развиваются под влиянием стероидных гормонов, вырабатываемых гонадами. Развитие мужских вторичных половых признаков контролирует тестостерон, воздействующий на все клетки организма, включая клетки гонад. Мутация всего одного Х-хромосомы, кодирующего белок-рецептор тестостерона, приводит к синдрому тестикумерной фелинизации особей XY. Клетки-мутанты не чувствительны в действию тестостерона, в результате чего взрослый организм приобретает черты, характерные для женского пола. При этом внутренние половые органы оказываются недоразвитыми и такие особи полностью стерильные. Таким образом, в определении и дифференцировке пола млекопитающих и человека взаимодействуют хромосомный и генный механизмы.
Несмотря на то, что женщины имеют две Х-хромосомы, а мужчины — только одну, экспрессия генов Х-хромосомы происходит на одном и том же уровне у обоих полов. Это объясняется тем, что у женщин в каждой клетке полностью инактивирована одна Х-хромосома (тельце Барра), о чем уже было сказано выше. Х-хромосома инактивируется на ранней стадии эмбрионального развития, соответствующей времени имплантации. при этом в разных клетках отцовская и материнская Х-хромосомы выключаются случайно. Состояние инактивации данной Х-хромосомы наследуется в ряду клеточных делений. Таким образом, женские особи, гетерозиготные по генам половых хромосом, представляют собой мозаики (пример, черепаховые кошки).
Таким образом, пол человека представляет собой менделирующий признак, наследуемый по принципу обратного (анализирующего) скрещивания. Гетерозиготой оказывается гетерогаметный пол (XY), который скрещивается с рецессивной гомозиготой, представленной гомогаметным полом (XX). В результате в природе обнаруживается наследственная дифференцировка организмов на мужской и женский пол и устойчивое сокращение во всех поколениях количественного равенства полов.
1.2. Наследование признаков, сцепленных с полом
Морган и его сотрудники заметили, что наследование окраски глаз у дрозофилы зависит от пола родительских особей, несущих альтернативные аллели. Красная окраска глаз доминирует над белой. При скрещивании красноглазого самца с белоглазой самкой в F1, получали равное число красноглазых самок и белоглазых самцов. Однако при скрещивании белоглазого самца с красноглазой самкой в F1 были получены в равном числе красноглазые самцы и самки. При скрещивании этих мух F1, между собой были получены красноглазые самки, красноглазые и белоглазые самцы, но не было ни одной белоглазой самки. Тот факт, что у самцов частота проявления рецессивного признака была выше, чем у самок, наводил на мысль, что рецессивный аллель, определяющий белоглазость, находится в Х — хромосоме, а Y — хромосома лишена гена окраски глаз. Чтобы проверить эту гипотезу, Морган скрестил исходного белоглазого самца с красноглазой самкой из F1. В потомстве были получены красноглазые и белоглазые самцы и самки. Из этого Морган справедливо заключил, что только Х — хромосома несет ген окраски глаз. В Y — хромосоме соответствующего локуса вообще нет. Это явление известно под названием наследования, сцепленного с полом.
Гены, находящиеся в половых хромосомах, называют сцепленными с полом. В Х-хромосоме имеется участок, для которого в Y-хромосоме нет гомолога. Поэтому у особей мужского пола признаки, определяемые генами этого участка, проявляются даже в том случае, если они рецессивны. Эта особая форма сцепления позволяет объяснить наследование признаков, сцепленных с полом.
При локализации признаков как в аутосоме, так и в Х- b Y-хромосоме наблюдается полное сцепление с полом.
У человека около 60 генов наследуются в связи с Х-хромосомой, в том числе гемофелия, дальтонизм (цветовая слепота), мускульная дистрофия, потемнение эмали зубов, одна из форм агаммглобулинемии и другие. Наследование таких признаков отклоняется от закономерностей, установленных Г.Менделем. Х-хромосома закономерно переходит от одного пола к другому, при этом дочь наследует Х-хромосому отца, а сын Х-хромосому матери. Наследование, при котором сыновья наследуют признак матери, а дочери — признак отца получило, название крисс-кросс (или крест-накрест).
Известны нарушения цветового зрения, так называемая цветовая слепота. В основе появления этих дефектов зрения лежит действие ряда генов. Красно-зеленая слепота обычно называется дальтонизмом. Еще задолго до появления генетики в конце XVIII и в XIX в. было установлено, что цветовая слепота наследуется согласно вполне закономерным правилам. Так, если женщина, страдающая цветовой слепотой, выходит замуж за мужчину с нормальным зрением, то у их детей наблюдается очень своеобразная картина перекрестного наследования. Все дочери от такого брака получат признак отца, т.е. они имеют нормальное зрение, а все сыновья, получая признак матери, страдают цветовой слепотой (а-дальтонизм, сцепленный с Х-хромосомой) .
В том же случае, когда наоборот, отец является дальтоником, а мать имеет нормальное зрение, все дети оказываются нормальными. В отдельных браках, где мать и отец обладают нормальным зрением, половина сыновей может оказаться пораженными цветовой слепотой. В основном наличие цветовой слепоты чаще встречается у мужчин. Э.Вильсон объяснил наследование этого признака, предположив, что он локализовал в Х-хромосоме и что у человека гетерогаметным (XY) является мужской пол. Становится вполне понятным, что в браке гомозиготной нормальной женщины (Х а Х а ) с мужчиной дальтоником (Х а y) все дети рождаются нормальными. Однако при этом, все дочери становятся скрытыми носителями дальтонизма, что может проявиться в последующих поколениях.
Другим примером наследования сцепленного с полом, может послужить рецессивныйполулетальный ген, вызывающий несвертываемость крови на воздухе — гемофилию. Это заболевание появляется почти исключительно только у мальчиков. При гемофилии нарушается образование фактора VIII, ускоряющего свертывание крови. ген, детерминирующий синтех фактора VIII, находится в участке Х-хромосомы, недоминантным нормальным и рецессивным мутантным. Возможны следующие генотипы и фенотипы:
В своем реферате я рассмотрю такие вопросы, как законы наследования, генную инженерию и биотехнологии.
Генетика является одной из самых прогрессивных наук естествознания. Ее достижения изменили естественнонаучное и во многом философское понимание явлений жизни. Роль генетики для практики селекции и медицины очень велика. Значение генетики для медицины будет возрастать с каждым годом, ибо генетика касается самых сокровенных сторон биологии и физиологии человека. Благодаря генетике, ее знаниям, разрабатываются методы лечения ряда наследственных заболеваний, таких, как фенилкетонурия, сахарный диабет и другие. Здесь медико-генетическая работа призвана облегчить страдания людей от действия дефектных генов, полученных ими от родителей. Внедряются в практику приемы медико-генетического консультирования и прентальной диагностики, что позволяет предупредить развитие наследственных заболеваний.
1. ГЕНЕТИКА ПОЛА
Пол — совокупность признаков, по которым производится специфическое разделение особей или клеток, основанное на морфологических и физиологических особенностях, позволяющее осуществлять в процессе полового размножения комбинирование в потомках наследственных задатков родителей.
Морфологические и физиологические признаки, по которым производится специфическое разделение особей, называется половым.
Признаки, связанные с формированием и функционированием половых клеток, называется первичными половыми признаками. Это гонады (яичники или семенники), их выводные протоки, добавочные железы полового аппарата, копулятивные органы. Все другие признаки, по которым один пол отличается од другого, получили название вторичных половых признаков. К ним относят: характер волосяного покрова, наличие и развитие молочных желез, строение скелета, тип развития подкожной жировой клетчатки, строение трубчатых костей и др.
1.1. Генетические механизмы формирования пола
Начало изучению генотипического определения пола было положено открытием американскими цитологами у насекомых различия в форме, а иногда и в числе хромосом у особей разного пола (Мак-Кланг, 1906, Уилсон, 1906) и классическими опытами немецкого генетика Корренса по скрещиванию однодомного и двудомного видов брионии. Уилсон обнаружил, что у клопа Lydaeus turucus самки имеют 7 пар хромосом, у самцов же 6 пар одинаковых с самкой хромосом, а в седьмой паре одна хромосома такая же, как соответствующая хромосома самки, а другая маленькая.
Пара хромосом, которые у самца и самки разные, получила название идио, или гетерохромосомы, или половые хромосомы. У самки две одинаковые половые хромосомы, обозначаемые как Х-хромосомы, у самца одна Х-хромосома, другая — Y-хромосома. Остальные хромосомы одинаковые у самца и у самки, были названы аутосомами. Таким образом, хромосомная формула у самки названного клопа запишется 12A + XX, у самца 2A + XY. У ряда других организмов, хотя и существует в принципе тот же аппарат для определения пола, однако гетерозиготны в отношении реализаторов пола не мужские, а женские организмы. Особи мужского пола имеют две одинаковые половые хромосомы ZZ, а особи женского пола — ZO или ZW. ZZ-ZW тип определения пола наблюдается у бабочек, птиц, ZZ-ZO — ящериц, некоторых птиц.
Совершенно другой механизм определения пола, называемый гаплодиплоидный, широко распространен у пчел и муравьев. У этих организмов нет половых хромосом: самки — это диплоидные особи, а самцы (трутни) — гаплоидные. Самки развиваются из оплодотворенных яиц, а из неоплодотворенных развиваются трутни.
Человек в отношении определения пола относится к типу XX-XY. При гаметогенезе наблюдается типичное менделевское расщепление по половым хромосомам. каждая яйцеклетка содержит одну Х-хромосому, а другая половина — одну Y-хромосому. Пол потомка зависит от того, какой спермий оплодотворит яйцеклетку. Пол с генотипом ХХ называют гомогаметным, так как у него образуются одинаковые гаметы, содержащие только Х-хромосомы, а пол с генотипом XY-гетерогаметным, так как половина гамет содержит Х-, а половина — Y-хромосому. У человека генотипический пол данного индивидума определяют, изучая неделящиеся клетки. Одна Х-хромосома всегда оказывается в активном состоянии и имеет обычный вид. Другая, если она имеется, бывает в покоящемся состоянии в виде плотного темно-окрашенного тельца, называемого тельцем Барра (факультативный гетерохроматин). Число телец Барра всегда на единицу меньше числа наличных х-хромосом, т.е. в мужском организме их нет вовсе, у женщин (ХХ) — одно. У человека Y-хромосома является генетически инертной, так как в ней очень мало генов. Однако влияние Y-хромосомы на детерминацию пола у человека очень сильное. Хромосомная структура мужчины 44A+XY и женщины 44A+XX такая же, как и у дрозофины, однако у человека особь кариотипом 44A+XD оказалась женщиной, а особь 44A+XXY мужчиной. В обоих случаях они проявляли дефекты развития, но все же пол определялся наличием или отсутствием y-хромосомы. Люди генотипа XXX2A представляют собой бесплодную женщину, с генотипом XXXY2A — бесплодных умственно отстающих мужчин. Такие генотипы возникают в результате нерасхождения половых хромосом, что приводит к нарушению развития (например, синдром Клайнфельтера (XXY). Нерасхождение хромосом изучаются как в мейозе, так и в нитозе. Нерасхождение может быть следствием физического сцепления Х-хромосом, в таком случае нерасхождение имеет место в 100% случаев.
Всем млекопитающим мужского пола, включая человека, свойственен так называемый H-Y антиген, находящийся на поверхности клеток, несущих Y-хромосому. Единственной функцией его считается дифференцировка гонад. Вторичные половые признаки развиваются под влиянием стероидных гормонов, вырабатываемых гонадами. Развитие мужских вторичных половых признаков контролирует тестостерон, воздействующий на все клетки организма, включая клетки гонад. Мутация всего одного Х-хромосомы, кодирующего белок-рецептор тестостерона, приводит к синдрому тестикумерной фелинизации особей XY. Клетки-мутанты не чувствительны в действию тестостерона, в результате чего взрослый организм приобретает черты, характерные для женского пола. При этом внутренние половые органы оказываются недоразвитыми и такие особи полностью стерильные. Таким образом, в определении и дифференцировке пола млекопитающих и человека взаимодействуют хромосомный и генный механизмы.
Несмотря на то, что женщины имеют две Х-хромосомы, а мужчины — только одну, экспрессия генов Х-хромосомы происходит на одном и том же уровне у обоих полов. Это объясняется тем, что у женщин в каждой клетке полностью инактивирована одна Х-хромосома (тельце Барра), о чем уже было сказано выше. Х-хромосома инактивируется на ранней стадии эмбрионального развития, соответствующей времени имплантации. при этом в разных клетках отцовская и материнская Х-хромосомы выключаются случайно. Состояние инактивации данной Х-хромосомы наследуется в ряду клеточных делений. Таким образом, женские особи, гетерозиготные по генам половых хромосом, представляют собой мозаики (пример, черепаховые кошки).
Таким образом, пол человека представляет собой менделирующий признак, наследуемый по принципу обратного (анализирующего) скрещивания. Гетерозиготой оказывается гетерогаметный пол (XY), который скрещивается с рецессивной гомозиготой, представленной гомогаметным полом (XX). В результате в природе обнаруживается наследственная дифференцировка организмов на мужской и женский пол и устойчивое сокращение во всех поколениях количественного равенства полов.
1.2. Наследование признаков, сцепленных с полом
Морган и его сотрудники заметили, что наследование окраски глаз у дрозофилы зависит от пола родительских особей, несущих альтернативные аллели. Красная окраска глаз доминирует над белой. При скрещивании красноглазого самца с белоглазой самкой в F1, получали равное число красноглазых самок и белоглазых самцов. Однако при скрещивании белоглазого самца с красноглазой самкой в F1 были получены в равном числе красноглазые самцы и самки. При скрещивании этих мух F1, между собой были получены красноглазые самки, красноглазые и белоглазые самцы, но не было ни одной белоглазой самки. Тот факт, что у самцов частота проявления рецессивного признака была выше, чем у самок, наводил на мысль, что рецессивный аллель, определяющий белоглазость, находится в Х — хромосоме, а Y — хромосома лишена гена окраски глаз. Чтобы проверить эту гипотезу, Морган скрестил исходного белоглазого самца с красноглазой самкой из F1. В потомстве были получены красноглазые и белоглазые самцы и самки. Из этого Морган справедливо заключил, что только Х — хромосома несет ген окраски глаз. В Y — хромосоме соответствующего локуса вообще нет. Это явление известно под названием наследования, сцепленного с полом.
Гены, находящиеся в половых хромосомах, называют сцепленными с полом. В Х-хромосоме имеется участок, для которого в Y-хромосоме нет гомолога. Поэтому у особей мужского пола признаки, определяемые генами этого участка, проявляются даже в том случае, если они рецессивны. Эта особая форма сцепления позволяет объяснить наследование признаков, сцепленных с полом.
При локализации признаков как в аутосоме, так и в Х- b Y-хромосоме наблюдается полное сцепление с полом.
У человека около 60 генов наследуются в связи с Х-хромосомой, в том числе гемофелия, дальтонизм (цветовая слепота), мускульная дистрофия, потемнение эмали зубов, одна из форм агаммглобулинемии и другие. Наследование таких признаков отклоняется от закономерностей, установленных Г.Менделем. Х-хромосома закономерно переходит от одного пола к другому, при этом дочь наследует Х-хромосому отца, а сын Х-хромосому матери. Наследование, при котором сыновья наследуют признак матери, а дочери — признак отца получило, название крисс-кросс (или крест-накрест).
Известны нарушения цветового зрения, так называемая цветовая слепота. В основе появления этих дефектов зрения лежит действие ряда генов. Красно-зеленая слепота обычно называется дальтонизмом. Еще задолго до появления генетики в конце XVIII и в XIX в. было установлено, что цветовая слепота наследуется согласно вполне закономерным правилам. Так, если женщина, страдающая цветовой слепотой, выходит замуж за мужчину с нормальным зрением, то у их детей наблюдается очень своеобразная картина перекрестного наследования. Все дочери от такого брака получат признак отца, т.е. они имеют нормальное зрение, а все сыновья, получая признак матери, страдают цветовой слепотой (а-дальтонизм, сцепленный с Х-хромосомой) .
В том же случае, когда наоборот, отец является дальтоником, а мать имеет нормальное зрение, все дети оказываются нормальными. В отдельных браках, где мать и отец обладают нормальным зрением, половина сыновей может оказаться пораженными цветовой слепотой. В основном наличие цветовой слепоты чаще встречается у мужчин. Э.Вильсон объяснил наследование этого признака, предположив, что он локализовал в Х-хромосоме и что у человека гетерогаметным (XY) является мужской пол. Становится вполне понятным, что в браке гомозиготной нормальной женщины (Х а Х а ) с мужчиной дальтоником (Х а y) все дети рождаются нормальными. Однако при этом, все дочери становятся скрытыми носителями дальтонизма, что может проявиться в последующих поколениях.
Другим примером наследования сцепленного с полом, может послужить рецессивныйполулетальный ген, вызывающий несвертываемость крови на воздухе — гемофилию. Это заболевание появляется почти исключительно только у мальчиков. При гемофилии нарушается образование фактора VIII, ускоряющего свертывание крови. ген, детерминирующий синтех фактора VIII, находится в участке Х-хромосомы, недоминантным нормальным и рецессивным мутантным. Возможны следующие генотипы и фенотипы:
Читайте также: