
Жгутики кратко и понятно
Обновлено: 05.07.2024
Жгутик (лат. Flagellum) — поверхностная и внеклеточная структура, присутствует во многих прокариот и эукариот, что служит для передвижения в жидкой среде или поверхностью влажных твердых сред. Жгутики прокариот и эукариот резко отличаются по своему строению, например, бактерильний жгутик имеет толщину 10-20 нм и длину 3-15 мкм, он пассивно вращается расположенным в мембране мотором; тогда как жгутики эукариот имеют толщину до 200 нм и длину до 200 мкм, они могут самостоятельно изгибаться по всей длине. У эукариот часто также присутствуют реснички, идентичные по своему строению жгутика, но короче (до 10 мкм), вместе они называются ундулиподиямы.
Жгутики бактерий
Жгутики бактерий состоят из трех субструктур:
- Филамент (фибриллами, пропеллер) — полый белковое волокно толщиной 10-20 нм и длиной 3-15 мкм, состоящий из белков FlaA (флагеллина) и FlaB, субъединицы FlaA заключены по спирали. Полость внутри используется при синтезе жгутика — он происходит в направлении от цитоплазматической мембраны. Полостью к участку, что собирается на данный момент, переносятся субъединицы флагеллина.
- Крючок — образование, толще, чем филамент (20-45 нм), составленное из белка FlgE (возможно, также других белков).
- Базальное мотор (базальное тело).
Базальное мотор и механизм его работы
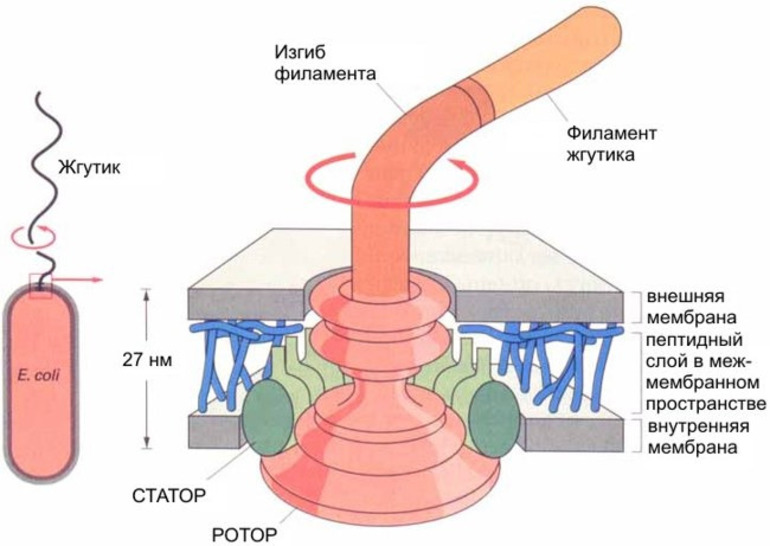
Базальное мотор представляет собой систему колец, находящихся в цитоплазматической и внешний мембранах и в клеточной стенке бактерии. Два внутренних кольца — M (белок FliF) и S (белки FliG, FliM, FliN) (также рассматриваются как единое MS-кольцо) — являются обязательными элементами, причем M-кольцо находится в цитоплазматической мембране, а S — в периплазматическое грамм -негативных и пептидогликановый слое грамположительных бактерий. Еще два кольца — P и L (белки FlgH, FlgI) — присутствуют только в грамм-отрицательных бактерий, они расположены в пептидогликановый слое и наружной мембране соответственно, неподвижные и только направляют стержень ротора двигателя. Вокруг MS-кольца расположены статоры — белковые комплексы MotA 4 / MotB 4 (в других видах вместо них могут быть белки PomA, PomB, MotX и MotY), каждый из этих комплексов имеет протонный канал (их может быть от 8 до 16).
Механизм работы жгутикового мотора очень сильно напоминает механизм работы трансмембранной части АТФ-синтазы или F 1 F 0, комплекса, синтезирует АТФ и присутствует во всех живых клетках. Существует теория, что эволюционно Базальное мотор происходит именно от АТФ-синтазы, скомбинувалася в процессе эволюции с системой секреции белков 3 типа. Особенностью этого мотора является возможность (в большинстве видов) работы в любом направлении и быстрое переключение между направлениями работы.
Энергия для работы мотора приобретается за счет электрического потенциала через цитоплазматическую мембрану. Мотор пропускает протоны с периплазматическое (или внешней среды) в цитоплазму. Некоторые бактерии используют ионы натрия вместо протонов (некоторые морские бактерии рода Vibrio, алкалофильного Bacillus, Acetobacterium woodii). Эти ионы должны пройти через канал, расположенный частично в статоре (точнее, белка FliG), а частично в роторе мотора (MotA, MotB). Так как часть Калалы параллельная мембране и направления вращения, электрический потенциал, толкает ион вдоль направления электрического поля, вращает ротор относительно статора. Например, в Escherichia coli для одного оборота жгутика требуется перемещение около 1000 протонов. Показано, что жгутик может работать даже в пустых клеточных оболочках при наличии электрического потенциала на мембране. Жгутик может вращаться с скорость и до 100 об / сек, при этом направление вращения может изменяться менее чем за 0,1 сек.
Механизм движения клетки
Вращения мотора вызывает пассивное вращения филамента. Массивная клетка начинает вращаться примерно со скоростью 1/3 от скорости жгутика в обратном направлении, а также приобретает поступательного движения. Подавляющее большинство наделенных жгутиком бактерий имеют палочковидную форму. С гидродинамических расчетов получается, что для эффективного движения отношение длины клетки к ширине должно составлять 3,7. Движение кокков крайне неэффективен, поэтому они чаще неподвижны.
Некоторые бактерии, например, спирохеты, имеют жгутики, расположенные в периплазматическое бактерий (так называемые продольные филаменты), движение жгутиков заставляет всю бактерию менять свою форму, за счет чего она и движется.
Скорости движения бактерий варьируют от 20 мкм / с в некоторых Bacillus до 200 мкм / с в Vibrio.
Регуляция движения
У ряда бактерий мотор и жгутик могут вращаться только в одном направлении, переориентация направления движения бактерии происходит при остановке за счет броуновского движения.
При полярном расположении жгутиков один из них может толкать бактерию, а другой тянуть и наоборот, в зависимости от направления вращения.
Жгутики архей
Жгутики архей относительно достаточно подобные бактериальных жгутиков. В 1980-х годах они даже считались гомологичными на основании подобной морфологии и поведения (Cavalier-Smith, 1987). Как и у бактерий, жгутики архей состоят из филаментов, которые тянутся за пределами клетки и вращаются, двигая клетку. Однако исследования 1990-х годов выявили многочисленные детальные различия между жгутиками архей и бактерий, например:
- Бактериальные жгутики работают за счет пропускания в цитоплазму протонов H + (иногда ионов натрия Na +), архейни жгутики почти наверняка тратят энергию АТФ. Хотя мотор архей все еще не исследован.
- В то время, когда бактериальные клетки часто имеют много жгутиков, каждый из которых вращается независимо архейни жгутики состоят из пучка многих филаментов, которые вращаются как единое целое.
- Бактериальные жгутики растут пополнением субъединиц флагеллина на наконечнике, архейни жгутики растут достройкой судодиниць к основанию.
- Бактериальные жгутики толще от архейних, и бактериальный филаминт имеет достаточно широкий простор внутри, через который субъединицы поставляются для роста жгутика, архейни жгутики слишком тонкие, чтобы позволить это.
- Гены многих компонентов бактериальных жгутиков имеют некоторое сходство последовательности ДНК к компонентам системы секреции 3 типа, но компоненты бактериальных и архейних жгутиков не разделяют никакого сходства в последовательности. В свою очередь, некоторые компоненты архейних жгутиков разделяют последовательность и морфологическое сходство с компонентами ворсинок 4 типа, которые собираются с помощью системы секреции 2 типа (номенклатура ворсинок / пили и систем секреции белка не последовательная).
Эти различия означают, что бактериальные и архейни жгутики — классический случай конвергентной эволюции, а не гомологии. Однако, в сравнении с десятилетия детального изучения бактериальных жгутиков (например Говардом Бергом), архейни жгутики только недавно начали получать серьезную научную внимание. Поэтому многие все еще помолково считает, что существует только один основной вид жгутиков прокариот, и архейни жгутики принадлежат к нему.
Жгутики эукариот
Жгутики эукариот — совершенно другая структура, чем жгутики бактерий и архей, и имеют совсем другое эволюционное происхождение. Единственная общая черта между бактериальными, архейнимы и эукариотическими жгутиками — их внешний вид, то, что они все — внеклеточные структуры, используемые для движения. Вместе с ресничками они составляют группу органелл, известных как ундулиподии.
Подвижные жгутики служат для движения отдельных клеток (например, плавающих простейшие и сперматозоидов) и транспорта жидкостей (например, транспорт слизи стационарными клетками в трахеи). В дополнение к этому, неподвижные жгутики — жизненно важные органеллы в органах чувств и передачи сигналов в большом количестве типов клеток (например, фоторецепторные палочки глаза, обонятельные нейроне носа, киносилиум в улитке уха).
Внутришньоджгутиковий транспорт (ВДТ, IFT), процесс, при котором субъединицы аксонема, трансмембранные рецепторы и другие белки продвигаются увздовж жгутика, существенный для надлежащего функционирования жгутика, как движения, так и передачи сигналов.
Жгутик — поверхностная структура, присутствующая у многих прокариотических и эукариотических клеток и служащая для их движения в жидкой среде или по поверхности твёрдых сред. Жгутики прокариот и эукариот резко различаются: бактериальный жгутик имеет толщину 10-20 нм и длину 3-15 мкм, он пассивно вращается расположенным в мембране мотором; жгутики же эукариот толщиной до 200 нм и длиной до 200 мкм, они могут самостоятельно изгибаться по всей длине. У эукариот часто также присутствуют реснички, идентичные по своему строению жгутику, но более короткие (до 10 мкм).
Содержание
Жгутики прокариот

Жгутикование бактерий: A — монотрихиальное,
B — лофотрихиальное,
C — амфитрихиальное,
D — перитрихиальное.
Жгутики бактерий состоят из трёх субструктур:
- Филамент (фибрилла, пропеллер) — полая белковая нить толщиной 10-20 нм и длиной 3-15 мкм, состоящая из флагеллина, субъединицы которого уложены по спирали. Полость внутри используется при синтезе жгутика — он происходит в направлении от ЦПМ. По полости к собираемому в настоящий момент участку переносятся субъединицы флагеллина.
- Крюк — более толстое чем филамент (20-45 нм) белковое (не флагеллиновое) образование.
- Базальное тело (трансмембранный мотор)
Базальное тело и механизм его работы
Базальное тело представляет собой систему колец, находящихся в ЦПМ и клеточной стенке бактерий. Два внутренних кольца — M и S-кольца (сейчас чаще рассматриваются как единое MS-кольцо) — являются обязательными элементами, причём M-кольцо находится в ЦПМ, а S — в периплазме грамотрицательных и пептидогликановом слое грамположительных бактерий. Ещё два кольца — P и L — есть только у грамотрицательных бактерий, они расположены в пептидогликановом слое и наружной мембране соответственно, неподвижны и лишь направляют стержень ротора мотора. Вокруг MS-кольца расположены статоры — белковые комплексы MotA4/MotB4 представляющие собой протонный канал (их может быть от 8 до 16).
Точный механизм работы базального тела не известен. Большинство исследователей полагает что поступление протона из периплазмы или внешней среды в MotA4/MotB4 комплекс вызывает конформационные изменения белков, благодаря электростатическому взаимодействию или прямому контакту это изменение приводит к повороту MS-кольца, а его дальнейшее движение возвращает исходную конформацию комплексу и выталкивает протон в цитозоль. У Escherichia coli для одного оборота жгутика требуется перемещение около 1000 протонов. Показано, что жгутик может работать даже у пустых клеточных оболочек при условии что внешний pH ниже внутреннего.
Таким образом, базальное тело преобразует химическую энергию в работу, вращаясь за счёт градиента концентрации протонов или, в редких случаях, ионов натрия (некоторые морские бактерии рода Vibrio, алкалофильные Bacillus, Acetobacterium woodii), это вращение осуществляется со скоростью до 100 об/сек, причём его направление может изменяться менее чем за 0,1 сек.
Механизм движения клетки
Вращение мотора вызывает пассивное вращение филамента. Более массивная клетка начинает вращаться примерно со скоростью 1/3 от скорости жгутика и в обратном направлении, а также приобретает поступательное движение.
Подавляющее большинство наделённых жгутиком бактерий имеют палочковидную форму. Из гидродинамических расчётов следует, что для наиболее эффективного движения отношение длины клетки к ширине должно составлять 3,7. Движение кокков крайне неэффективно, поэтому они чаще всего неподвижны.
У ряда бактерий мотор и жгутик могут вращаться только в одном направлении, переориентация происходит при остановке за счёт броуновского движения. Бактерии-перитрихи собирают при движении все свои жгутики (каждый из которых вращается против часовой стрелки) в один пучок. Для совершения кувырка они либо меняют направление вращения, либо изменяют его скорость, из-за чего пучок распадается. При полярном расположении жгутиков один из них может служить и толкающим, и тянущим приспособлением в зависимости от направления вращения.
Скорости движения бактерий варьируют от 20 мкм/с у некоторых Bacillus до 200 мкм/с у Vibrio.
Особенности жгутиков архей
В результате секвенирования геномов архей не удалось выявить какой-либо гомологии генов, ответственных за биогенез жгутиков архей и бактерий. Вместо флагеллина, неустойчивого в среде с повышенной кислотностью, в жгутиках архебактерий этот белок заменён гликопротеинами. Архебактериальный жгутик тоньше и не имеет центрального полого канала, поэтому, по всей видимости, его синтез происходит по принципиально иному механизму у поверхности клетки.
Синтез жгутика
Процесс синтеза жгутика эубактерий (Caulobacter sp.) запускается экспрессией гена сtrA. Продуктом этого гена является белок CtrA. Синтез Ctr A происходит сразу после перехода клетки из G0-фазы в S-фазу. Обычно участок ДНК, содержащий ген сtrA, метилирован. Синтезу белка CtrA предшествует деметилирование ДНК, которая затем реплицируется. После этого происходит синтез CtrA и его фосфорилирование киназами. Ген сtrA имеет два промотора: Р1 и Р2. С первого промотора осуществляется ингибирование синтеза CtrA. Со второго промотора осуществляется стимулирование синтеза CtrA. Следует отметить, что белок CtrA найден не у всех эубактерий, и механизм синтеза жгутика не универсален.
Жгутики эукариот
Схема поперечного среза жгутика эукариот. 1A и 1B — A и B микротрубочки периферического дублета, 2 — центральная пара микротрубочек и центральная капсула, 3 — динеиновые ручки, 4 — радиальная спица, 5 — нексиновый мостик, 6 — клеточная мембрана.
Жгутики эукариот имеют толщину до 200 нм и длину до 200 мкм. Они окружены выступами цитоплазматической мембраны и содержат 9 пар микротрубочек, выстроенных вокруг двух не объединённых в пару микротрубочек (структура 9+2). Эти микротрубочки скользят друг относительно друга с использованием энергии АТФ, поэтому изгиб эукариотического жгутика может осуществляться в любой его части.
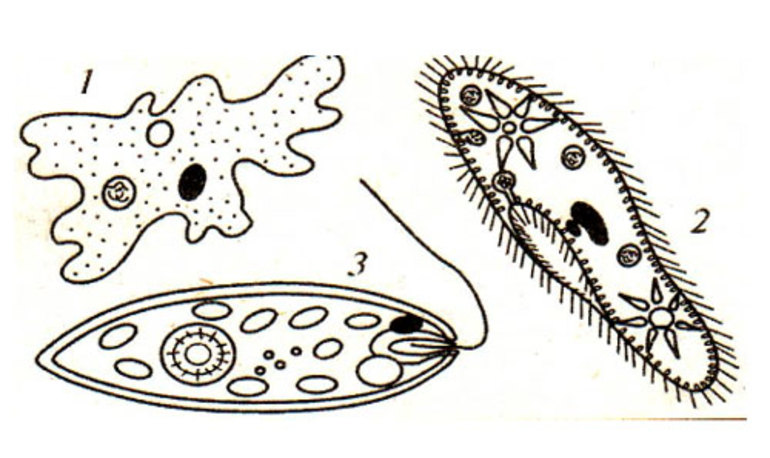
Органоиды движения — небольшие наросты на клеточной мембране, состоящие из системы микротрубочек. Они позволяют клеткам свободно перемещаться, что способствует росту живых организмов. Реснички, жгутики, псевдоподии и миофибриллы являются основными представителями органоидов движения. Строение и функции этих органелл изучаются на уроках биологии в 6 классе.
Краткая информация
Органоиды движения содержатся в растительных и животных клетках, входящих в состав многоклеточных организмов. Структура этих органелл формируется из молекул белков и фосфолипидов. Их средний размер составляет 0,25—100 мкм. В таблице перечислены основные особенности органоидов движения.
| Наименование органоида движения | Строение | Функции органоидов движения | Название одноклеточного организма |
| Жгутики | Цитоплазматические наросты, расположенные на поверхности мембраны | Передвижение клеток | Жгутиковые инфузории |
| Реснички | Тонкие выросты на эластичной структуре клетки | Очистка органов от пыли | Ресничные инфузории |
| Псевдоподии (ложноножки) | Выступы в цитоплазме клетки | Питание и передвижение организма | Саркодовые |
| Миофибриллы | Нити малой толщины | Сокращение мышц | Корненожки |
В человеческом организме присутствует большое количество ресничек и жгутиков. Они предназначены для очищения легких, защиты эпителия и стабильного функционирования репродуктивной системы. Принцип работы этих органоидов движения заключается в установлении прочных связей с клеточной мембраной.
Жгутики бактерий и архей
Жгутик — органоид движения эукариотов, обеспечивающий передвижение клеточных организмов в жидкой среде. Они содержатся в протистах, зооспорах и половых клетках. Эти органеллы представляют собой небольшие наросты, окруженные эластичной пленкой. Жгутики имеют цитоскелет, где осуществляется процесс гидролиза АТФ. Второстепенные функции жгутиков:
- формирование биологических пленок;
- обеспечение контакта клеточных организмов с субстратами;
- облегчение проникания симбиотических бактерий в клетки;
- включение защитных механизмов иммунной системы;
- предотвращение заражения клетки инфекционными вирусами.
Жгутик эукариотических клеток представляет собой комплексный структурный элемент. Он включает в себя 9 пар микротрубочек, соединенных нексиновыми мостиками. Между ними присутствует переходная зона эксонемы. В центральной части жгутика располагается ось с центриолями. На следующем рисунке описано строение органоида в разрезе.
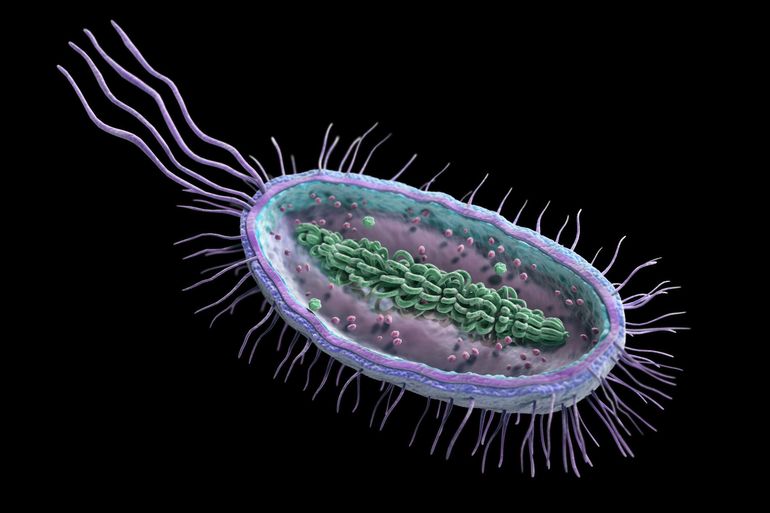
80% бактерий состоят из жгутиков. Они находятся на противоположных полюсах клеточного организма. Отличительной чертой жгутиков бактерий является их расположение в клетке. Они вмонтированы в оболочку клеточного организма. Вращение жгутика осуществляется при помощи энергии, получаемой при гидролизе АТФ. Органоид перемещается по часовой стрелке. Частотный диапазон вращения органеллы составляет от 200 до 1850 Гц. Бактериальный жгутик состоит из следующих компонентов:
- Филамент. Представляет собой нитевидную структуру. Длина этого компонента составляет не более 14 мкм. Нить располагается за пределами цитоплазмы.
- Базальное тело. Представлено в виде муреинового чехла, окруженного мембранной оболочкой. Оно состоит из системы секреции и мотора.
- Крюк. Гибкий элемент, соединенный с филаментом и базальным телом. Его длина составляет 55 нм.
Основным рабочим элементом жгутика бактерий является филамент. Этот компонент объединяет несколько тысяч субъединиц фосфолипидов и белка. При вращении органоида филамент приобретает форму спирали, закрученной в левую сторону.
Базальное тело бактериального жгутика состоит из следующих частей:
- оси клеточного центра;
- L-кольца;
- P-кольца;
- MS-кольца;
- ротора;
- C-кольца.
Для определения характера передвижения жгутика требуется знать количество ресурсов, поставляемых основными компонентами базального тела.
Жгутики архей состоят из археллума, включающего в себя 7—13 различных генов. Структура этого органоида формируется едиными оперонами. Функцией жгутиков архей является перемещение клеточных организмов во влажной среде, но они не принимают участия в формировании биологических пленок.
Механизм работы ресничек
Реснички представляют собой тонкие органеллы в форме волоса. Находясь в неподвижном состоянии, эти органоиды выполняют роль рецепторов. Поверхность ресничек покрыта плотной цитоплазматической мембраной. В состав органеллы входит большое количество микротрубочек. В центральной части ресничек расположено базальтовое тело с центриолями.
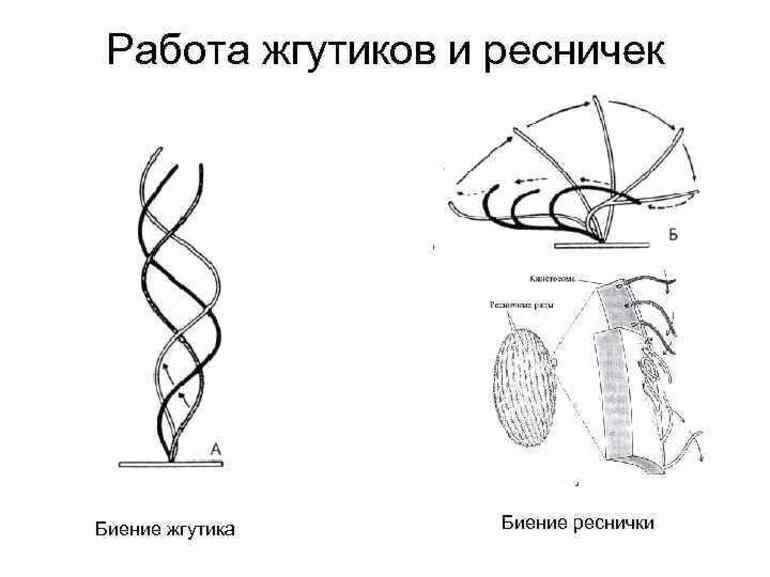
В микротрубочках присутствуют белковые структуры, обеспечивающие стабильное скольжение ресничек. Во время передвижения органоиды совершают удары. Они предназначены для деполяризации клеточной мембраны. Характер и направление ударов зависят от процентного содержания ионов кальция в структуре ресничек.
Псевдоподии и миофибриллы
Псевдоподии представляют собой цитоплазматические выросты без плотной клеточной оболочки. В школьных учебниках эти органеллы также могут называться ложноножками. Псевдоподии сдержат простейшие организмы:
- амебы;
- арцеллы;
- фораминиферы.
Ложноножки предназначены для всасывания питательных веществ. Также к их функциям относится перемещение клеточных организмов. Скорость движения клеток составляет не более 0,2 мм/мин. Во время перемещения псевдоподии закрепляются в субстрате и захватывают частицы пищи. В результате этого процесса формируется пищеварительная вакуоль.
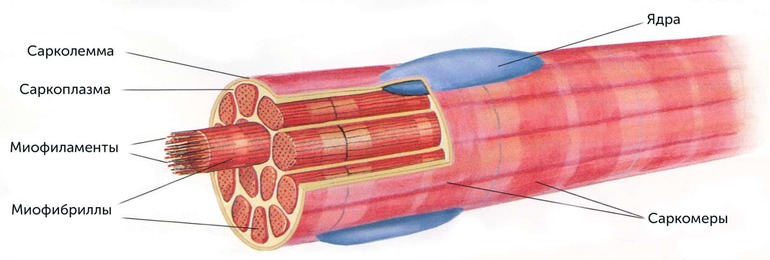
Миофибриллы представляют собой органоиды, состоящие из тонких белковых филаментов. Они располагаются в мышечном волокне и не имеют клеточной оболочки. Миофибриллы окружены саркоплазматическим ретикулумом, состоящим из саркомеров. Связь этих органелл с мышцами обеспечивается при помощи белковых нитей.
Миофибриллы обеспечивают стабильное сокращение мышечных волокон. При воздействии нервных импульсов эти органоиды начинают уменьшаться. В результате сокращения числа органоидов образуется энергия, определяющая силу мышцы. Во время силовых тренировок количество миофибрилл увеличивается. Этот процесс называется гипертрофией. Большие органоиды начинают делиться на несколько маленьких, иначе мышцы не смогут получать достаточное количество энергии.
После травмирования мышечных волокон число миофибрилл уменьшается. Этот процесс ускоряется при наличии гипса, фиксирующего мышцы в определенном положении. Это обусловлено разрушением саркомеров. В результате сокращения количества миофибрилл мышцы получают меньше энергии.

Жгу́тик (англ. Flagellum ) — поверхностная структура, присутствующая у многих прокариотических (бактерий и архей) и эукариотических клеток и служащая для их движения в жидкой среде или по поверхности твёрдых сред.
Жгутики прокариот и эукариот принципиально различаются и по структуре, и по источнику энергии для движения и по механизму движения. Жгутики могут присутствовать на клетках по одному или во множестве, они имеются у многих протистов, зооспор и гамет. Эукариотический жгутик представляет собой вырост клетки, окружённый мембраной, с элементами цитоскелета внутри, который осуществляет биения за счёт энергии гидролиза АТФ. Бактериальный жгутик, как и жгутик архей, вмонтирован в оболочку клетки и вращается за счёт энергии трансмембранного градиента протонов или ионов натрия.
Помимо плавания, жгутики играют важную роль и в других процессах в жизни бактерий. Они участвуют в образовании биоплёнок, обеспечивают контакт клеток с субстратом, облегчают колонизацию хозяина симбиотическими бактериями [en] , служат одним из факторов вирулентности, запускают иммунный ответ организма хозяина.
Читайте также: