
Трансдукция у бактерий кратко
Обновлено: 05.07.2024
11. Механизмы передачи генетического материала у бактерий: конъюгация, трансдукция, трансформация.
Рекомбинация между геномами бактерий осуществляется 3-мя механизмами: конъюгацией, трансдукцией и трансформацией
Трансформация – передача генетической информации через выделенную из клетки-донора ДНК.
1928г. Ф Гриффит (опыт с вирулентнотью пневмококков)
По происхождению ДНК может быть плазмидной или хромосомной и нести гены, трансформирующие реципиента. Подобным путем среди бактериальных популяций могут распространять гены, кодирующие факторы вирулентности, однако в обмене генетической информацией трансформация играет незначительную роль.
Трансформирующей активностью обладает только двунитчатая высокоспирализованная ДНК.
В клетку-реципиент проникает только одна нить ДНК, другая – в клеточной мембране подвергается деградации с освобождением энергии, необходимой для проникновения в клетку второй нити ДНК.
Интеграция с хромосомой требует наличия гомологичных участков с трансформирующей ДНК.
Процесс трансформации зависит от компетентности клетки-реципиента и состояния трансформирующей донорской ДНК.
Компетентность клеток бактерий (способность воспринимать трансформирующую ДНК) зависит от присутствия в ЦПМ особых белков, обладающих специфическим аффинитетом к ДНК. Компетентность зависит от фазы роста бактериальной культуры, вида бактерий. Может увеличиваться при обработке культуры раствором хлорида кальция при пониженной температуре.
Трансформация служит хорошим инструментом для картирования хромосом, поскольку трансформированные клетки включают различные фрагменты ДНК. Определение частоты одновременного приобретения двух заданных характеристик (чем ближе расположены гены, тем более вероятно, что они оба включатся в один и тот же участок ДНК) дает информацию о взаиморасположении соответствующих генов в хромосоме.
Трансформация является основным методом генной инженерии, используемым при конструировании рекомбинантных штаммов с заданным геномом.
Трансдукция – передача бактериальной ДНК посредством бактериофага.
1952г. Дж. Ледебург и Н. Циндер (опыты с сальмонеллами)
Так как трансдуцируются лишь небольшие фрагменты ДНК, вероятность рекомбинации, затрагивающей какой-то определенный признак, очень мала: она составляет от 10 -6 до 10 -8 .
· Общая (неспецифическая) трансдукция – перенос бактериофагом фрагмента любой части бактериальной хромосомы. Феномен неспецифической трансдукции может быть использован для картирования бактериальной хромосомы.
● Специфическая трансдукция – фаговая ДНК интегрирует в бактерию с образованием профага. При исключении ДНК фага из бактериальной хромосомы захватывается прилегающий к месту включения фаговой ДНК фрагмент бактериальной хромосомы. Специфическая трансдукция может служить механизмом переноса вирулентных генов среди бактерий при условии, что эти гены локализованы в непосредственной близости от мест интеграции профага.
● Абортивная трансдукция . При абортивной трансдукции внесенный фрагмент ДНК донора не встраивается в хромосому реципиента, а остается в цитоплазме и там самостоятельно функционирует. Впоследствии он передается одной из дочерних клеток (т.е. наследуется однолинейно) и затем теряется в потомстве.
Обязательное условие – трансмиссивная плазмида ( F , R ), обладающая tra -опероном .
Интегративной трансмиссивной плазмидой является F -фактор . Донорские клетки, имеющие встроенный в хромосому F -фактор, называются Hfr -клетками.
Биологическое значение конъюгации хорошо видно на примере распространения резистентности бактерий к антибиотикам. Устойчивость к антибиотикам бактерия может получить в результате мутации, что происходит 1 раз на каждые 106 клеточных делений. Изменившаяся генетическая информация может быстро распространяться среди сходных бактерий посредством конъюгации.
Трансдукция. Неспецифическая трансдукция. Специфическая трансдукция. Абортивная трансдукция. Феномен лизогении.
Трансдукция — перенос бактериофагом в заражаемую клетку фрагментов генетического материала клетки, исходно содержавшей бактериофаг. Трансдуцирующий бактериофаг обычно переносит лишь небольшой фрагмент ДНК хозяина от одной клетки (донор) к другой (реципиент).
Образовавшийся фаг и есть частица, вызывающая неспецифическую (общую) трансдукцию. При такой форме трансдукции в клетки-реципиенты могут быть внесены практически любые гены.
При неспецифической трансдукции фагом может быть перенесён любой фрагмент ДНК хозяина, а при специфической лишь строго определённые фрагменты ДНК. Наиболее известным примером специфической трансдукции служит трансдукция, осуществляемая фагом. Поскольку этот фаг при переходе в состояние профага включается в хромосому бактерий между генами, кодирующими синтез галактозы и биотина, именно эти гены он может переносить при транедукции. При абортивной трансдукции внесённый фрагмент ДНК донора не встраивается в генофор реципиента, а остаётся в цитоплазме, где его ДНК транскрибируется, но не реплицируется. Это приводит к тому, что при клеточном делении он передаётся только одной из дочерних клеток (то есть наследуется однолинейно) и затем теряется в потомстве.
Свойства трансдуцирующих фаговых частиц следующие:
• Частицы несут лишь часть ДНК фага, то есть не являются функциональными вирусами, а скорее ёмкостями, переносящими фрагменты бактериальной ДНК.
• Подобно прочим дефектным вирусам, частицы не способны к репликации.
• Трансдуцирующие фаги могут содержать какую-либо часть хромосомы хозяина с генами, дающими реципиентной бактерии некоторые преимущества (например, гены устойчивости к антибиотикам или гены, кодирующие способность к синтезу различных веществ). Подобное приобретение бактериями новых свойств получило название феномен лизогении.
• Феномен трансдукции может быть использован для картирования бактериальной хромосомы, если следовать тем же принципам, что и при картировании с использованием феномена трансформации.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Трансдукция – перенос генетического материала из клетки донора в клетку реципиента через трансдуцирующий бактериофаг. Последний представляет собой умеренный фаг, который в состоянии профага получил участок ДНК от донорской клетки в результате неточного вырезания своей последовательности из генома клетки-донора. При этом бактериофаг становится дефектным, т.к. теряет часть собственной нуклеиновой кислоты. Такой фаг упаковывается в свою оболочку, выделяется из клетки и может проникать в клетку-реципиент.
Этот вид рекомбинаций открыт Н. Циндером и Дж. Ледербергом в 1951 г.
Различают 3 вида трансдукции:
Неспецифическая трансдукция. При этом трансдуцирующий бактериофаг передает в реципиентную клетку любой ген донорской клетки и включает его в гомологичную область ДНК реципиента путем рекомбинации этого гена с нуклеоидом. Трансдуцирующий бактериофаг выступает лишь в роли переносчика, в нуклеоид не встраивается, и лизогенизации реципиентной культуры не происходит.
Специфическая трансдукция. Здесь бактериофаг переносит строго определенный ген (или гены) от клетки донора к реципиенту и встраивает его в определенном участке ДНК реципиента путем сайт-специфической рекомбинации. В этом случае бактериофаг может встраиваться в нуклеоид клетки-реципиента, т.е. происходит лизогенизация бактерии. При этом такие клетки становятся невосприимчивыми, как и все лизогенные клетки, к последующему заражению гомологичным вирулентным фагом.
Обычно при специфической трансдукции переносятся бактериальные гены, сцепленные с геномом встроенного бактериофага. Чаще всего они окаймляют (фланкируют) профаг. Для E.coli и фага лямбда это гены gal и bio, контролирующие, соответственно, метаболизм галактозы и синтез витамина биотина.
Абортивная трансдукция. В этом случае фрагмент ДНК донора, доставленный при трансдукции, не включается в ДНК реципиента и остается в цитоплазме. Клетка не лизогенизируется, а новый признак по мере деления клетки исчезает.
5.8.3. Конъюгация
Конъюгация – передача генетического материала из клетки донора в клетку реципиента при непосредственном контакте клеток через цитоплазматический мостик (рис. 9).
Это явление впервые было установлено Д.Ледербергом и Э.Татумом в 1946 г. при совместном культивировании двух штаммов кишечной палочки. В конъгации участвуют клетки, действующие в качестве доноров и реципиентов генетического материала. Перенос генетического материала является односторонним.
Не все клетки могут быть донорскими. Они должны содержать особый репликон, ответственный за конъюгацию – F-фактор (фактор фертильности, половой фактор).
F + клетки были названы генетическими донорами, т.к. они содержат данный фактор. F - - реципиентные клетки не содержат полового фактора, но могут приобрести его в процессе конъюгации.
F-фактор может находиться и в интегрированном в хромосому клетки-донора состоянии. Такая бактерия получила название Hfr-клетки (англ. high frequency of recombination – высокая частота рекомбинации). При этом у донора образуется кольцевая хромосома, включающая F-фактор.
Трансдукция — перенос генов из одной бактериальной клетки в другую при помощи бактериофага. Впервые это явление установили в 1952 г. Н. Зиндер и Дж. Ледерберг. Они проводили исследования на патогенных для мышей бактериях Salmonella typhimurium. Были отобраны два штамма этих бактерий: штамм 22А ауксотрофный, не способный синтезировать триптофан (Т~), и штамм 2А, способный синтезировать триптофан (Т 1 "). Эти штаммы засевали в U-образную трубку, разделенную внизу бактериальным фильтром (рис. 24). В одно колено трубки засевали штамм 22А (Т~), в другое — штамм 2А (Т 1 "). После определенного периода инкубации бактерии штамма 22А при посеве на минимальную питательную среду дали небольшое количество колоний (частота появления трансдуцированных клеток была равна Ы0~ 5 ). Это свидетельствовало о том, что некоторые клетки приобрели способность синтезировать триптофан. Каким же образом бактерии могли приобрести это свойство? Исследования
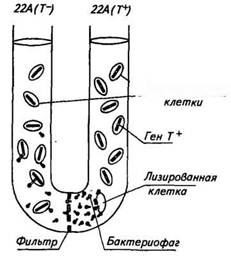
Рис. 24. Схема опыта по трансдукцин
показали, что штамм 22А был лизогенен по фагу Р-22. Этот
фаг освобождался из лизогенной культуры, проходил через
фильтр и лизировал штамм 2А. Присоединив часть генетичес-
кого материала штамма 2А, фаг Бактериальные возвращался обратно и переда вал этот генетический материал штамму 22А. Штамм 22А при
обретал специфические наследственные свойства штамма 2А,
в данном случае свойство синтезировать триптофан. Аналогичным образом могут бытьтрансдуцированы и другие признаки, в том числе способность
к сбраживанию, устойчивость кантибиотикам и т. д.
Явление трансдукции установлено также у кишечной па-лочки и актиномицетов. Как правило, трансдуцируется один ген, реже два и очень редко три сцепленных гена. При переносе генетического материала заменяется участок молекулы ДНК фага. Фаг при этом теряет свой собственный фрагмент и становится дефектным. Включение генетического материала в хромосому бактерии реципиента осуществляется механизмом типа кроссинговера. Происходит обмен наследственным материалом между гомологичными участками хромосомы реципиента и материала, привнесенного фагом.
Различают три вида трансдукции: общую, или неспецифическую, специфическую и абортивную. При неспецифической трансдукции в период сборки фаговых частиц в их головку вместе с фаговой ДНК может включиться любой из фрагментов ДНК пораженной бактерии. В результате в реципиентные клетки могут переноситься различные гены бактерии донора. Неспецифическую трансдукцию могут осуществлять фаги Р-1 и Р-22 у эшерихий, шигелл и сальмонелл. При специфической трансдукции профаг включается в определенное место хромосомы бактерии и трансдуцирует определенные гены, расположенные в хромосоме клетки донора рядом с профагом. Например, фаг "к (лямбда) в состоянии профага всегда включается в одно и то же место в хромосоме кишечной палочки и трансдуцирует локус, обусловливающий способность к сбраживанию галактозы. При отделении профагов от ДНК хозяина прилегающие к профагу бактериальные гены вместе с ним выщепляются из состава хромосомы, а часть генов профага остается в ее составе. Частота общей трансдукции составляет от 1 на 1 млн до 1 на 100 млн. Специфическая трансдукция происходит чаще.
Установлено, что фрагмент хромосомы донора, перенесенный в% клетку реципиента, не всегда включается в хромосому реципиента, а может сохраняться в цитоплазме клетки. При делении бактерий он попадает только в одну из дочерних клеток. Такое состояние получило название абортивной трансдукции.
Трансдукция — перенос генов из одной бактериальной клетки в другую при помощи бактериофага. Впервые это явление установили в 1952 г. Н. Зиндер и Дж. Ледерберг. Они проводили исследования на патогенных для мышей бактериях Salmonella typhimurium. Были отобраны два штамма этих бактерий: штамм 22А ауксотрофный, не способный синтезировать триптофан (Т~), и штамм 2А, способный синтезировать триптофан (Т 1 "). Эти штаммы засевали в U-образную трубку, разделенную внизу бактериальным фильтром (рис. 24). В одно колено трубки засевали штамм 22А (Т~), в другое — штамм 2А (Т 1 "). После определенного периода инкубации бактерии штамма 22А при посеве на минимальную питательную среду дали небольшое количество колоний (частота появления трансдуцированных клеток была равна Ы0~ 5 ). Это свидетельствовало о том, что некоторые клетки приобрели способность синтезировать триптофан. Каким же образом бактерии могли приобрести это свойство? Исследования

Рис. 24. Схема опыта по трансдукцин
показали, что штамм 22А был лизогенен по фагу Р-22. Этот
фаг освобождался из лизогенной культуры, проходил через
фильтр и лизировал штамм 2А. Присоединив часть генетичес-
кого материала штамма 2А, фаг Бактериальные возвращался обратно и переда вал этот генетический материал штамму 22А. Штамм 22А при
обретал специфические наследственные свойства штамма 2А,
в данном случае свойство синтезировать триптофан. Аналогичным образом могут бытьтрансдуцированы и другие признаки, в том числе способность
к сбраживанию, устойчивость кантибиотикам и т. д.
Явление трансдукции установлено также у кишечной па-лочки и актиномицетов. Как правило, трансдуцируется один ген, реже два и очень редко три сцепленных гена. При переносе генетического материала заменяется участок молекулы ДНК фага. Фаг при этом теряет свой собственный фрагмент и становится дефектным. Включение генетического материала в хромосому бактерии реципиента осуществляется механизмом типа кроссинговера. Происходит обмен наследственным материалом между гомологичными участками хромосомы реципиента и материала, привнесенного фагом.
Различают три вида трансдукции: общую, или неспецифическую, специфическую и абортивную. При неспецифической трансдукции в период сборки фаговых частиц в их головку вместе с фаговой ДНК может включиться любой из фрагментов ДНК пораженной бактерии. В результате в реципиентные клетки могут переноситься различные гены бактерии донора. Неспецифическую трансдукцию могут осуществлять фаги Р-1 и Р-22 у эшерихий, шигелл и сальмонелл. При специфической трансдукции профаг включается в определенное место хромосомы бактерии и трансдуцирует определенные гены, расположенные в хромосоме клетки донора рядом с профагом. Например, фаг "к (лямбда) в состоянии профага всегда включается в одно и то же место в хромосоме кишечной палочки и трансдуцирует локус, обусловливающий способность к сбраживанию галактозы. При отделении профагов от ДНК хозяина прилегающие к профагу бактериальные гены вместе с ним выщепляются из состава хромосомы, а часть генов профага остается в ее составе. Частота общей трансдукции составляет от 1 на 1 млн до 1 на 100 млн. Специфическая трансдукция происходит чаще.
Установлено, что фрагмент хромосомы донора, перенесенный в% клетку реципиента, не всегда включается в хромосому реципиента, а может сохраняться в цитоплазме клетки. При делении бактерий он попадает только в одну из дочерних клеток. Такое состояние получило название абортивной трансдукции.
Генетическая трансдукция — это перенос ДНК бактерий из одной клетки в другую с помощью бактериофага. В этом процессе ведущая роль принадлежит вириону, который является трансдукционной частицей. В результате происходит изменение наследственных признаков.
Генетическая трансдукция — процесс переноса бактериальной ДНК из одной клетки в другую.
История исследования
Процесс перемещения ДНК был установлен американскими исследователями Д. Ледербергом и Н. Циндером в 1952 г. Ученые изучали функции и строение бактерии Salmonella typhimurium и фага Р22. В результате проведенных опытов они пришли к выводу: происходит генетический обмен между микроорганизмами, не способными к синтезу соединений, и прототрофами.
Между встраивающимися генами не наблюдается физического контакта, процесс не идентичен трансформации. Открытие ученых оказалось случайным, один из штаммов был подвергнут состоянию дезорганизации благодаря вмешательству вириона Р22.
В ходе эксперимента в клетке наблюдали вегетативный рост профага. Через некоторое время образовавшиеся частицы фага проникали в ауксотрофный вид и переносили в его клетки гены своего прежнего хозяина.
Реакции фагов
Вирионы, избирательно уничтожающие бактериальные клетки, перемещают генетический материал от донора, в котором размножался возбудитель, к реципиенту, зараженному фагом. Вирус Х переносит лишь избранные гены — gal и Ыо. Носитель вирусных частиц проникает в фазу генома фага, внедренную в хромосомную часть ДНК зараженной клетки.
Вирионы перемещают генетический материал от донора к реципиенту.
Вирусная ДНК отделяется от бактериальной нуклеопротеидной структуры, несущей наследственную информацию. Возможно ошибочное деление вирусной ДНК. В этом случае ее фрагмент сохранится в нуклеопротеиде, но гены основной зиготы будут уничтожены. Главная причина появления неточности — неправильное перераспределение генетического материала.
Фаги уничтожают бактериальную структуру, используя литический или лизогенный метод,
Генетика фагов изучает мутации вирионов Та и Ти, а также физические и химические свойства фагов F1 и I2. Мутанты вирионов чувствительны к колебаниям температуры. К ним принадлежат фаг ts и холодочувствительный вирус es. Они размножаются и разрушают клетки при высокой температуре.
Литическая
Клеточный процесс сложный. Вирионы Т (Т2, Ти, Т6) содержат в ДНК вещество 5-гидроксиметилцитозин. Измененные формы фага способны к репродукции за счет заблокированных фаз. В вирионах-мутантах различные части расположены в четкой последовательности.
Фаги не способны двигаться. В результате взаимодействия вириона и взвеси бактерий частицы фага после столкновения с клетками остаются на их поверхности и вводят в структурную единицу бактерии свою ДНК.
Через несколько часов часть клеток разрушается. Вирусы, участвующие в литической фазе, становятся вирулентными.
На последней стадии литического процесса протеиновые частицы окружают нуклеиновую кислоту, формируя новые фаги.
Лизогенная
Умеренные вирусы взаимодействуют с микроорганизмами. Нуклеиновая кислота, находящаяся внутри бактериальной структурной единицы, внедряется в геном. В течение длительного периода наблюдают сосуществование вируса и клетки, не сопровождающееся гибелью структурной единицы.
Вирусы взаимодействуют с микроорганизмами.
При изменении условий среды вирус проникает в клетку. Начинается развитие активной стадии заболевания. Клетка, содержащая геном вириона, встраивающийся в хромосомную ДНК бактерии, называется лизогенной. Процесс сопровождается мутацией наследственной структурной единицы.
Умеренный фаг обладает способностью к заражению, которая определяется ДНК-связывающими свойствами белка. Лизогения меняет наследственные признаки бактериальной клетки.
Трансдукционное картирование
При переносе бактериальной ДНК вирионы размножаются на чувствительных клетках донора. Для мутагенного ДНК-связывающего белка, содержащего фрагмент нуклеопротеида и несущего наследственную информацию, проводят идентификацию донора и хозяина по нескольким признакам. Для установления сцепления используют отборное скрещивание.
Вирус размножают на штамме, имеющем фенотип Itr+Leu+, и заражают им клетки хозяина, которые нуждаются в аминокислотах.
После обработки вирионом структурные единицы организма помещают в питательную среду, не содержащую треонина. В питательной смеси не происходит рост гибридной нуклеиновой кислоты, но образуются структурные единицы, получившие часть хромосомы донора с многофакторным наследственным материалом.
На этом же участке может располагаться второй ген, встречаются двойные Iht+Leu+. Их можно обнаружить, поместив колонии трансдуктантов в питательную среду, не содержащую лейцина. Аминокислоту используют как неселективный отборный материал, необходимый для установления котрансдукции.
Общая (неспецифическая) трансдукция
Вирус Р1, находящийся в структурной единице хозяина в виде плазмиды, а также фаги Р22 и Ми осуществляют неспецифическое перемещение бактериальной ДНК. Вирусы Р22 и Ми встраиваются в любой участок нуклеопротеидной структуры инфицированной зиготы.
Вирусы осуществляют неспецифическое перемещение бактериальной ДНК.
Усиление генной структуры фага сопровождается проникновением части ДНК бактерии в его структуру. ДНК вириона в капсиде отсутствует. Величина этого участка равна длине стандартной вирусной ДНК.
Проникая в иную клетку, часть ДНК попадает в состав генетического материала после соответствующей рекомбинации. Перенесенные вирусом небольшие молекулы ДНК образуют кольцо и подвергаются репликации в новой бактериальной структурной единице.
События в клетке реципиента
Частицы, появившиеся в результате трансдукции, доставляют ДНК в клетки бактерий. Инфекционный процесс не развивается, т.к. наследственные факторы, отвечающие за воспроизведение фага, отсутствуют.
ДНК, принадлежащая донору, придерживается одного из двух путей развития: прикрепляется к хромосоме или остается в цитоплазме. Если часть ДНК не способна к процессу перекодирования, через некоторое время она разрушается.
Иногда отрезок ДНК удваивается самостоятельно и передается в другие поколения как носитель наследственных признаков, находящихся вне хромосомы.
Вирусы Р22 и Р1 принимают участие в двухниточной замене частей бактериальной хромосомы на вирусную ДНК. В клетку реципиента в процессе трансдукции попадает лишь 5% всей нуклеиновой кислоты.
Вирусы принимают участие в замене частей бактериальной хромосомы.
Использование
Практическое применение вирусов основано на литическом воздействии фагов на бактерии. Вирион используют в следующих случаях:
- для лечения инфекционных болезней;
- с целью идентификации микроорганизмов, вызывающих холеру, сальмонеллез, дифтерию.
Вирионы используют для изучения генетики микроорганизмов. Лечение заболеваний антибиотиками не всегда приводит к желаемому результату, поэтому применяют фаготерапию.
Вирионы применяют в сельском хозяйстве, животноводстве, растениеводстве.
Бактериальные фаги имеют следующие достоинства: не влияют на рост микрофлоры живого организма, уничтожают устойчивые к лечению АБ патогенные микроорганизмы, не оказывают воздействия на формирование эукариотических клеток.
Специфическая трансдукция
Ученые изучили перенос бактериальной ДНК вирусом Л. Фаг располагается на att-участке нуклеопротеидной структуры Е.coli, имеющей одинаковую последовательность фосфорных эфиров.
В период выведения вириона его исчезновение часто сопровождается ошибкой: удаляется часть, по размеру равная фрагменту ДНК. Особенность процесса состоит в том, что начало обломка располагается в другом месте. Некоторые гены вируса исчезают, другие наследственные факторы, несущие информацию, поглощаются фагом.
Некоторые фаги внедряются в любую часть нуклеопротеиновой структуры и переносят наследственную информацию, используя механизм специфического перемещения бактериальной ДНК из одной клетки в другую.
В хромосоме находится пространство, имеющее общие черты с att-участком и ДНК вируса. При разрушении гомологичного ресурса осуществляют внедрения фага в нуклеопротеид и его передачу при специфическом перемещении генов, расположенных рядом.
Фаги переносят наследственную информацию.
Формирование частиц фага лямбда
Вирус принадлежит к умеренным фагам E.coli, обладающим двухцепочной ДНК. Фаг внедряется в генную структуру бактерий в виде профага и передается нескольким поколениям.
Вирион лямбда состоит из следующих элементов: головки, хвоста, боковых нитей, белковой пленки. Внедрение сопровождается генетическим перераспределением материала на участке бактерии. В результате обмена наследственной информацией в присутствии вирусного и бактериального белка формируется форма Холлидея.
Вирус лямбда имеет 2 пути развития — мутагенный и литический. Фаг образует новые частицы, которые попадают во внешнюю среду, а клетка хозяина гибнет. В случае развития лизогенной реакции геном вириона проникает в хромосому E.coli и передается нескольким поколениям.
Вирус лямбда используют как вектор для клонирования, применяют для лечения холеры, дизентерии.
Какие события происходят в клетке реципиента
Вирус Л перемещает хромосомы от клеток донора к реципиенту. Ученые вывели геном фага, внедренного в хромосомную ДНК бактерии в культуре клеток кишечной бактерии К 12. В процессе получили расщепленные вирусом структурные элементы. Ими заражали культуры типа К 12. Затем их высевали в питательные среды и учитывали количество колоний, проверяя наличие аллелей от клеток-доноров, передавшихся мутантным реципиентам.

Ученые вывели геном фага, внедренного в хромосомную ДНК.
Вирион может переносить часть хромосомы, но процесс специфический, его осуществляет геном gal, находящийся на хромосоме бактериальной структуры. Многие трансдуктанты Cal+ нестабильны, теряют этот признак.
Абортивная трансдукция
При этом виде перемещения часть бактериального наследственного фактора не обменивается. Геном располагается в клетке, но не размножается. В случае деления клетки эта часть переходит в дочерние образования. После нескольких делений все клетки потомства остаются без этого фрагмента.
Процесс абортивного перемещения установлены Дж. Ледербергом в 1953 г. Он доказал, что часть хромосомы, оказавшейся в другой бактериальной зиготе, находится в клетке реципиента в виде непродуктивной частицы и передается 1 дочерней единице, а затем исчезает в потомстве.
Трансдукции клеток млекопитающих с вирусными векторами
Перенос фаговых векторов применяют для модификации наследственных факторов в клетках млекопитающих. В небольших молекулах ДНК гены локализуют в установленных местах вирусные частицы.
Плазмиды внедряют методом фланирования в структуру, образующуюся вместе с другими частями ДНК-конструкции, которые снабжены вирусными частицами, создающими инфекционные полноценные комплексы из нуклеиновой кислоты и капсида.
Для обеспечения безопасности процесса каждая плазмида содержит все данные, необходимые для формирования фага. Одновременное воспроизведение вируса в 2-3 плазмидах приводит к образованию инфекционного вириона.
Векторы, существующие в семействе палочковидных вирусов, могут синтезировать эукариотические протеины в клетках насекомых. Бакуловирус передает наследственную информацию зиготе млекопитающих, он не аутопродуктивен, а векторы, созданные на его основе, безопасны.
Векторы вирусов млекопитающих поражают гены в клетках-мишенях и используются как естественные градиенты для перемещения ДНК в эукариотические клетки. Учитывают такие факторы, как простота использования, регуляция во времени, скорость образования протеинов эукариотов.
Применяют векторы, созданные на основе аденовируса, ретровируса, возбудителя герпеса, лентовирусов.
Система имеет недостатки:
- неадекватность модификаций;
- большая чувствительность бакуловирусов.
Векторы, созданные на их основе, используют в биологии, биомедицине, для вакцинации и в генной терапии.
Читайте также: