
Полное и неполное дробление кратко
Обновлено: 05.07.2024
У разных видов животных дробление имеет свою особенную организацию, характер которой определяется прежде всего строением яйца, в частности распределением и количеством желтка, а также положением митотического веретена в бластомерах относительно анимально-вегетативной оси яйца.
Дробление бывает полным — голобластическим, неполным — меробластическим и поверхностным — абластическим. Полным называется дробление, при котором яйцо делится на бластомеры целиком. При неполном дроблении деления затрагивают только часть яйца, тогда как другая часть, обычно с высокой концентрацией желтка, не делится. При поверхностном типе дробления яйцеклетка не делится вовсе, и дело ограничивается лишь кариокинезом — делениями ядер.
Голобластическое дробление . Такое дробление характерно для яиц, содержащих сравнительно мало желтка, т. е. для олиго- и мезолецитальных, а также для умеренно телолецитальных яиц. Если в результате дробления образуются бластомеры примерно одинаковых размеров, говорят о равномерном дроблении, если же бластомеры явно различаются по величине — о неравномерном. Неравномерность дробления может быть связана с концентрацией желтка в вегетативном полушарии. Иногда она обусловлена сосредоточением в отдельных бластомерах больших объемов специализированной цитоплазмы, например цитоплазмы полярной лопасти у некоторых моллюсков, или иными причинами, как в случае образования микромеров у морского ежа.
Меробластическое дробление . Этот тип дробления наблюдается у животных с телолецитальными яйцами, которые отличаются высокой степенью концентрации желтка в вегетативной области. У головоногих моллюсков, многих рыб, а также у рептилий и птиц дробление происходит только в относительно небольшой части яйца, образующей как бы диск на поверхности яйцеклетки, — дискоидальное дробление.
Абластическое дробление . Такое дробление характерно для центролецитальных яиц насекомых, поэтому его называют также централецитальным. В этом случае не происходит цитокинеза и деления цитоплазмы. Делятся только ядра, которые находятся в центральной области яйца, откуда они мигрируют по цитоплазматическим тяжам, пронизывающим яйцо, на поверхность. Длительное время зародыш имеет синцитиальную структуру. Попав в поверхностную цитоплазматическую бластему, или периплазму ядра образуют синцитиальную бластодерму, которая позднее целлюляризуется и дает начало клеточной бластодерме зародыша.
По характеру расположения бластомеров в развивающемся зародыше различают несколько типов дробления, в частности радиальное, бирадиальное, спиральное, билатеральное, ротационное и неупорядоченное дробление, а также табличную палинтомию и полиаксиальный тип дробления.
Радиальное дробление . У многих животных (книдарии, иглокожие, некоторые первично-хордовые, рыбы и амфибии) дробящееся яйцо имеет радиальную ось симметрии, при которой плоскость, проходящая через любой меридиан, делит зародыш на две геометрически тождественные половины. При радиальном дроблении два первых деления проходят во взаимно перпендикулярных меридиональных плоскостях, а третье — в экваториальной плоскости. Последующие деления чередуются в широтной и меридиональной плоскостях. Если третье деление происходит в экваториальной плоскости, то дробление равномерное, если же плоскость этого деления смещена в анимальное полушарие, то дробление неравномерное и ведет к образованию микромеров в анимальном и макромеров в вегетативном полушариях.
Спиральное дробление . У аннелид и моллюсков в результате первых двух взаимно перпендикулярных меридиональных делений образуется стадия четырех бластомеров. Начиная с третьего деления дробления митотические веретена располагаются под некоторым углом к меридиональной плоскости. Благодаря этому образующиеся четыре клетки анимального полушария несколько смешаются относительно клеток вегетативного квартета и располагаются в промежутках между его бластомерами квадрантами. Если смещение происходит по часовой стрелке (при наблюдении с анимального полюса), дробление называют дексиотропным (от лат. dexter — правый), если же смещение происходит в противоположном направлении, дробление называют леотропным. При последующих делениях наклоны веретен чередуются: за дексиотропным следует леотропное деление и наоборот. В случае спирального дробления его неравномерность может обнаружиться уже после первого деления дробления. Если на стадии 4 бластомеров все клетки одинаковых размеров, говорят о гомоквадрантном дроблении, если же они различаются по размерам, то — о гетероквадрантном.
Неравномерностъ дробления может проявляться и вдоль анимально-вегетативной оси: при достаточно больших запасах желтка основной (вегетативный) квартет представлен макромерами, а анимальный — микромерами. В ходе спирального дробления бластомеры занимают строго фиксированное положение в системе и обозначаются специальными индексами.
Билатеральный тип дробления . Этот тип дробления характерен для нематод, а также для многих низших хордовых, в том числе для асцидий, аппендикулярий и бесчерепных. Характерной его особенностью является раннее проявление билатеральной симметрии. Например, у оболочников подразделение на левую и правую части происходит уже при первом меридиональном делении дробления, плоскость которого рассекает желтый серп оплодотворенного яйца на две симметричные половины. Билатеральность становится очевидной, когда вторая, тоже меридиональная борозда отделяет крупные передние бластомеры от задних более мелких клеток.
Некоторые авторы выделяют в особый, ротационный, тип дробления млекопитающих, у которых бластомеры при втором дроблении делятся во взаимно перпендикулярных плоскостях. Наконец, у некоторых животных описано вращение бластомеров. Изменение положения бластомеров относительно анимально-вегетативной оси, вероятно, характерно для ряда кишечнополостных. Согласно некоторым исследованиям, у книдарий иногда наблюдается слабая взаимосвязь между клетками, которая приводит к возникновению так называемого анархического, или неупорядоченного дробления.
Что определяет характер дробления? От чего зависит положение плоскости дробления? Что является причиной того или иного расположения митотического веретена? Конечно, характер дробления, как мы видели, отчасти определяется особенностями распределения желтка в яйцеклетке: увеличение концентрации желтка обычно приводит к замедлению образования борозд в вегетативном полушарии или даже к полному прекращению делений. В этих случаях наблюдается меробластическое или абластическое дробление.
Вместе с тем очевидно, что особенности того или иного типа дробления являются результатом длительной эволюции и контролируются не только концентрацией желтка, но многими факторами, природа которых во многом остается еще не раскрытой.
Из опытов Герстадиуса вытекало два важных следствия. Во-первых, они свидетельствовали, что в дробящемся яйце имеется некий счетчик времени, который, будучи независимым, тем не менее сопряжен с процессами, непосредственно контролирующими кариокинез. Во-вторых, эти опыты указывали на существование механизма, который предопределяет последовательный ряд событий, реализующих генетически детерминированную ориентацию веретен деления.
Важный вклад в понимание проблемы дробления примерно в то же время внесла американский эмбриолог Этель Гарвей (1885-1968), работы которой также показали, что механизмы кариокинеза и цитокинеза относительно автономны и могут быть разобщены в эксперименте. Центрифугируя неоплодотворенные яйца морского ежа в растворе сахарозы той же плотности, что и яйца, она смогла разделить яйцо на две половины — мерогоны. В легкой половине оставалось ядро, а в тяжелой концентрировался желток и пигментные зерна. После активации гипертонической морской водой наблюдалась партеногенетическая мерогоиия: безъядерные половинки начинали дробиться и формировали аномальные бластулы, которые после вылупления погибали, поскольку составляющие их клетки не имели ядер.
Относительная автономия кариокинеза и цитокинеза обусловлена тем, что в их основе лежат различные клеточные механизмы. Поскольку эти механизмы и, в частности, роль цитоскелета в осуществлении клеточного деления подробно рассматриваются в курсе цитологии, здесь мы ограничимся лишь кратким упоминанием о них.
Кариокинез происходит при участии митотического веретена, основу которого составляют микротрубочки, построенные из белка тубулина. Цитокинез осуществляется сократительным кольцом, которое образуется в области борозды дробления из актиновых микрофиламентов. Если дробящееся яйцо обработать ингибитором сборки микротрубочек (например, колхицином или нокодазолом), то кариокинетические деления блокируются в метафазе. При действии ингибиторов формирования микрофиламентов (например, цитохалазина) деление ядер сохраняется, но подавляется цитокинез.
При нормальном развитии образование веретена и борозды дробления скоординированы. Уже давно была отмечена связь между положением митотического веретена и направлением борозды дробления: последняя всегда проходит в плоскости метафазной пластинки, перпендикулярно длинной оси веретена. Экспериментальные воздействия, ведущие к изменению положения веретена, неизбежно вызывают смещение борозды дробления в соответствии с новым положением веретена. Неудивительно поэтому, что появление сверхчисленных веретен (например, при полиспермии) резко нарушает нормальный ход развития, а разрушение звезд веретена ведет к остановке дробления. Возможно, однако, что образование борозды дробления зависит главным образом от взаимодействия двух звезд, поскольку в экспериментальных условиях удается вызвать появление борозды между звездами, не связанными между собой веретеном. Можно предполагать, что область формирования сократительного кольца микрофиламентов определяется взаимодействием микротрубочек звезд с кортексом яйца.
Положение митотического веретена, а следовательно, и звезд зависит не только от элементов цитоскелета, но и от других цитоплазматических факторов, хотя реальный механизм их действия пока не известен. Например, у животных со спиральным типом дробления обнаружено, что ориентация плоскости третьего деления в правую или левую сторону (т. е. дексиотропность или леотропность) контролируется цитоплазматическим фактором, который синтезируется под контролем генов материнского организма в период оогенеза. Значение этого фактора было показано как в опытах, в которых цитоплазму из яиц одного типа инъецировали в яйца другого типа, так и в генетических исследованиях закрученности раковины у моллюска Lymnea, которая коррелирует с ориентацией веретена при дроблении. Так, у Lymnea аллель правозакрученности доминантен (D), а левозакрученности рецессивен (d). При скрещивании самок DD с самцами dd все потомки Dd были правозакрученными. При скрещивании самцов DD с самками dd все потомки Dd были левозакрученными, а при скрещивании самцов Dd с самками Dd потомки (1DD: 2Dd: 1dd) были все правозакрученными.
Прямые доказательства участия генетических факторов в определении характера дробления получены на нематоде Caenorhabditis elegans, у которой первое деление дробления дает крупный передний бластомер АВ и задний бластомер Р1 меньших размеров. При втором делении образуется характерная Т-образная фигура, так как оси митотических веретен в указанных бластомерах располагаются взаимно перпендикулярно. Асимметрия деления яйца у С, elegans определяется белками PAR и МЕХ, локализованными в кортикальном слое. Обнаружено 6 генов par (partitioning-defective). У большинства мутантов par первое деление равномерное, а веретена дочерних клеток расположены вдоль одной оси, а не взаимно перпендикулярны. Эти нарушения коррелируют с изменениями локализации белков PAR. У животных дикого типа белки PAR-3 и PAR-6 локализованы в передней области кортекса, тогда как PAR-1 и PAR-2 — в задней. Отсутствие одного из белков передней группы ведет в распространению задних факторов вперед. В отсутствии PAR-2 факторы переднего комплекса распространяются назад.
В настоящее время эмбриология не располагает данными, которые позволили бы дать сколько-нибудь связный очерк механизмов, лежащих в основе формирования тех или иных конкретных типов дробления. Тем не менее даже те отрывочные сведения, которые имеются в нашем распоряжении, позволяют утверждать, что тип дробления определяется сложным комплексом факторов, в том числе генетическими и клеточными механизмами, которые контролируют и интегрируют многие элементарные процессы. Среди последних особое значение имеют события, связанные с синтезом ДНК, организацией митотического деления ядра (кариокинез) и делением цитоплазматического тела клетки (цитокинез). Относительная автономия этих элементарных процессов создает предпосылки для формирования в ходе эволюции животных разнообразных типов дробления. Например, в основе возникновения поверхностного типа дробления лежит возможность разобщения кариокинеза и цитокинеза.
Сейчас же после оплодотворения зигота разделяется на 2 клетки, к-е вновь делятся; образовавшиеся 4 клетки делятся на 8 и т.д. Деления следуют одно за другим настолько быстро, что клетки не успевают расти и становятся все мельче. Т.к. увеличение числа клеток при дроблении сопровождается уменьшением их размеров, общий объем зародыша почти не меняется.
Клетки, возникающие в результате дробления, называются бластомерами, а те перетяжки по к-м они отделяются одна от другой, - бороздами дробления. Последние могут иметь различное направление, и соответственно этому различают борозды:
- меридиональные, к-е проходят от анимального полюса к вегетативному;
- экваториальные, пересекающие зиготу по экватору;
- широтные, параллельные экватору;
- тангенциальные, проходящие параллельно поверхности зиготы.
Правила дробления. Первое правило отражает местоположение веретена в бластомере, а второе – его направление. К этим двум правилам следует добавить еще третье: скорость прохождения борозд дробления всегда обратно пропорциональна кол-ву желтка в клетке.
Дробление у разных многоклеточных протекает различно. Это зависит от особенностей строения яйцеклеток, прежде всего от кол-ва и распределения в них желтка. При отсутствии желтка, при небольшом или даже среднем его кол-ве дробится вся оплодотворенная яйцеклетка. Чем больше в ней желтка, тем сильнее затрудняется прохождение борозд дробления. При значительном кол-ве желтка дробится только часть зиготы, к-я свободна от него. Поэтому различают дробление полное и неполное. Соответственно этому яйца разделяются на голобластические , дробящиеся полностью, и меробластические – с частичным дроблением.
Полное дробление м.б. равномерным и неравномерным. Это также обусловливается кол-вом и расположением желтка в яйцеклетке. Если его мало и он распределяется по всему яйцу, борозды дробления проходят с одинаковой скоростью на всем протяжении клетки и разделяют ее на равные бластомеры. Если желток в яйцеклетке размещается неравномерно, то участки, где его много, дробятся медленнее, чем участки, к-е бедны им. В результате получаются неравные бластомеры: мелкие –в анимальном полушарии (микромеры) и крупные – в вегетативном (макромеры). В таком случае дробление хотя и остается полным, но будет уже неравномерным.
Полное равномерное дробление наблюдается у ланцетника. Его яйцо относится к гомолецитальному типу: оно содержит небольшое кол-во желтка, равномерно распределенного по всей клетке. Как у б-ва животных, яйцеклетка ланцетника слегка сплюснута в направлении полюсов. В связи с такой формой наибольшая протяженность ее находится в горизонтальной плоскости, поэтому согласно правилам дробления 1-е веретено ложится горизонтально, а борозда дробления проходит вертикально. В каждом из образовавшихся бластомеров наибольшая протяженность ЦП сохраняется в горизонтальной плоскости, поэтому 2-я борозда дробления проходит опять меридионально. После 2-го дробления направление наибольшей протяженности ЦП в бластомерах меняется. Соответственно этому веретено дробления становится вертикально, и 3-я борозда проходит в экваториальной плоскости. Теперь уже отделяются 4 верхних (анимальных) бластомера от 4 нижних (вегетативных). 4-е дробление опять меридиональное. После него происходит правильное чередование поперечных (экваториальных и широтных) дроблений с продольными (меридиональными). Кол-во бластомеров увеличивается в отношениях, кратных 2: первые 2→ 4→ 8→ 16→ 32→ 64 и т.д. Бластомеры разных полушарий сохраняют почти одинаковые размеры.
В результате дробления получается многоклеточный зародыш в форме полого шара, к-й называется бластулой. В ней различают стенку, состоящую из большого кол-ва мелких клеток – бластодерму и полость – бластоцель (первичная полость тела). Анимальную часть бластулы называют крышей бластулы, а вегетативную – дном бластулы. Полное и равномерное дробление у ланцетника завершается всегда образованием бластулы, стенка к-й состоит из одного слоя клеток, а полость лежит в центре. Такая бластула называется целобластулой.

Полное неравномерное дробление наблюдается у земноводных, яйца к-х относятся к телолецитальным, со средним кол-вом желтка. Последний распределяется по всему яйцу, но большая часть его сосредоточена всегда в вегетативной половине, где и зерна его крупнее. В оплодотворенном яйце лягушки отчетливо выражена полярная дифференцировка, к-я проявляется в резкой пигментации анимальной половины. Первые две борозды дробления проходят меридионально. Они быстро разделяют анимальную часть, значительно задерживаясь в вегетативной. 3-я борозда, проходя параллельно экваториальной плоскости, разделяет бластомеры поперек. Она располагается ближе к анимальному полюсу. Это происходит потому, что веретено дробления смещается от от вегетативного полушария, где сосредоточена основная часть желтка, к анимальному. Верхние бластомеры получаются меньших размеров, чем нижние. При дальнейшем дроблении анимальные бластомеры дробятся заметно быстрее вегетативных, и поэтому разница между их размерами становится все более резкой.
Неполное дробление сопровождается делением только той части яйца, к-я свободна от желтка; часть, заполненная им, не дробится. По этому типу развиваются телолецитальные яйца (у костистых рыб, птиц, пресмыкающихся) и центролецитальные (у насекомых). Соответственно особенностям строения этих яиц различают дискоидальное и поверхностное дробление.
Дискоидальное дробление наблюдается у костистых рыб, птиц, пресмыкающихся. Яйца этих животных богаты желтком, большое кол-во к-го обусловливает их значительные размеры. Свободная от него ЦП, в виде небольшого участка находится в верхней части яйца и называется зародышевым диском. Т.к. дробится только последний, дробление получило название дискоидального.
Толщина зародышевого диска очень незначительна, он распластан на желтке, и его наибольшая протяженность совпадает с плоскостью, параллельной поверхности яйца. Поэтому веретена при первых 3-х и даже 4-х дроблениях располагаются горизонтально, а борозды дробления проходят вертикально. В результате получается 1 ряд клеток. После нескольких делений они становятся высокими и веретена располагаются в них вертикально, а борозды дробления – параллельно поверхности яйца. В связи с этим в зародышевом диске появляются поверхностные клетки и клетки, расположенные на желтке. Последующие деления проходят в самых различных направлениях, и зародышевый диск превращается в пластинку, состоящую из нескольких рядов клеток. Между диском и желтком возникает небольшая щелевидная полость, к-ю приравнивают к бластоцелю.
Поверхностное дробление наблюдается в центролецитальных яйцах с большим кол-вом желтка в их середине. ЦП в таких яйцах располагается по периферии клетки в центре, около ядра. Через массу желтка проходят тонкие ЦП-кие тяжи, соединяющие периферическую ЦП с околоядерной. Дробление начинается с деления ядра и обособления вокруг возникших ядер ЦП. Число ядер увеличивается. Окруженные ЦП, они постепенно передвигаются к периферии яйцеклетки. Как только ядра попадают в ее поверхностный слой, последний разделяется соответственно их кол-ву на бластомеры. Т.о., в результате дробления вся центр. часть ЦП перемещается к поверхности и сливается с периферической. Образуется сплошная бластодерма, из к-й развивается зародыш. Поверхностное дробление свойственно почти исключительно яйцам членистоногих.
После оплодотворения яйцеклетка начинает делиться. Многократное деление зиготы называется дроблением. Оно приводит к образованию многоклеточного зародыша.
Сейчас же после оплодотворения зигота разделяется на 2 клетки, к-е вновь делятся; образовавшиеся 4 клетки делятся на 8 и т.д. Деления следуют одно за другим настолько быстро, что клетки не успевают расти и становятся все мельче. Т.к. увеличение числа клеток при дроблении сопровождается уменьшением их размеров, общий объем зародыша почти не меняется.
Клетки, возникающие в результате дробления, называются бластомерами, а те перетяжки по к-м они отделяются одна от другой, - бороздами дробления. Последние могут иметь различное направление, и соответственно этому различают борозды:
- меридиональные, к-е проходят от анимального полюса к вегетативному;
- экваториальные, пересекающие зиготу по экватору;
- широтные, параллельные экватору;
- тангенциальные, проходящие параллельно поверхности зиготы.
Правила дробления. Первое правило отражает местоположение веретена в бластомере, а второе – его направление. К этим двум правилам следует добавить еще третье: скорость прохождения борозд дробления всегда обратно пропорциональна кол-ву желтка в клетке.
Дробление у разных многоклеточных протекает различно. Это зависит от особенностей строения яйцеклеток, прежде всего от кол-ва и распределения в них желтка. При отсутствии желтка, при небольшом или даже среднем его кол-ве дробится вся оплодотворенная яйцеклетка. Чем больше в ней желтка, тем сильнее затрудняется прохождение борозд дробления. При значительном кол-ве желтка дробится только часть зиготы, к-я свободна от него. Поэтому различают дробление полное и неполное. Соответственно этому яйца разделяются на голобластические , дробящиеся полностью, и меробластические – с частичным дроблением.
Полное дробление м.б. равномерным и неравномерным. Это также обусловливается кол-вом и расположением желтка в яйцеклетке. Если его мало и он распределяется по всему яйцу, борозды дробления проходят с одинаковой скоростью на всем протяжении клетки и разделяют ее на равные бластомеры. Если желток в яйцеклетке размещается неравномерно, то участки, где его много, дробятся медленнее, чем участки, к-е бедны им. В результате получаются неравные бластомеры: мелкие –в анимальном полушарии (микромеры) и крупные – в вегетативном (макромеры). В таком случае дробление хотя и остается полным, но будет уже неравномерным.
Полное равномерное дробление наблюдается у ланцетника. Его яйцо относится к гомолецитальному типу: оно содержит небольшое кол-во желтка, равномерно распределенного по всей клетке. Как у б-ва животных, яйцеклетка ланцетника слегка сплюснута в направлении полюсов. В связи с такой формой наибольшая протяженность ее находится в горизонтальной плоскости, поэтому согласно правилам дробления 1-е веретено ложится горизонтально, а борозда дробления проходит вертикально. В каждом из образовавшихся бластомеров наибольшая протяженность ЦП сохраняется в горизонтальной плоскости, поэтому 2-я борозда дробления проходит опять меридионально. После 2-го дробления направление наибольшей протяженности ЦП в бластомерах меняется. Соответственно этому веретено дробления становится вертикально, и 3-я борозда проходит в экваториальной плоскости. Теперь уже отделяются 4 верхних (анимальных) бластомера от 4 нижних (вегетативных). 4-е дробление опять меридиональное. После него происходит правильное чередование поперечных (экваториальных и широтных) дроблений с продольными (меридиональными). Кол-во бластомеров увеличивается в отношениях, кратных 2: первые 2→ 4→ 8→ 16→ 32→ 64 и т.д. Бластомеры разных полушарий сохраняют почти одинаковые размеры.
В результате дробления получается многоклеточный зародыш в форме полого шара, к-й называется бластулой. В ней различают стенку, состоящую из большого кол-ва мелких клеток – бластодерму и полость – бластоцель (первичная полость тела). Анимальную часть бластулы называют крышей бластулы, а вегетативную – дном бластулы. Полное и равномерное дробление у ланцетника завершается всегда образованием бластулы, стенка к-й состоит из одного слоя клеток, а полость лежит в центре. Такая бластула называется целобластулой.
Полное неравномерное дробление наблюдается у земноводных, яйца к-х относятся к телолецитальным, со средним кол-вом желтка. Последний распределяется по всему яйцу, но большая часть его сосредоточена всегда в вегетативной половине, где и зерна его крупнее. В оплодотворенном яйце лягушки отчетливо выражена полярная дифференцировка, к-я проявляется в резкой пигментации анимальной половины. Первые две борозды дробления проходят меридионально. Они быстро разделяют анимальную часть, значительно задерживаясь в вегетативной. 3-я борозда, проходя параллельно экваториальной плоскости, разделяет бластомеры поперек. Она располагается ближе к анимальному полюсу. Это происходит потому, что веретено дробления смещается от от вегетативного полушария, где сосредоточена основная часть желтка, к анимальному. Верхние бластомеры получаются меньших размеров, чем нижние. При дальнейшем дроблении анимальные бластомеры дробятся заметно быстрее вегетативных, и поэтому разница между их размерами становится все более резкой.
Неполное дробление сопровождается делением только той части яйца, к-я свободна от желтка; часть, заполненная им, не дробится. По этому типу развиваются телолецитальные яйца (у костистых рыб, птиц, пресмыкающихся) и центролецитальные (у насекомых). Соответственно особенностям строения этих яиц различают дискоидальное и поверхностное дробление.
Дискоидальное дробление наблюдается у костистых рыб, птиц, пресмыкающихся. Яйца этих животных богаты желтком, большое кол-во к-го обусловливает их значительные размеры. Свободная от него ЦП, в виде небольшого участка находится в верхней части яйца и называется зародышевым диском. Т.к. дробится только последний, дробление получило название дискоидального.
Толщина зародышевого диска очень незначительна, он распластан на желтке, и его наибольшая протяженность совпадает с плоскостью, параллельной поверхности яйца. Поэтому веретена при первых 3-х и даже 4-х дроблениях располагаются горизонтально, а борозды дробления проходят вертикально. В результате получается 1 ряд клеток. После нескольких делений они становятся высокими и веретена располагаются в них вертикально, а борозды дробления – параллельно поверхности яйца. В связи с этим в зародышевом диске появляются поверхностные клетки и клетки, расположенные на желтке. Последующие деления проходят в самых различных направлениях, и зародышевый диск превращается в пластинку, состоящую из нескольких рядов клеток. Между диском и желтком возникает небольшая щелевидная полость, к-ю приравнивают к бластоцелю.
Поверхностное дробление наблюдается в центролецитальных яйцах с большим кол-вом желтка в их середине. ЦП в таких яйцах располагается по периферии клетки в центре, около ядра. Через массу желтка проходят тонкие ЦП-кие тяжи, соединяющие периферическую ЦП с околоядерной. Дробление начинается с деления ядра и обособления вокруг возникших ядер ЦП. Число ядер увеличивается. Окруженные ЦП, они постепенно передвигаются к периферии яйцеклетки. Как только ядра попадают в ее поверхностный слой, последний разделяется соответственно их кол-ву на бластомеры. Т.о., в результате дробления вся центр. часть ЦП перемещается к поверхности и сливается с периферической. Образуется сплошная бластодерма, из к-й развивается зародыш. Поверхностное дробление свойственно почти исключительно яйцам членистоногих.

Дробле́ние — ряд последовательных митотических делений оплодотворенного или инициированного к развитию яйца. Дробление представляет собой первый период эмбрионального развития, который присутствует в онтогенезе всех многоклеточных животных. При этом масса зародыша и его объём не меняются, оставаясь такими же, как и в начале дробления. Яйцо разделяется на все более мелкие клетки — бластомеры. Характерная особенность дробления — ведущая регуляторная роль цитоплазмы в развитии. Характер дробления зависит от количества желтка и его расположения в яйце.
Содержание
Биологическое значение дробления
Характерные черты дробления
Дробление как особый этап онтогенеза животных имеет характерные черты, которые свойственны большинству животных, но могут отсутствовать у некоторых групп.
- Бластомеры делятся очень быстро (у дрозофилы — раз в 20 минут) и более или менее синхронно. сокращена до S-периода; в связи с этим транскрипция собственных генов зародыша полностью подавлена, транскрибируются только запасённые в яйцеклетке материнские мРНК.
- Между делениями нет периода роста, так что общая масса зародыша не растёт.
По всем этим характеристикам дробление млекопитающих резко отклоняется от типичного. Бластомеры делятся у них медленно, синхронность нарушается уже после 1—2 делений, в это же время активируется собственный геном зародыша.
Классификация типов дробления
На основе ряда существенных характеристик (степень детерминированности, полнота, равномерность и симметрия деления) выделяют ряд типов дробления. Типы дробления во многом определяются распределением веществ (в том числе, желтка) по цитоплазме яйца и характером межклеточных контактов, которые устанавливаются между бластомерами.
Дробление может быть: детерминированным и регулятивным; полным (голобластическим) или неполным (меробластическим); равномерным (бластомеры более-менее одинаковы по величине) и неравномерным (бластомеры не одинаковы по величине, выделяются две — три размерные группы, обычно называемые макро- и микромерами); наконец, по характеру симметрии различают радиальное, спиральное, различные варианты билатеризованных и анархическое дробление. В каждом из этих типов выделяют ряд вариантов.
По степени детерминированности
Детерминированное
Недетерминированное (регулятивное)
По степени полноты делений
Голобластическое дробление
Плоскости дробления разделяют яйцо полностью. Выделяют полное равномерное дробление, при котором бластомеры не различаются по размерам (такой тип дробления характерен для гомолецитальных и алецитальных яиц), и полное неравномерное дробление, при котором бластомеры могут существенно различаться по размерам. Такой тип дробления характерен для умеренно телолецитальных яиц.
Меробластическое дробление
- Дискоидальное
- ограничено относительно небольшим участком у анимального полюса,
- плоскости дробления не проходят через всё яйцо и не захватывают желток.
Такой тип дробления типичен для телолецитальных яиц, богатых желтком (птицы, рептилии). Такое дробление называют также дискоидальным, так как в результате дробления на анимальном полюсе образуется небольшой диск клеток (бластодиск).
- Поверхностное
- ядро зиготы делится в центральном островке цитоплазмы,
- получающиеся ядра перемещаются на поверхность яйца, образуя поверхностный слой ядер (синцитиальную бластодерму) вокруг лежащего в центре желтка. Затем ядра разделяются мембранами, и бластодерма становится клеточной.
Такой тип дробления наблюдается у членистоногих.
По типу симметрии дробящегося яйца
Радиальное
Ось яйца является осью радиальной симметрии. Типично для ланцетника, осетровых, амфибий, иглокожих, круглоротых.
Хотя дробление амфибий часто описывается как радиальное, на самом деле еще до дробления, сразу же после оплодотворения, их яйцо приобретает билатеральную симметрию - на будущей спинной стороне зародыша образуется "серый серп" (его материал позднее попадает в клетки шпемановского организатора, или дорзальной губы бластопора). Билатеральную симметрию имеет и бластула амфибий.
Спиральное
В анафазе бластомеры разворачиваются. Отличается лево-правой дисимметрией (энантиоморфизм) уже на стадии четырёх (иногда двух) бластомеров. Типично для некоторых моллюсков, кольчатых и ресничных червей.
Билатеральное
Имеется 1 плоскость симметрии. Типично для аскариды.
Анархическое
Бластомеры слабо связаны между собой, сначала образуют цепочки или бесформенную массу; часто у одного вида встречаются разные варианты расположения бластомеров. Типично для кишечнополостных.
Литература
Wikimedia Foundation . 2010 .
Полезное
Смотреть что такое "Дробление (эмбриология)" в других словарях:
Дробление — Дробление: Дробление (технология) измельчение твёрдого тела до определенного размера; Дробление (полиграфия) воспроизведение на оттиске одного и того же печатающего элемента дважды, со смещением; Дробление (эмбриология) ряд… … Википедия
Дробление (значения) — Дробление: Дробление (технология) измельчение твёрдого тела до определенного размера; Дробление (полиграфия) воспроизведение на оттиске одного и того же печатающего элемента дважды, со смещением; Дробление (эмбриология) ряд последовательных… … Википедия
Дробление яйца — Дробление: эмбрион млекопитающего. z.p. zona striata, p.gl полярные тельца, a. двуклеточная стадия, b. четырёхклеточная стадия, c. восьмиклеточная стадия, d, e. морула Дробление ряд последовательных митотических делений оплодотворенного… … Википедия
дробление радиальное — ЭМБРИОЛОГИЯ ЖИВОТНЫХ ДРОБЛЕНИЕ РАДИАЛЬНОЕ – первое дробление меридиональное, проходит в меридиональной плоскости яйца. Второе дробление также меридиональное, оно проходит через главную ось яйца, но под прямым углом к плоскости первого дробления.… … Общая эмбриология: Терминологический словарь
дробление анархическое — ЭМБРИОЛОГИЯ ЖИВОТНЫХ ДРОБЛЕНИЕ АНАРХИЧЕСКОЕ [БЕСПОРЯДОЧНОЕ, ХАОТИЧНОЕ] – дробление яиц метагенетических медуз – Oceania armata. Первая борозда дробления меридиональная, врезающаяся, появляется она на анимальном полюсе. Вторая борозда тоже… … Общая эмбриология: Терминологический словарь
дробление гетероквадратное — ЭМБРИОЛОГИЯ ЖИВОТНЫХ ДРОБЛЕНИЕ ГЕТЕРОКВАДРАТНОЕ – дробление аннелид, моллюсков, немертин, планарий. Неравномерное спиральное дробление, когда клетки основного квартета (первые четыре бластомера) неодинаковы по величине, то и их производные также… … Общая эмбриология: Терминологический словарь
дробление асинхронное — ЭМБРИОЛОГИЯ ЖИВОТНЫХ ДРОБЛЕНИЕ АСИНХРОННОЕ – дробление яиц телолецитальных (амфибия). Деление вегетативных бластомеров происходит медленней, по сравнению с бластомерами анимального полюса … Общая эмбриология: Терминологический словарь
дробление билатеральное — ЭМБРИОЛОГИЯ ЖИВОТНЫХ ДРОБЛЕНИЕ БИЛАТЕРАЛЬНОЕ [ДВУСТОРОННЕ СИММЕТРИЧНОЕ] – дробление яиц нематод, коловраток, асцидий. Характеризуется появлением билатеральной симметрии в расположении бластомеров уже на ранних этапах дробления. Каждый бластомер… … Общая эмбриология: Терминологический словарь
дробление голобластическое — ЭМБРИОЛОГИЯ ЖИВОТНЫХ ДРОБЛЕНИЕ ГОЛОБЛАСТИЧЕСКОЕ [ПОЛНОЕ] – дробление яиц алецитального (плоские черви), изолецитального типа (ланцетник) и некоторых яйцевых клеток телолецитального типа (амфибии). Дробятся все части зиготы. При дроблении вся… … Общая эмбриология: Терминологический словарь
Как Вы уже знаете, в результате оплодотворения яйцеклетки образуется зигота, способная к дальнейшему развитию. Деление зиготы называют дроблением. Дробление – это многократное деление зиготы после оплодотворения, в результате которого образуется многоклеточный зародыш.
Зигота делится очень быстро, клетки уменьшаются в размерах и не успевают расти. Поэтому зародыш не увеличивается в объеме. Клетки, образующиеся в результате дробления зиготы, называются бластомерами, а перетяжки, отделяющие их друг от друга, называются бороздами дробления.
По направлению различают следующие борозды дробления: меридиональные – это борозды, которые делят зиготу от анимального к вегетативному полюсу; экваториальная борозда разделяет зиготу по экватору; широтные борозды проходят параллельно экваториальной борозде; тангенциальные борозды проходят параллельно поверхности зиготы.
Экваториальная борозда всегда одна, а меридиональных, широтных и тангенциальных может быть много. Направление борозд дробления всегда определяется положением веретена деления.
Дробление всегда проходит по определенным правилам:
Первое правило отражает местоположение веретена дробления в бластомере, а именно:
– веретено дробления располагается в сторону наибольшей протяженности цитоплазмы, свободной от включений.
Второе правило отражает направление борозд дробления:
– борозды дробления проходят всегда перпендикулярно веретену деления.
Третье правило отражает скорость прохождения борозд дробления:
– скорость прохождения борозд дробления обратно пропорционально количеству желтка в яйцеклетке, т.е. в той части клетки, где желтка мало, борозды будут проходить с большей скоростью, а в той части, где желтка больше, скорость прохождения борозд дробления замедляется.
Дробление зависит от количества и местоположения желтка в яйцеклетке. При небольшом содержании желтка дробится вся зигота, при значительном количестве дробится только часть зиготы, свободная от желтка. В связи с этим яйцеклетки разделяют на голобластические (дробящиеся полностью) и меробластические (с частичным дроблением). Следовательно, дробление зависит от количества желтка и с учетом ряда признаков подразделяется: по полноте охвата процессом материала зиготы на полное и неполное; по отношению размеров образующихся бластомеров на равномерное и неравномерное и по согласованности делений бластомеров – синхронное и асинхронное.
Полное дробление может быть равномерным и неравномерным. Полное равномерное дробление характерно для яйцеклеток с небольшим количеством желтка и его более или менее равномерным расположением в оплодотворённой яйцеклетке. Таким типом дробится яйцеклетка ланцетника. В этом случае первая борозда проходит от анимального к вегетативному полюсу, образуется два бластомера; вторая борозда тоже меридиональная, но проходит перпендикулярно первой, образуются четыре бластомера. Третья – экваториальная, образуются восемь бластомеров. После этого идет чередование меридиональных и широтных борозд дробления. Количество бластомеров после каждого деления увеличивается кратно двум (2; 4; 16; 32 и т.д.). В результате такого дробления образуется шарообразный зародыш, который называется бластулой. Клетки, которые образуют стенку бластулы, называют бластодермой, а полость внутри бластоцелью. Анимальная часть бластулы называется – крышей, а вегетативная часть – дном бластулы.

Полное неравномерное дробление характерно для яйцеклеток со средним содержанием желтка, расположенным в вегетативной части. Такие яйцеклетки характерны для круглоротых и земноводных. При этом типе дробления образуются бластомеры неодинаковых размеров. В анимальном полюсе образуются мелкие бластомеры, которые называются микромерами, а в вегетативном – крупные – макромеры. Первые две борозды, как и у ланцетника, проходят меридионально; третья борозда соответствует экваториальной борозде, но сдвинута от экватора к анимальному полюсу. Поскольку в анимальном полюсе находится свободная от желтка цитоплазма, то здесь дробление происходит быстрее и образуются мелкие бластомеры. В вегетативном полюсе содержится основная масса желтка, поэтому борозды дробления проходят медленнее и образуются крупные бластомеры.
Неполное дробление характерно для телолецитальных и центролецитальных яйцеклеток. В дроблении принимает участие только часть яйца, свободная от желтка. Неполное дробление делится на дискоидальное (костистые рыбы, пресмыкающиеся, птицы) и поверхностное (членистоногие).
Неполным дискоидальным дроблением делятся телолецитальные яйцеклетки, у которых большое количество желтка сконцентрировано в вегетативной части. У этих яйцеклеток безжелтковая часть цитоплазмы в виде зародышевого диска распластана на желтке в анимальном полюсе. Дробление происходит только в области зародышевого диска. Вегетативная часть яйцеклетки, заполненная желтком, участия в дроблении не принимает. Толщина зародышевого диска незначительна, поэтому веретена дробления при первых четырех делениях располагаются горизонтально, а борозды дробления проходят вертикально. Образуется один ряд клеток. После нескольких делений клетки становятся высокими и веретена дробления располагаются в них в вертикальном направлении, а борозды дробления проходят параллельно поверхности яйца. В результате зародышевый диск превращается в пластинку, состоящую из нескольких рядов клеток. Между зародышевым диском и желтком возникает небольшая полость в виде щели, которая аналогична бластоцели.
Неполное поверхностное дробление наблюдается в центролецитальных яйцеклетках с большим количеством желтка в его середине. Цитоплазма в таких яйцеклетках располагается по периферии и незначительная ее часть в центре около ядра. Вся остальная часть клетки заполнена желтком. Через массу желтка проходят тонкие цитоплазматические тяжи, соединяющие периферическую цитоплазму с околоядерной. Дробление начинается с деления ядер, в результате количество ядер увеличивается. Они окружаются тонким ободком цитоплазмы, передвигаются к периферии и располагаются в свободной от желтка цитоплазме. Как только ядра попадают в поверхностный слой, он делится соответственно их количеству на бластомеры. В результате такого дробления вся центральная часть цитоплазмы перемещается к поверхности и сливается с периферической. Снаружи образуется сплошная бластодерма, из которой развивается зародыш, а внутри находится желток. Поверхностное дробление свойственно яйцеклеткам членистоногих.
На характер дробления оказывают влияние и свойства цитоплазмы, которые определяют взаимное расположение бластомеров. По этому признаку выделяют радиальное, спиральное и билатеральное дробление. При радиальном дроблении каждый верхний бластомер располагается точно под нижним (кишечнополостные, иглокожие, ланцетник и др.). При спиральном дроблении каждый верхний бластомер смещен относительно нижнего наполовину, т.е. каждый верхний бластомер располагается между двумя нижними. В этом случае бластомеры располагаются как бы по спирали (черви, моллюски). При билатеральном дроблении через зиготу можно провести только одну плоскость, по обеим сторонам которой будут наблюдаться одинаковые бластомеры (круглые черви, асцидии).
От момента образования зиготы и до выхода зародыша из яйцевых оболочек длится эмбриональный период развития.
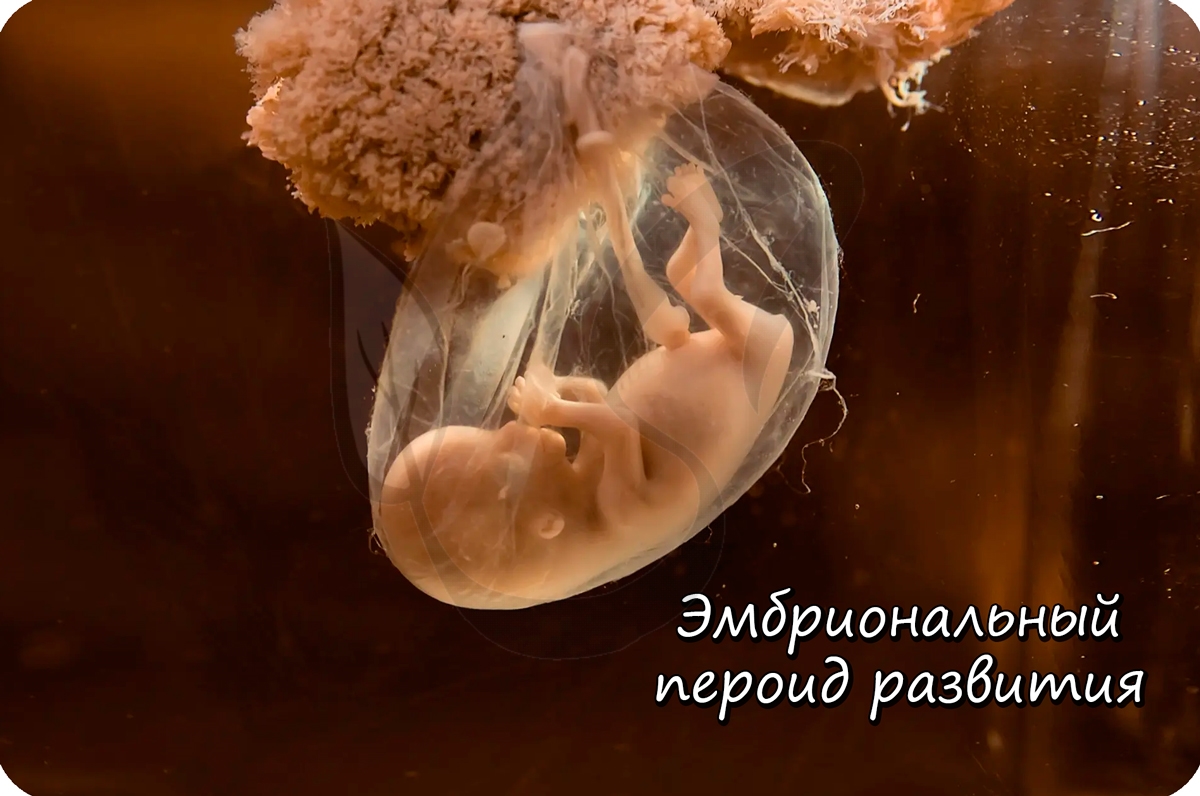
Дробление зиготы
После того, как произошло оплодотворение - слияние сперматозоида и яйцеклетки, образовавшаяся зигота начинает интенсивно делиться. Ее множественные митотические деления называют дроблением.
Важная особенность дробления в том, что не происходит увеличение в размере зародыша: клетки дробятся (делятся) настолько быстро, что не успевают накопить цитоплазматическую массу. Дробление зиготы человека является полным неравномерным асинхронным.
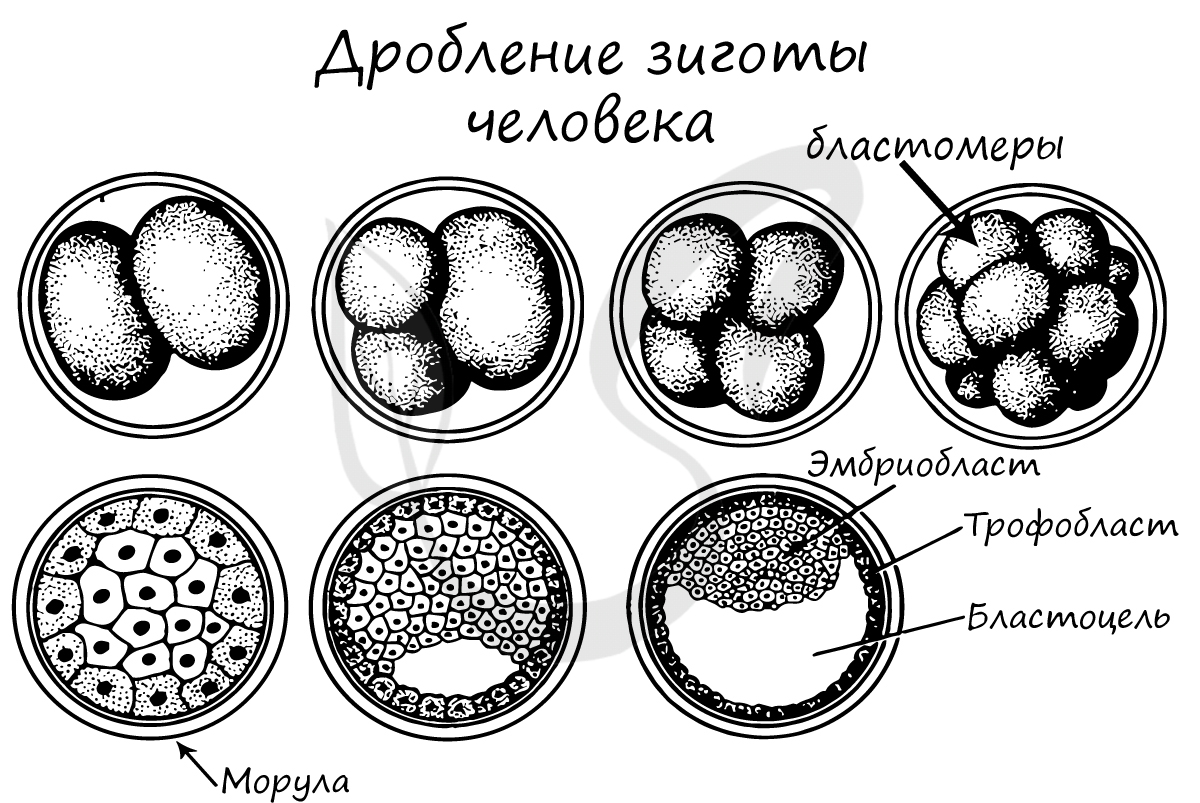
В результате дробления образуется морула. Морула (лат. morum - ягода тутового дерева) - клетка на стадии этапа дробления, когда зародыш представляет собой компактную совокупность клеток (без полости внутри).
Бластуляция
Бластуляция - заключительный период дробления, в который зародыш называется бластулой.
После очередных этапов многократного деления образуется однослойный зародыш с полостью внутри - бластула (греч. blastos — зачаток).
Стенки бластулы состоят из бластомеров, которые окружают центральную полость - бластоцель (греч. koilos — полый). Соединяясь друг с другом, бластомеры образуют бластодерму из одного слоя клеток.

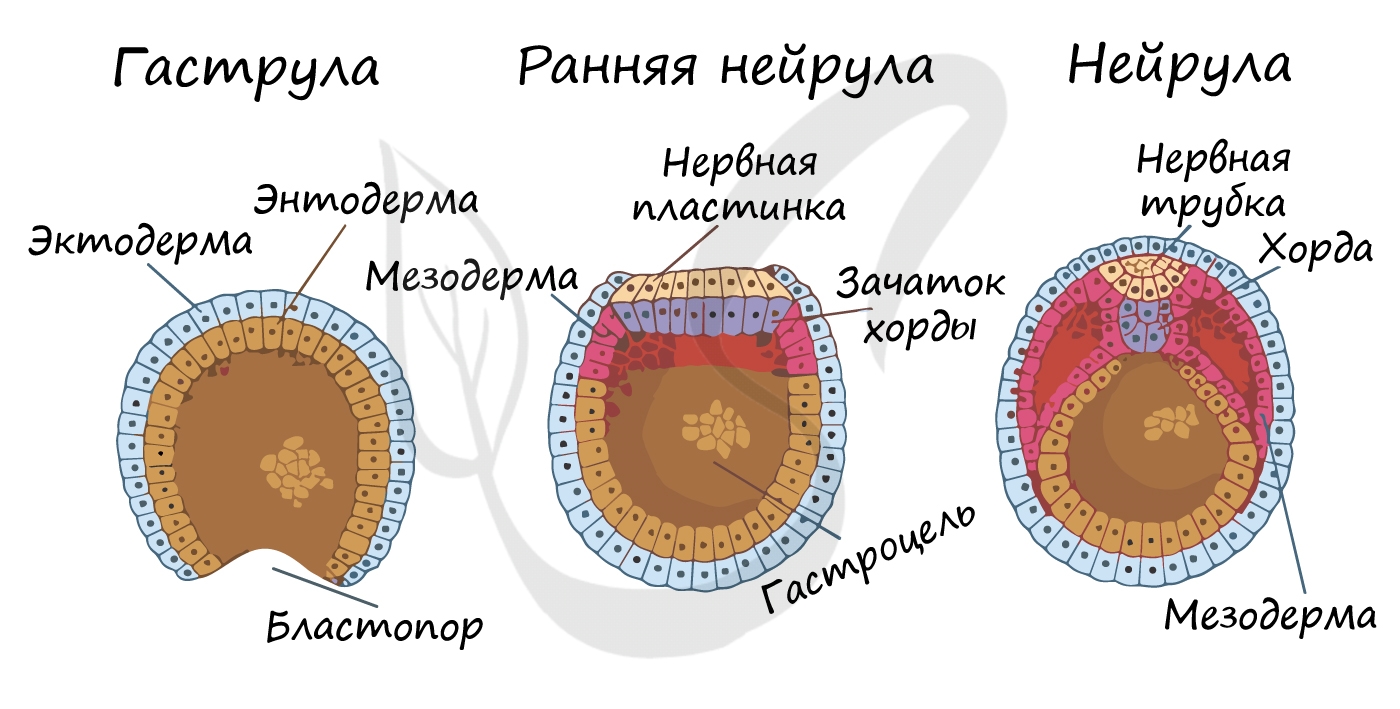
Гаструляция (греч. gaster — желудок, чрево)
Гаструляцией называют стадию эмбрионального развития, в ходе которой клетки, возникшие в результате дробления зиготы, формируют три зародышевых листка: эктодерму, мезодерму и энтодерму.
Стенка бластулы начинается впячиваться внутрь - происходит инвагинация стенки. По итогу такого впячивания зародыш становится двухслойным. Двухслойный зародыш называется - гаструла. Полость гаструлы называется гастроцель (полость первичной кишки), а отверстие, соединяющее гастроцель и внешнюю среду - первичный рот (бластопор).

У первичноротых животных на месте первичного рта (бластопора) образуется ротовое отверстие. К первичноротым относятся: кишечнополостные, плоские, круглые и кольчатые черви, моллюски, членистоногие.
У вторичноротых на месте бластопора формируется анальное отверстие, а ротовое отверстие образуется на противоположном полюсе. К вторичноротым относят хордовых и иглокожих (морских звезд, морских ежей).

При впячивании части бластулы (инвагинации) клетки бластодермы мигрируют внутрь и становятся энтодермой (греч. entós — внутренний). Оставшаяся часть бластодермы снаружи называется эктодермой (греч. ἔκτος - наружный).
Между энто- и эктодермой из группы клеток формируется третий зародышевый листок - мезодерма (греч. μέσος — средний).

Нейрула
Эта стадия следует за гаструлой. Ранняя нейрула представляет собой трехслойный зародыш, состоящий из энто-, экто- и мезодермы. На этапе нейрулы происходит закладка отдельных органов.
Важно отметить, что на стадии нейрулы происходит процесс нейруляции - закладывание нервной трубки. Нервная пластинка, образовавшаяся на ранних этапах, прогибается внутрь, при этом ее края сближаются и, замыкаясь, формируют нервную трубку.

Итак, как уже было сказано, на стадии нейрулы закладываются отдельные органы. Эктодерма образует покровный эпителий и нервную пластинку, мезодерма (из которой в дальнейшем появятся все соединительные ткани), энтодерма - окружает полость первичной кишки (гастроцель), образуя кишечник. От энтодермы отшнуровывается хорда.
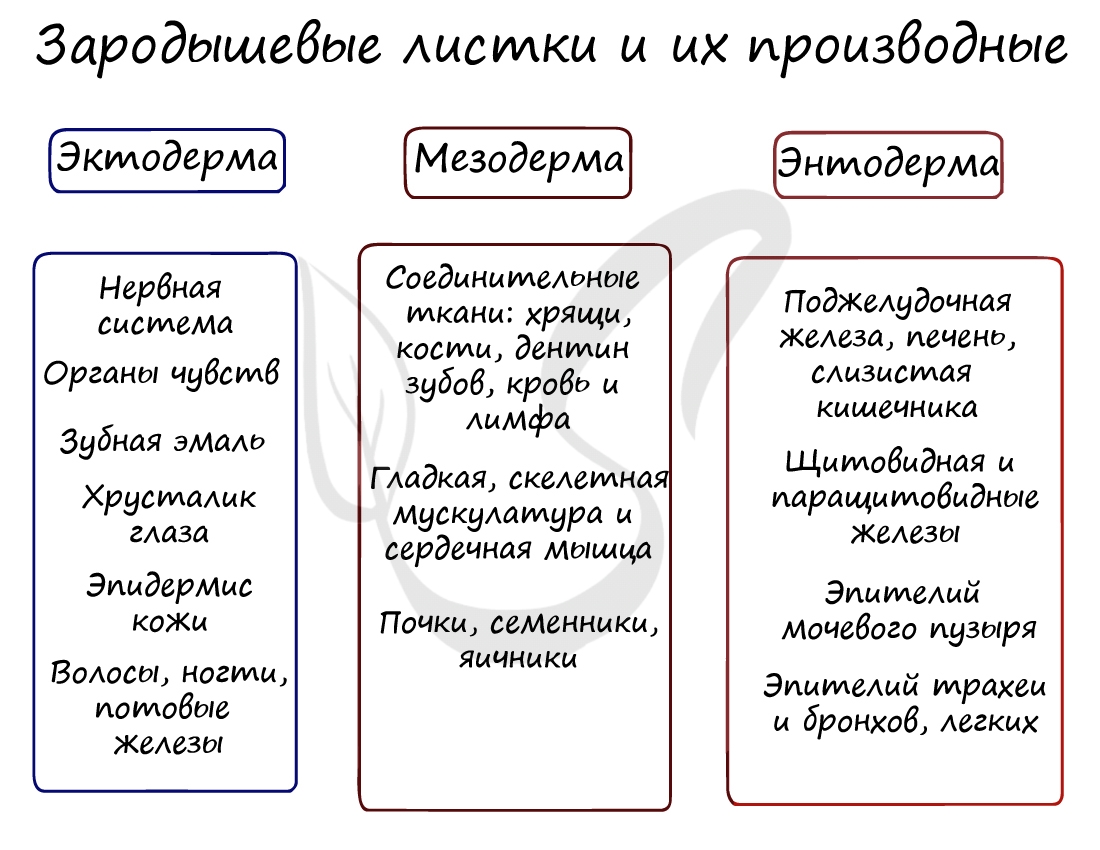
Все три зародышевых листка требуют нашего особого внимания, а также понимания того, какие органы и структуры из них образуются.
Эктодерма (греч. ἔκτος - наружный) - наружный зародышевый листок, образует головной и спинной мозг, органы чувств, периферические нервы, эпителий кожи, эмаль зубов, эпителий ротовой полости, эпителий промежуточного и анального отделов прямой кишки, гипофиз, гипоталамус.
Мезодерма (греч. μέσος — средний) - средний зародышевый листок, образует соединительные ткани: кровеносную и лимфатическую системы, костную и хрящевую ткань, мышечные ткани, дентин и цемент зубов, а также выделительную (почки) и половую системы (семенники, яичники).
Из зародышевых листков образуются ткани, органы и системы органов. Такой процесс называется органогенезом. В период закладки органов важное значение имеет воздержание матери от вредных привычек (алкоголь, курение), которые могут нарушить процесс дифференцировки клеток и привести к тяжелейшим аномалиям, уродствам плода.
Некоторые лекарства также могут оказывать на плод тератогенный эффект (греч. τέρας — чудовище, урод), приводя к развитию уродств. Периоды закладки органов и система органов вследствие их большой важности носят название критических периодов эмбриогенеза.
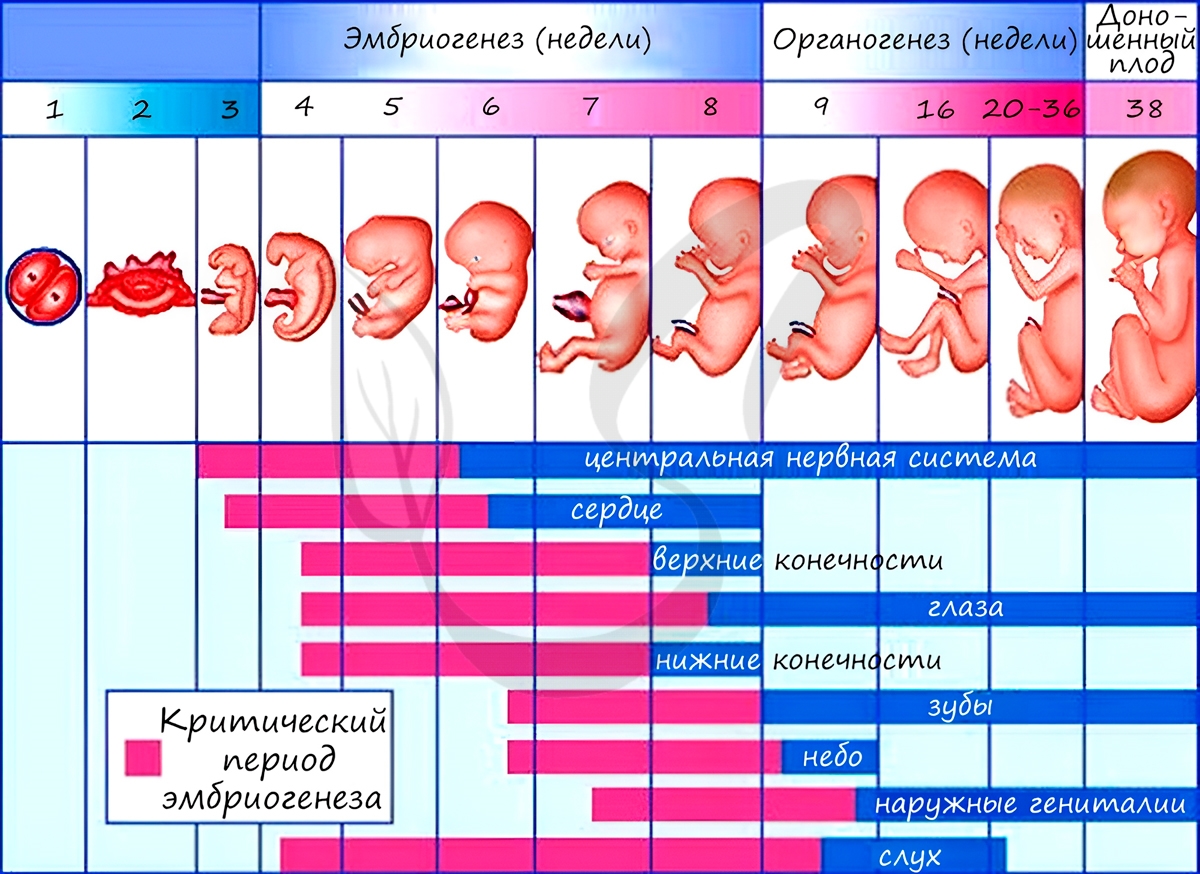
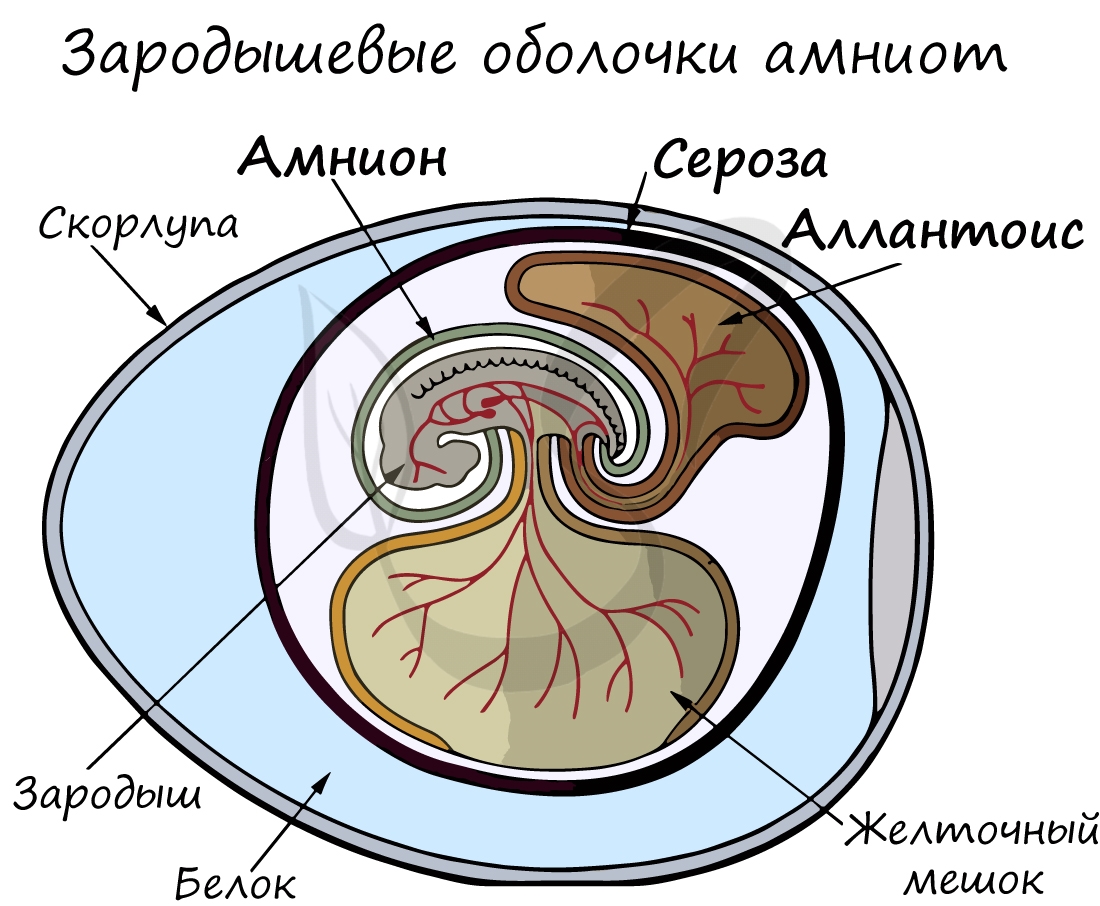
Анамнии и амниоты
Анамнии, или низшие позвоночные - группа животных, не имеющая зародышевых оболочек (зародышевого органа - аллантоиса и амниона). Анамнии проводят большую часть жизни в воде, без которой невозможно их размножение.
К анамниям относятся рыбы, земноводные.

Амниоты - группа высших позвоночных, характеризующаяся наличием зародышевых оболочек. К амниотам относятся пресмыкающиеся, птицы и млекопитающие.
Зародышевый орган, аллантоис, является органом дыхания и выделения.
За счет особых оболочек, развивающихся в ходе эмбрионального развития, амниона и серозы, у амниот формируется амниотическая полость. В ней находится зародыш, окруженный околоплодными водами. Благодаря такому гениальному устройству, амниотам для размножения и развития более не нужно постоянное нахождение в водоеме, они "обрели независимость" от него.
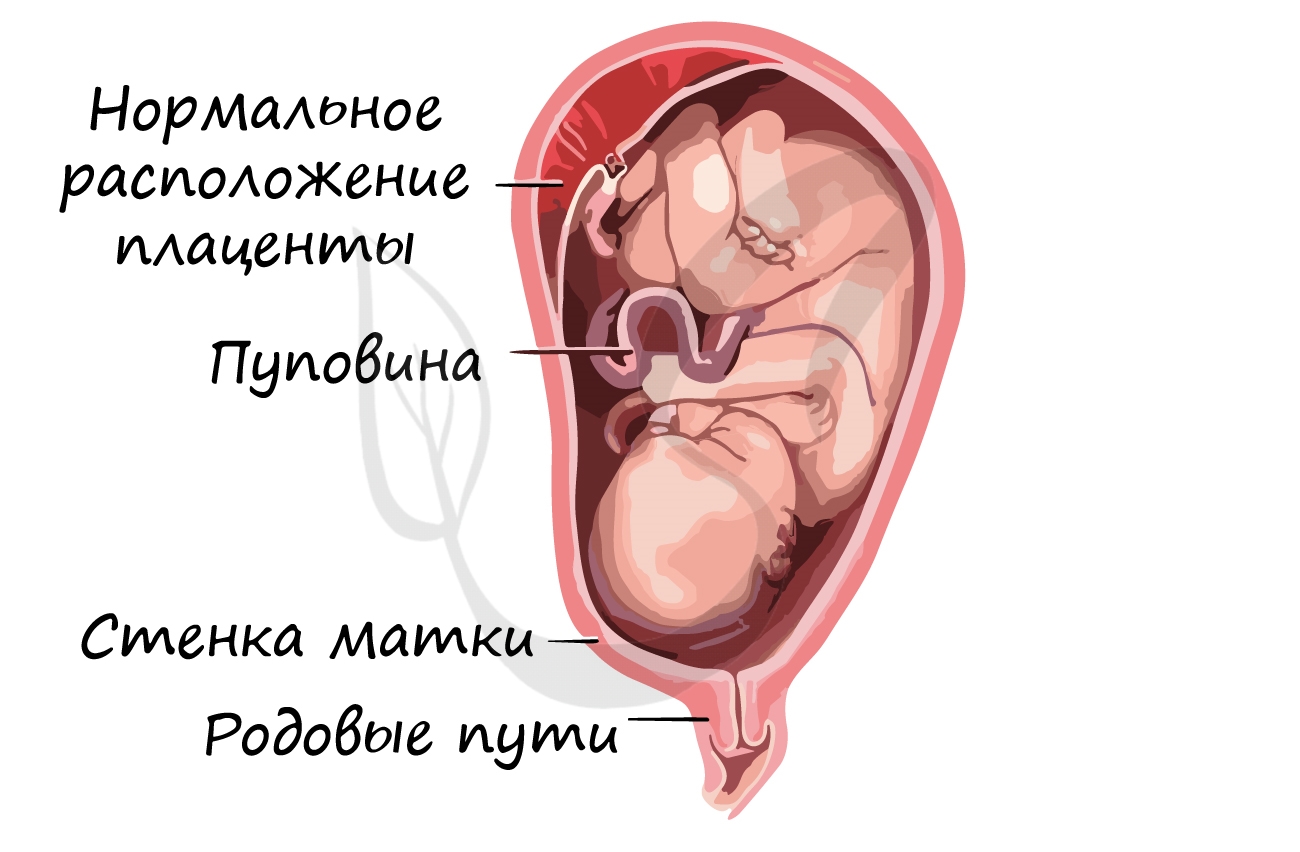
Развитие плода происходит в мышечном органе - матке, которая, сокращаясь во время родов, стимулирует изгнание плода через родовые пути. Питание осуществляется через плаценту - "детское место" - орган, который с одной стороны омывается кровью матери, а с другой - кровью плода. Через плаценту происходит транспорт питательных веществ и газообмен.
Соединяет плаценту и плод особый орган - пуповина, внутри которой проходят артерии, вены.
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Читайте также: