
Мутации у бактерий кратко
Обновлено: 08.07.2024
Мутации - стабильное изменение первичной структуры геномной ДНК , которое приводит к наследственно закрепленному изменению или утрате одного или нескольких признаков, не связанное с генетическим обменом или присутствием плазмид.
Спонтанные мутации возникают в обычных условиях под влиянием природных факторов,ошибок в репликации генома, действия транспозонов и Is-элементов.
Индуцированные мутации получают с помощью обработки бактериальной культуры мутагенами - факторами химической, физической или биологической природы, способными вызывать повреждения ДНК .
В зависимости от степени мутационных повреждений мутации подразделяют на :
Хромосомные - крупные перестройки в отдельных фрагментах ДНК. (Делеции - выпадение нескольких нуклеотидов , инверсии - поворот участка хромосомы на 180*, дупликации - повторение участка хромосомы, дислокации - изменение последовательности нескольких оснований). Бывают летальными, обычно приводящими мутировавшие бактерии к гибели.
Генные (точковые) мутации - изменение одной пары оснований. Различают : выпадение или вставки оснований, простые замены (транзиции) - при которых пурин заменяется на другой пурин( аденин>гуанин) , пиримидин на другой пиримидин ( тимин>цитозин), сложные замены (трансверии) , когда пурины заменяются на пиримидины(а>т , ц>г). Точковые мутации возникают с большой частотой и имеют большое значение в Генотипической изменчивости бактерий, способствуя их эволюции.
Прямые мутации - связаны с потерей функцией.Обратные мутации - с восстановлением этой функции
№8 Генетические рекомбинации у бактерий. Способы переноса генетической информации из клетки в клетку. Трансформация: сущность, условия возникновения, этапы, значение.
Рекомбинации- это процесс взаимодействия между двумя молекулами ДНК, приводящий к образованию новой рекомбинантной молекулы.
Типы генетических рекомбинаций у бактерий: трансформация, трансдукция, конъюгация.
Трансформация- тип генетического обмена, при котором участок нативной молекулы ДНК донора проникает в бактерию-реципиент и вызывает изменение ее генотипа. Для того, чтобы трансформация была успешной, ДНК донора и бактерии-реципиента должны обладать определёнными свойствами:
- ДНК донора должна быть выделена из бактериальной культуры того же вида, что и реципиент (или близкородственного). Участок трансформирующей ДНК должен сохранять интактную структуру (двунитчатую спирализацию) и иметь значительную молекулярную массу.
- Клетки реципиентной бактериальной культуры для осуществления трансформации должны быть компетентными, то есть способными адсорбировать на своей поверхности ДНК донора и поглащать ее
Адсорбция фрагментов двунитивой ДНК донора на поверхности клеточной стенки компетентных клеток. ДНК связывается с участками поверхности, доступными только у компетентных клеток.
Ферментативное расщепление адсорбированной молекулы двунитивой ДНК на более мелкие фрагменты с помощью индуцированной клеточной эндонуклеазы.
Проникновение фрагментов ДНК сопровождается разрушением одной из нитей ДНК и освобождением энергии, необходимой для дальнейшего проникновения.
Проникший однонитевый фрагмент ДНК взаимодействует с участком высокой гомологии генома клетки реципиента с образованием гибридной молекулы ДНК (гомологичная рекомбинация), в которой одна нить содержит генотип реципиента, а вторая нить является рекомбинантной.
Трансформация является одним из факторов эволюции бактерий. Появляющиеся рекомбинанты с новым сочетанием генов играют определённую роль в естественном отборе. С помощью трансформации проводится тонкое картирование генов, уточняются генетические карты.
№9 Генетический обмен у бактерий. Конъюгация: сущность, этапы, значение. Типы штаммов-доноров: F+; Hfr; F`: свойства донорных клеток и особенности взаимодействия с реципиентом.
Конъюгация- форма обмена генетическим материалом между бактериями при их непосредственном клеточном контакте. Этот процесс контролируется F-плазмидами, а также другими конъюгативными плазмидами. При конъюгации донорами являются “мужские” клетки, содержащие F-плазмиды, а реципиентами являются “женские” F-минус клетки, у которых плазмида отсутствует.
Бактериальные штаммы, в клетках которых F-фактор интегрирован в хромосомы носят название Hfr-штаммы.
F`-свободная плазмида,несущая несколько бактериальных генов.
Типы скрещиваний: F+ x F-; Hfr x F-;F` x F-
Клетки-доноры возможно отличить от реципиентных клеток по наличию F-пилей, дополнительного поверхностного антигена, по чувствительности к некоторым РНК-бактериофагам, а также по ряду физико-химических свойств.
№10 Генетические рекомбинации у бактерий. Типы рекомбинации по молекулярному механизму. Способы переноса генетического материала из клетки в клетку. Трансдукция: сущность, типы трансдукции, значение.
Генетические рекомбинации по молекулярному механизму делятся на гомологичную, сайт-специфическую и незаконную рекомбинацию.
Трансдукция- процесс переноса генетического материала от бактерии-донора к бактерии-рецепиенту с помощью бактериофага. Трансдукция осуществляется дефектными бактериофагами, которые не способны к самостоятельной репродукции в клетке и для формирования фагового потомства нуждаются в присутствии родственного фага.
Типы трансдукции: неспецифическая (общая) и специфическая (локализованная) и абортивная.
Спомощью трансдукции происходит перенос бактериальных генов, плазмид и транспозонов, что используется в генной инженерии.
№11 Микробиологические основы генной инженерии и биотехнологии- клонирующий вектор, эндонуклеазы рестрикции, ДНК-лигазы, клонирование ДНК. Применение генной инженерии в микробиологической биотехнологии: создание диагностических, лечебных и профилактических препаратов.
Генная инженерия- раздел молекулярной генетики, представляющий собой совокупность методов и технологий для создания in vitro рекомбинантных молекул ДНК или РНК и способов их переноса. В результате клетки, получившие чужеродные гены, приобретают способность синтезировать белки, которые они ранее не синтезировали.
Рекомбинантные молекулы содержат два компонента- вектор(переносчик) и клонируемую “чужеродную” ДНК.
Векторами являются циркулярно замкнутные молекулы ДНК- плазмиды, умеренные бактериофаги или вирусы животных, иногда космиды( плазмиды с липкими концами, cos-сайтами ДНК бактериофага лямбда). Они должны обладать рядом свойств, в частности:
- нести в своем составе участки рестрикции
- содержать маркеры фенотипа
Клонируемые молекулы ДНК- это фрагменты ДНК, имеющие определённые гены, кодирующие синтез нужного вещества. Их получают экстракцией натриевой ДНК, с помощью фермента обратной транскриптазы на молекуле и-РНК и синтезов генов химическим способом.
Рекомбинантную молекулу ДНК можно получить с помощью фермента рестриктазы. Обрабатываются ДНК вектора и изолированные ДНК эндонуклеазой рестрикции,которая расщепляет взятые молекулы ДНК в строго определенном месте с образованием однонитчатых, комплементарных друг друг концов. Далее фермент полинуклеотидлигаза сшивает две разные линейные молекулы ДНК в одну рекомбинантную ДНК.
Генная инженерия применяется для переноса генной информации из одной клетки в другую,преодолевая даже межвидовые барьеры; для создания микроорганизмов-продуцентов ферментов, гормонов, вакцин, антигенных препаратов и т.д.
№12 Молекулярно-генетические методы диагностики. Амплификационные технологии: конвенциональная ПЦР, ПЦР в реальном времени. Некоторые разновидности ПЦР: ПЦР с обратной транскрипцией, мультиплексная ПЦР и др. Области применения генетических технологий.
Молекулярно-генетические методы- методы, позволяющие в генетических материалах обнаруживать фрагменты геномов.
ПЦР- метод получения большого количества специфических нуклеотидных последовательностей ДНК in vitro. Амплификация-умножение определенных генов или их фрагментов с определенной нуклеотидной последовательностью ДНК. В результате реакции исследуемый генетический материал накапливается в значительном количестве и может быть легко выявлен и индентифицирован.
Денатурация- разъединение определяемой двухцепочечной ДНК на две изолированные цепочки при нагревании до 90-95 градусов по Цельсию в течении 0.5-1 минут.
Отжиг- восстановление двухцепочечной структуры определяемой ДНК в области присоединения комплементарного праймера при температуре 40-60 0.5 минут.
Удлинение (элонгация)- достройка каждой цепи определяемой ДНК до исходного двухцепочечного состояния с помощью термостабильной ДНК- полимеразы- проводится при температуре 70-75 в течение 2-5 минут.
Конвенциональная ПЦР
В данном варианте постановки ПЦР реакция идет заранее выбранное число циклов (30-40), после чего анализируется, произошло ли накопление двуцепочечных молекул ДНК в реакционной смеси.
ПЦР в реальном времени
В данном варианте постановки ПЦР количество продукта ПЦР в реакционной смеси регистрируется постоянно в ходе протекания реакции. Это позволяет построить кривую протекания реакции и, исходя из неё, рассчитать количество искомых молекул ДНК в образцах.
Цифровая количественная ПЦР
Новый, дорогостоящий и пока малораспространенный вариант ПЦР, позволяющий более точно определять количество ДНК в образце.В данном варианте реакционная смесь, содержащая флуоресцентный краситель, разбивается на огромное число микроскопических объемов (например, капелек в эмульсии). После протекания ПЦР анализируется, в какой доле капелек реакция оказалась положительной и, соответственно, наблюдается флуоресценция. Эта доля будет пропорциональна числу искомых молекул ДНК в образце.
ПЦР с обратной транскрипцией
В данном случае перед тем или иным вариантом ПЦР производится реакция обратной транскрипции (РНК в ДНК) с использованием фермента ревертазы. Таким образом, этот метод позволяет проводить качественное или количественное обнаружение молекул РНК. Это может использоваться для детекции РНК-содержащих вирусов или определения уровня транскрипции (количества мРНК) того или иного гена.
№13 Молекулярно-генетические методы диагностики. Гибридизационный анализ нуклеиновых кислот, секвенирование ДНК. Области применения генетических технологий.
Секвенирование биополимеров (белков и нуклеиновых кислот — ДНК и РНК) — определение их аминокислотной или нуклеотиднойпоследовательности (от лат. sequentum — последовательность). В результате секвенирования получают формальное описание первичной структуры линейной макромолекулы в виде последовательности мономеров в текстовом виде. Размеры секвенируемых участков ДНК обычно не превышают 100 пар нуклеотидов (next-generation sequencing) и 1000 пар нуклеотидов при секвенировании по Сенгеру. В результате секвенирования перекрывающихся участков ДНК, получают последовательности участков генов, целых генов, тотальной мРНК и даже полных геномов организмов [1][2] .
Для секвенирования применяют методы Эдмана, Сэнгера и другие; в настоящее время для секвенирования генов обычно применяют метод Сэнгера с дидезоксинуклеозидтрифосфатами (ddNTP). Обычно до начала секвенирования производят амплификацию участка ДНК, последовательность которого требуется определить, при помощи ПЦР.
Мутация. Мутации у бактерий. Мутагены. Спонтанные мутации. Обратные мутации ( реверсии ).
Они обычно имеют физическую или химическую природу. По происхождению выделяют мутации индуцированные, то есть вызванные искусственно, и спонтанные
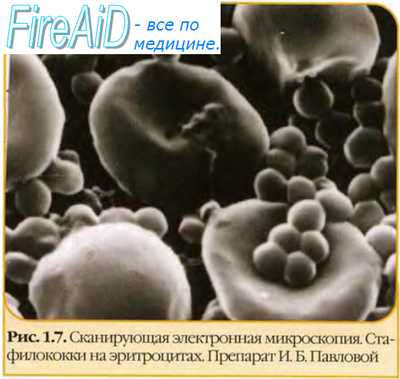
Спонтанные мутации. Обратные мутации ( реверсии ).
К появлению спонтанных мутаций приводят ошибки репликации, неправильное формирование комплементарных пар оснований или структурные искажения ДНК под действием естественный мутагенов. Спонтанные мутации могут вызывать благоприятные и неблагоприятные генетические изменения. Примерный уровень спонтанного мутирования — одна мутация на каждые 10 6 -10 7 клеток. Численная доля мутантов в клеточной популяции для разных признаков различна и может варьировать от 10 -4 до 10 -11 .
Для конкретного гена частота мутирования составляет величину порядка 10 -5 , а для определённой пары нуклеотидов 10 -8 . Например, если на среду с антибиотиком посеять миллион бактерий, можно ожидать, что в результате спонтанной мутации одна колония выживет.
• Обратные мутации (реверсии) возвращают спонтанно мутировавшую клетку к исходному генетическому состоянию. Их наблюдают с частотой одна клетка на 10 7 -10 8 (то есть по меньшей мере в 10 раз реже, чем прямые спонтанные мутации).
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
11. Мутации и рекомбинации у бактерий. Виды рекомбинаций: гомологичная, сайт-специфическая, незаконная (репликативная).
Мутации – это изменения в последовательности нуклеотидов ДНК, проявляющиеся наследственно закрепленной утратой или изменением какого-либо признака или группы признаков. В их основе лежат ошибки копирования наследственной информации, возникающие при репликации. Фенотипическим проявлением мутации могут быть: изменение морфологии бактериальной клетки, возникновение потребности в факторах роста (ауксотрофность), появление устойчивости к антибиотикам, изменение чувствительности к температуре, снижение вирулентности (аттенуация).
Рекомбинативаня изменчивость у бактерий:
Генетическая рекомбинация – взаимодействие между двумя геномами, которое приводит к образованию рекомбинаций ДНК и формированию дочернего генома, сочетающего гены обоих родителей.
Особенности рекомбинаций у бактерий определяются отсутствием истинного полового процесса и мейоза у прокариот и гаплоидным набором генов.
В процессе рекомбинации бактерии условно делятся на клетки-доноры, которые передают генетический материал, и клетки-реципиенты, которые этот материал воспринимают. В клетку-реципиент проникает не вся, а только часть хромосомы клетки-донора, т.е. один или несколько генов. Образуется только один рекомбинант, генотип которого представлен в основном генотипом реципиента с включением фрагментов хромосомы донора.
v Гомологичная – обмен между участками ДНК, обладающими высокой степенью гомологии, происходит в процессе разрыва и воссоединения ДНК через образование промежуточного соединения (крестообразной структуры Холидея или полухиазмы).
Процесс находится под контролем генов REC -системы (гены А, В, С и D ), продукты экспрессии которых производят расплетание нитей ДНК, их переориентацию с образованием полухиазмы, затем ее разрезание.
v Сайт-специфичная – происходит в определенных участках генома и не требует высокой степени гомологиии ДНК. Не зависит от генов REC -системы.
- встраивание плазмиды в хромосому бактерий (между IS элементами хромосомы и плазмиды);
- интеграция ДНК фага в хромосому бактерии.
v Незаконная (репликативная) – происходит на любом участке ДНК, не зависит от генов REC -системы.
Например, транспозиция подвижных генетических элементов по репликону или между репликонами.
Рекомбинация между геномами бактерий осуществляется 3-мя механизмами: конъюгацией, трансдукцией и трансформацией
Трансформация – передача генетической информации через выделенную из клетки-донора ДНК.
1928г. Ф Гриффит (опыт с вирулентнотью пневмококков)
По происхождению ДНК может быть плазмидной или хромосомной и нести гены, трансформирующие реципиента. Подобным путем среди бактериальных популяций могут распространять гены, кодирующие факторы вирулентности, однако в обмене генетической информацией трансформация играет незначительную роль.
Трансформирующей активностью обладает только двунитчатая высокоспирализованная ДНК.
В клетку-реципиент проникает только одна нить ДНК, другая – в клеточной мембране подвергается деградации с освобождением энергии, необходимой для проникновения в клетку второй нити ДНК.
Интеграция с хромосомой требует наличия гомологичных участков с трансформирующей ДНК.
Процесс трансформации зависит от компетентности клетки-реципиента и состояния трансформирующей донорской ДНК.
Компетентность клеток бактерий (способность воспринимать трансформирующую ДНК) зависит от присутствия в ЦПМ особых белков, обладающих специфическим аффинитетом к ДНК. Компетентность зависит от фазы роста бактериальной культуры, вида бактерий. Может увеличиваться при обработке культуры раствором хлорида кальция при пониженной температуре.
Трансформация служит хорошим инструментом для картирования хромосом, поскольку трансформированные клетки включают различные фрагменты ДНК. Определение частоты одновременного приобретения двух заданных характеристик (чем ближе расположены гены, тем более вероятно, что они оба включатся в один и тот же участок ДНК) дает информацию о взаиморасположении соответствующих генов в хромосоме.
Трансформация является основным методом генной инженерии, используемым при конструировании рекомбинантных штаммов с заданным геномом.
Трансдукция – передача бактериальной ДНК посредством бактериофага.
1952г. Дж. Ледебург и Н. Циндер (опыты с сальмонеллами)
Так как трансдуцируются лишь небольшие фрагменты ДНК, вероятность рекомбинации, затрагивающей какой-то определенный признак, очень мала: она составляет от 10 -6 до 10 -8 .
· Общая (неспецифическая) трансдукция – перенос бактериофагом фрагмента любой части бактериальной хромосомы. Феномен неспецифической трансдукции может быть использован для картирования бактериальной хромосомы.
● Специфическая трансдукция – фаговая ДНК интегрирует в бактерию с образованием профага. При исключении ДНК фага из бактериальной хромосомы захватывается прилегающий к месту включения фаговой ДНК фрагмент бактериальной хромосомы. Специфическая трансдукция может служить механизмом переноса вирулентных генов среди бактерий при условии, что эти гены локализованы в непосредственной близости от мест интеграции профага.
● Абортивная трансдукция . При абортивной трансдукции внесенный фрагмент ДНК донора не встраивается в хромосому реципиента, а остается в цитоплазме и там самостоятельно функционирует. Впоследствии он передается одной из дочерних клеток (т.е. наследуется однолинейно) и затем теряется в потомстве.
Обязательное условие – трансмиссивная плазмида ( F , R ), обладающая tra -опероном .
Интегративной трансмиссивной плазмидой является F -фактор . Донорские клетки, имеющие встроенный в хромосому F -фактор, называются Hfr -клетками.
Биологическое значение конъюгации хорошо видна на примере распространения резистентности бактерий к антибиотикам. Устойчивость к антибиотикам бактерия может получить в результате мутации, что происходит 1 раз на каждые 106 клеточных делений. Изменившаяся генетическая информация может быстро распространяться среди сходных бактерий посредством конъюгации.
Мутации —это изменения в последовательности отдельных нуклеотидов ДНК, которые ведут к таким проявлениям, как изменения морфологии бактериальной клетки, возникновение потребностей факторах роста, например в аминокислотах, витаминах, т. е. ауксотрофности; к устойчивости к антибиотикам, изменению чувствительности к температуре, снижению вирулентноти (аттенуация) и т. д.
1.По протяженности изменений повреждения ДНК различают мутации точечные, когда повреждения ограничиваются одной парой нуклеотидов, и протяженные или аберрации. (В последнем случае могут наблюдаться выпадения нескольких пар нуклеотидов, которые называются делецией, добавление нуклеотидных пар, т. е. дупликации,перемещения фрагментов хромосомы, транслокациии перестановки нуклеотидных пар — инверсии.)
2.Мутации могут быть спонтанными,т. е. возникающими самопроизвольно, без воздействия извне, и индуцированными.
Точечные спонтанные мутации возникают в результате возникновения ошибок при репликации ДНК, что связано с таутомерным перемещением электронов в азотистых основаниях.
Спонтанные хромосомные аберрации возникают вследствие перемещения подвижных генетических элементов.
Цель настоящей статьи - раскрытие сущности понятия мутагенеза, а также роли данного процесса в понимании эволюции всех живых организмов на нашей планете. Подробно изложены современные классификации мутагенеза, в том числе разбору подвергнуто понятие о генных мутациях. В материале работы изложены наиболее вероятные причины, которые могут потенцировать частоту возникновения и масштабность различных нарушений в хранении, воспроизведении и передаче генетической информации.

2. Ходжкин Ю. Генетическое подавление. 2005 г., 27 декабря. В: WormBook: Интернет-обзор биологии C. elegans. Пасадена (Калифорния): WormBook; 2005-2018.
5. Жимулёв, И.Ф. Общая и молекулярная генетика /И.Ф. Жимулёв. — Издание четвертое. — Новосибирск: Новосибирское университетское издательство, 2007. — 480с.
Введение. Мутагенез - внесение изменений в нуклеотидную последовательность ДНК (мутаций). (Гуго де Фриз 1901)
Виды мутагенеза. Различают естественный и искусственный мутагенез.
Естественный (спонтанный) - возникает вследствие УФ-лучей, химических мутагенов, радиации.
Искусственный (индуцированный) - искусственное получение мутаций путем воздействия радиационного излучения и химических веществ. Широко используется в селекции (полиплоидия).
Роль мутагенеза. Зачастую мутации выступают в качестве материала для естественного отбор. Например: при кардинальном изменении окружающих организм условий мутации, считавшиеся ранее ненужными, могут стать полезными, и повысят процент выживаемости данного организма и впоследствии его потомков.
Согласно одной из теорий происхождения жизни на нашей планете все живое произошло от одной клетки. В процессе эволюции эта клетка дифференцировалась с помощью мутаций. Так возникли мы и самое важное –
разные люди (цвет волос, глаз и т.д.) Стоит также отметить, что мутации играют большую роль в селекции. Путем искусственного мутагенеза получают более крупные плоды. Таким образом, благодаря мутациям возникают новые штаммы, сорта, породы организмов.
Мутации с нарушением генетического кода (генные мутации). Генные мутации – это изменение строения одного гена, т.е. изменение в последовательности нуклеотидов, а следовательно, изменение генетического кода и изменение молекулы белка, синтезируемого по этому коду. Если изменяется код, то изменяется и кодируемый им признак. Последствия генных мутаций могут быть разные – все зависит от гена, с которым произойдет спонтанное изменение. Если случится нарушение синтеза аминокислоты, необходимой для полноценного функционирования организма, то будут серьезные осложнения вплоть до смертельного исхода. Если действие мутировавшего гена будет подавлено парным геном из гомологичной хромосомы или если изменение в молекуле синтезируемого белка не будет нарушать его функций, то мутация никак не отразится на фенотипе.
Виды генных мутаций:
1. Дупликация – удвоение пары или нескольких пар нуклеотидов;
2. Инсерция – вставка пары нуклеотидов (или несколько);
3. Делеция – выпадение участка генома;
4. Инверсия – переворот на 180 градусов;
5. Замена – замена пары нуклеотидов на другую.
Так, например, замена глутаминовой кислоты на валин в молекуле глобина (белковой части гемоглобина) приводит к катастрофическим последствиям). Гемоглобин начинает хуже связывать и переносить кислород. Эритроциты, в которых содержится гемоглобин, становятся непрочными и легко разрушаются. Вследствие замены одной из ста сорока шести аминокислот на другую развивается тяжелое заболевание – серповидноклеточная анемия. Так назвали из-за формы гемоглобина – в форме серпа.
Читайте также: