
Матричные биосинтезы биохимия кратко
Обновлено: 04.07.2024
Основной фигурой матричных биосинтезов являются нуклеиновые кислоты РНК и ДНК. Они представляют собой полимерные молекулы, в состав которых входят азотистые основания пяти типов, пентозы двух типов и остатки фосфорной кислоты. Азотистые основания в нуклеиновых кислотах могут быть пуриновыми ( аденин , гуанин ) и пиримидиновыми ( цитозин , урацил (только в РНК), тимин (только в ДНК)). В зависимости от строения углевода выделяют рибонуклеиновые кислоты – содержат рибозу (РНК), и дезоксирибонуклеиновые кислоты – содержат дезоксирибозу (ДНК).
Термин " матричные биосинтезы " подразумевает способность клетки синтезировать полимерные молекулы, таких как нуклеиновые кислоты и белки, на основе шаблона – матрицы . Это обеспечивает точную передачу сложнейшей структуры от уже существующих молекул к новосинтезируемым.
Основной постулат молекулярной биологии
В подавляющем большинстве случаев передача наследственной информации от материнской клетки к дочерней осуществляется при помощи ДНК ( репликация ). Для использования генетической информации самой клеткой необходимы РНК, образуемые на матрице ДНК ( транскрипция ). Далее РНК непосредственно участвуют на всех этапах синтеза белковых молекул ( трансляция ), обеспечивающих структуру и деятельность клетки.
На вышесказанном основана центральная догма молекулярной биологии , согласно которой перенос генетической информации осуществляется только от нуклеиновой кислоты (ДНК и РНК). Получателем информации может быть другая нуклеиновая кислота (ДНК или РНК) и белок.

Лекция № 12 Биохимия нуклеиновых кислот. Матричные биосинтезы.

1. Структура и функции ДНК (закономерности нуклеотидного состава, структура, гистоны, укладка ДНК в хроматине).

Химическая организация гена А. Строение нуклеозида. Нуклеозиды состоят из азотистых оснований и углевода – пентоза. Название пуриновых нуклеозидов имеют окончание – ОЗИН, а пиримидиновых нуклеозидов окончание – ИДИН. Б. Строение нуклеотидов. Нуклеотиды являются мономерами нуклеиновых кислот они состоят из азотистых оснований углевода – пентоза и фосфорной кислоты. Нуклеотиды хорошо растворимы в воде и обладают кислотными свойствами → нуклеотиды являются кислотами. Разные нуклеотиды отличатся между собой природой углеводов и азотистых оснований.

В. Строение и виды азотистых оснований: Азотистые основания нуклеотидов делятся на 2 типа: 1. Пиримидиновые – они состоят из 1 шестичленного кольца; 2. Пуриновые - состоят из 2 конденсированных 5 и 6-членных колец.
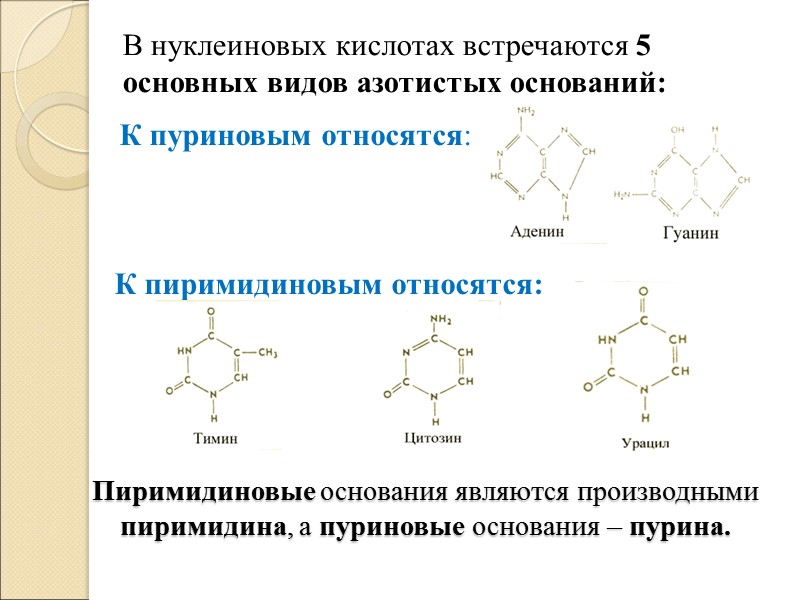
Пиримидиновые основания являются производными пиримидина, а пуриновые основания – пурина. В нуклеиновых кислотах встречаются 5 основных видов азотистых оснований: К пуриновым относятся: К пиримидиновым относятся:
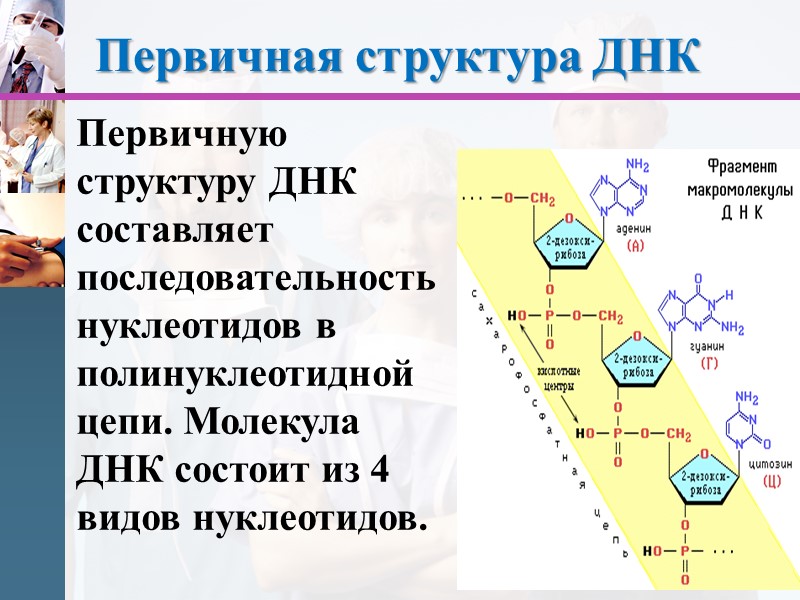
Первичная структура ДНК Первичную структуру ДНК составляет последовательность нуклеотидов в полинуклеотидной цепи. Молекула ДНК состоит из 4 видов нуклеотидов.

Больших успехов в определении структуры ДНКдостигли Эрвин Чаргафф и его сотрудники (1950г.) Используя метод хроматографии, они впервые определили нуклеотидный состав ДНК. Они установили, что соотношение азотистых оснований ДНК подчиняется универсальным. Правила Чаргаффа: 1. Сумма пуриновых нуклеотидов = сумме пиримидиновых нуклеотидов. 2. Молярное содержание аденинов = молярному содержанию тиминов и их отношение =1. 3. Количество аденина = количеству цитозина, а количество гуанина = количеству тимина, сумма аденина и цитозина = сумме гуанина и тимина.

Это супер спираль или кольцо более высокого порядка, представляет собой дальнейшую спирализацию и суперспирализацию молекулы ДНК. Третичная структура ДНК

Хромосомы эукариот представляют собой линейную молекулу ДНК. Эукариотическая ДНК обматывает белковые частицы- гистоны, располагающиеся вдоль ДНК.

2. Репликация ДНК (механизмы, ферменты). Повреждения ДНК. Репарация повреждений и ошибок репликации (механизмы, ферменты).
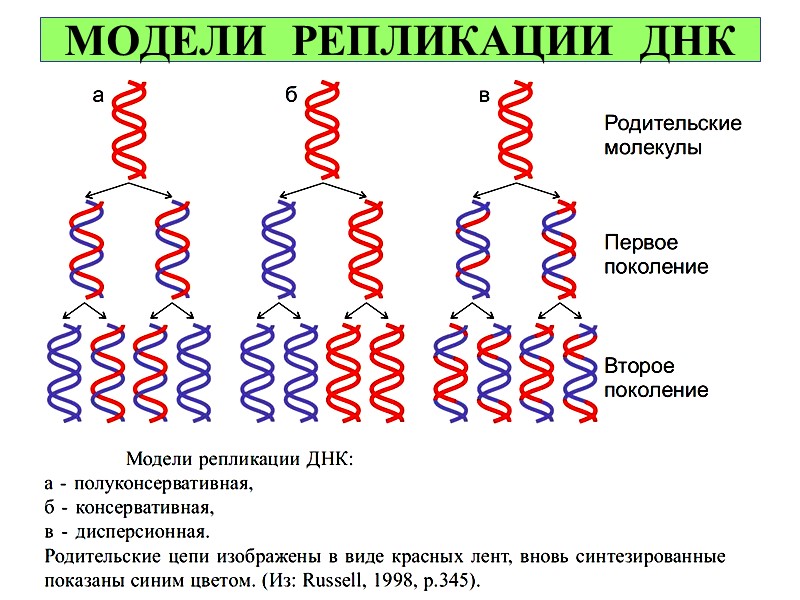
МОДЕЛИ РЕПЛИКАЦИИ ДНК

Репликон Единица, с помощью которой клетка контролирует отдельные акты репликации, получила название репликона. Каждый репликон в каждом клеточном цикле активизируется только один раз. В нем обязательно должны присутствовать необходимые для репликации контролирующие элементы: точка начала (origin), в которой инициируется репликация, точка окончания (terminus), в которой репликация останавливается.

ЭЛОНГАЦИЯ РЕПЛИКАЦИИ РНК-праймер ДНК-хеликаза РНК-праймер РНК-праймер Фрагменты Оказаки Праймаза (ДНК-полимераза α) ДНК-полимераза δ ДНК полимераза ε узнает РНК праймер и начинает синтезировать ДНК SSB-белки Лидирующая ДНК Отстающая ДНК

3. Структура и функции РНК (разновидности, структура). Биосинтез РНК и ее процессинг.

www.themegallery.com Нить ДНК, которая служит матрицей для синтеза РНК при транскрипции называется смысловой или матричной. Вторая нить ДНК называется некодирующей (комплементарная ей РНК не кодирует белки, она "бессмысленная").

Специфическая последовательность ДНК (сайт), в которой РНК-полимераза связывается с матрицей и начинает синтез РНК, называется промотором, а последовательность, на которой завершается синтез РНК, - сайтом терминации. Участок ДНК, ограниченный промотором и сайтом терминации, представляет собой единицу транскрипции -транскриптон. У эукариотов в состав транскриптона, как правило, входит только один ген.

Структура РНК-полимеразы РНК-полимераза — достаточно большая молекула. Основной фермент содержит 5 субъединиц (~400 кДа): α2: две α-субъединицы связывают остальные элементы фермента и распознают регулирующие факторы. Каждая субъединица состоит из двух доменов: αСКД (С-концевой домен) связывает первый элемент промотора, и αNКД (N-концевой домен) связывается с остальными компонентами полимеразы. β: эта субъединица обладает собственно полимеразным действием, катализируя синтез РНК. Она осуществляет инициацию процесса и управляет элонгацией. β': неспецифически связывается с ДНК. ω: восстанавливает денатурированную РНК-полимеразу обратно в дееспособную форму in vitro. Также обнаружено ее защитное/шаперонное действие на β'-субъединицу у Mycobacterium smegmatis. Для связывания с промоторными областями ДНК, основной фермент нуждается в еще одной субъединице — сигма (σ). Сигма-фактор значительно снижает сродство РНК-полимеразы к неспецифичным областям ДНК, и в то же время повышает ее чувствительность к определенным промоторам, в зависимости от своей структуры. С его помощью транскрипция начинается с нужного участка ДНК.

Эукариоты обладают различными типами РНК-полимераз, классифицируемыми по типам РНК, которые они производят: РНК-полимераза I, синтезирующая 45S-предшественника рРНК, превращающуюся затем в рРНК 28S, 18S и 5,8S, которые уже образуют главные РНК-секциирибосомы. РНК-полимераза II, производящая предшественников для мРНК, а также для большинства мяРНК и миРНК. Это наиболее хорошо изученный тип РНК-полимеразы. Ввиду того, что транскрипция должна происходить под строгим контролем, РНК-полимеразе II для связывания с промоторами требуется целый набор факторов транскрипции. РНК-полимераза III, синтезирующая тРНК, 5S рРНК и другие малые РНК, присутствующее в ядре и цитозоле.

Модификации пре-тРНК В процессе посттранскрипционных модификаций первичных транскриптов тРНК: • молекулы укорачиваются с 5'- и 3'-концов и удаляется интрон; • 10-15% азотистых оснований в молекулах модифицируется; • на 3'-конце формируется акцепторный участок (-ССА) для присоединения аминокислот, а в средней части антикодон - триплет нуклеотидов, обеспечивающий взаимодействие тРНК с кодоном мРНК.

Обратная транскрипция Некоторые вирусы (такие как ВИЧ, вызывающий СПИД), имеют возможность транскрибировать РНК в ДНК. ВИЧ имеет РНК-геном, который встраивается в ДНК. В результате, ДНК вируса может быть объединено с геномом клетки-хозяина. Главный фермент, ответственный за синтез ДНК из РНК, называется ревертазой. Одной из функций ревертазы является создание комплементарной ДНК (кДНК) из вирусного генома. Ассоциированый фермент рибонуклеаза H расщепляет РНК, а ревертаза синтезирует кДНК из двойной спирали ДНК. кДНК интегрируется в геном клетки-хозяина с помощью интегразы. Результатом является синтез вирусных протеинов клеткой-хозяином, которые образуют новые вирусы. В случае с ВИЧ так же программируется апоптоз (смерть клетки) Т-лимфоцитов. В иных случаях клетка может остаться распростанителем вирусов. Некоторые клетки эукариотов содержат фермент теломеразу, так же проявляющую активность обратной транскрипции. С её помощью синтезируются повторяющиеся последовательности в ДНК. Теломераза часто активирутся в раковых клетках для бесконечной дупликации генома без потери кодирующей протеины последовательности ДНК.

Классификация интронов Существует четыре группы интронов: Ядерные интроны Интроны группы I Интроны группы II Интроны группы III Иногда интроны группы III также относят к группе II, потому что они похожи по структуре и функции. Ядерные, или спалайсосомные интроны подвергаются сплайсингу при помощи сплайсосомы и малых ядерных РНК (мяРНК, snRNA). В последовательности РНК, содержащей ядерные интроны, есть специальные сигнальные последовательности, которые узнаются сплайсосомой. Интроны I, II и III группы способны к автосплайсингу и встречаются реже, чем сплайсосомные интроны.

4. Обмен нуклеотидов (состав, биосинтез, катаболизм). Нарушение обмена азотистых оснований (подагра).

Метаболизм нуклеотидов

ФУНКЦИИ НУКЛЕОТИДОВ

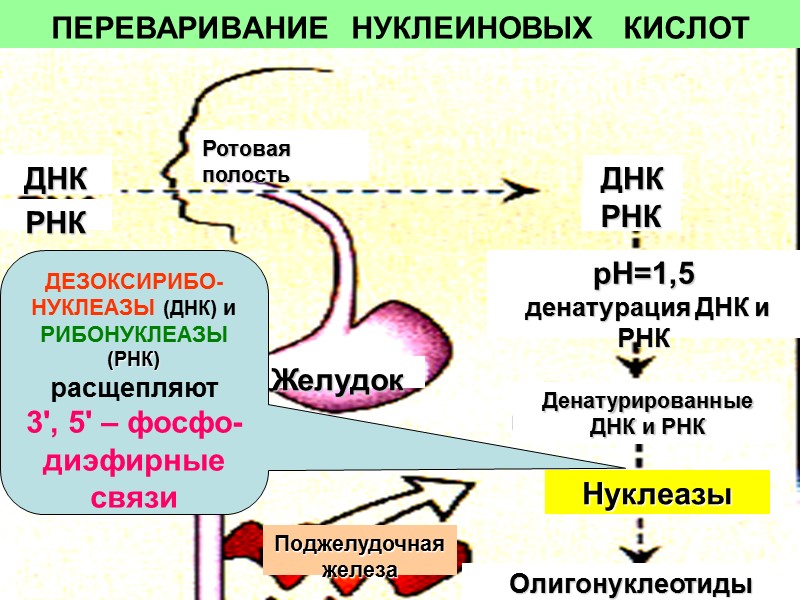
ПЕРЕВАРИВАНИЕ НУКЛЕИНОВЫХ КИСЛОТ Ротовая полость ДНК РНК ДНК РНК Желудок Поджелудочная железа рН=1,5 денатурация ДНК и РНК Денатурированные ДНК и РНК Нуклеазы Олигонуклеотиды ДЕЗОКСИРИБО-НУКЛЕАЗЫ (ДНК) и РИБОНУКЛЕАЗЫ (РНК) расщепляют 3', 5' – фосфо-диэфирные связи

Поджелудочная железа Олигонуклеотиды Фосфодиэстеразы Мононуклеотиды Нуклеотидазы Нуклеозиды Нуклеозидазы Пиримидины Пурины (Дезокси) рибоза Кровоток Мочевая кислота Моча Тонкий кишечник Ф Энтероцит

МЕТАБОЛИЗМ ПУРИНОВ

ИСТОЧНИКИ АТОМОВ В ПУРИНОВЫХ ОСНОВАНИЯХ АСПАРТАТ ГЛИЦИН ГЛУТАМИН ФОРМИЛ ТГФК СЕРИН

Особенность синтеза пуринов Атомы С и N поэтапно добавляются к рибозе 5-фосфат.

Синтез ФРПФ (ФРДФ) Активатор Ингибиторы Пуриновые нуклеотиды Рибозо-5-фосфат 5-фосфорибозил-1-пирофосфат (ФРПФ) АТФ АМФ ФРПФ синтетаза

ЛИМИТИРУЮЩАЯ РЕАКЦИЯ 5-фосфорибозиламин Глутамин Глутамат Ингибиторы АМФ, ГМФ, ИМФ 5-фосфорибозил 1-пирофосфат (ФРПФ) Глутамин фосфорибозил пирофосфат амидо- трансфераза [S] 3.

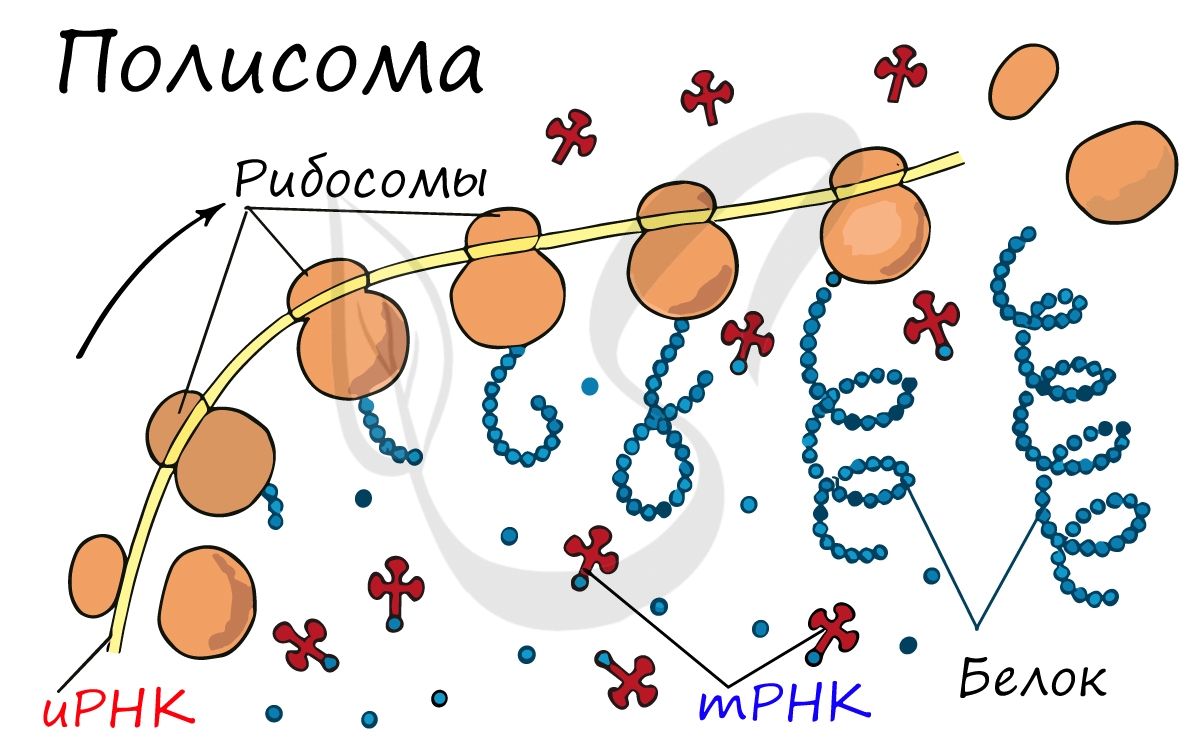
Полисома Одновременно несколько рибосом могут участвовать в трансляции одной мРНК. Каждая рибосома занимает участок, равный примерно 80 нуклеотидам мРНК. Таким образом, рибосомы располагаются на мРНК с интервалами около 100 нуклеотидов, образуя комплекс, называемый полисомой.

Функционально активные белки образуются в результате посттрансляционных модификаций полипептидных цепей, синтезированных на рибосомах. Они включают: частичный протеолиз; фолдинг, или формирование пространственной структуры, в котором принимают участие белки-шапероны, обеспечивающие образование функционально активной конформации полипептидной цепи; модификации аминокислот: карбоксилирование, фосфорилирование, йодирование, гидроксилирование, ацилирование и гликозилирование; образование дисульфидных связей между остатками цистеина, участвующими в формировании трехмерной структуры белка; присоединение простетических групп; образование олигомерных структур, которое также осуществляется при участии шаперонов.

Структурная часть гена Промотор Начало транскрипции Старт-кодон Стоп - кодон 5’ 3’ 3’ 5’ Терминатор транскрипции Строение гена Участок связывания фермента РНК-полимеразы (место начала транскрипции) Участок, кодирующий последовательность аминокислот в молекуле белка. Триплет ТАЦ в ДНК и АУГ в РНК иницирующий начало синтеза белка Триплеты – бессмысленные кодоны на которых завершается трансляция Участок, сигнализирующий о прекращении транскрипции

Виды оперонов Оперон — это тесно связанная последовательность структурных генов, определяющих синтез группы белков, которые участвуют в одной цепи биохимических преобразований. К наиболее хорошо изученным оперонам бактерий относят: Лактозный (lac)-оперон Галактозный (gal)-оперон Триптофановый (trp)-оперон

Лактозный оперон Оперон – участок бактериальной хромосомы, включающий следующие участки ДНК: Р – промотор, О – оператор, Z, Y, А – структурные гены, Т –терминатор. (В состав других оперонов может входить до 10 структурных генов.) Промотор – место присоединения РНК- полимеразы. Оператор – участок оперона, к которому присоединяются белки-репрессоры или активаторы транскрипции. Структурные гены – гены кодирующие ферменты, участвующие в метаболизме лактозы. Терминатор служит для отсоединения РНК-полимеразы после окончания синтеза иРНК, соответствующей ферментам Z, Y, А, необходимым для усвоения лактозы.
И транскрипция, и трансляция относятся к матричным биосинтезам. Матричным биосинтезом называется синтез биополимеров (нуклеиновых кислот, белков) на матрице - нуклеиновой кислоте ДНК или РНК. Процессы матричного биосинтеза относятся к пластическому обмену: клетка расходует энергию АТФ.
Матричный синтез можно представить как создание копии исходной информации на несколько другом или новом "генетическом языке". Скоро вы все поймете - мы научимся достраивать по одной цепи ДНК другую, переводить РНК в ДНК и наоборот, синтезировать белок с иРНК на рибосоме. В данной статье вас ждут подробные примеры решения задач, генетический словарик пригодится - перерисуйте его себе :)

Возьмем 3 абстрактных нуклеотида ДНК (триплет) - АТЦ. На иРНК этим нуклеотидам будут соответствовать - УАГ (кодон иРНК). тРНК, комплементарная иРНК, будет иметь запись - АУЦ (антикодон тРНК). Три нуклеотида в зависимости от своего расположения будут называться по-разному: триплет, кодон и антикодон. Обратите на это особое внимание.
Репликация ДНК - удвоение, дупликация (лат. replicatio — возобновление, лат. duplicatio - удвоение)
Процесс синтеза дочерней молекулы ДНК по матрице родительской ДНК. Нуклеотиды достраивает фермент ДНК-полимераза по принципу комплементарности. Переводя действия данного фермента на наш язык, он следует следующему правилу: А (аденин) переводит в Т (тимин), Г (гуанин) - в Ц (цитозин).
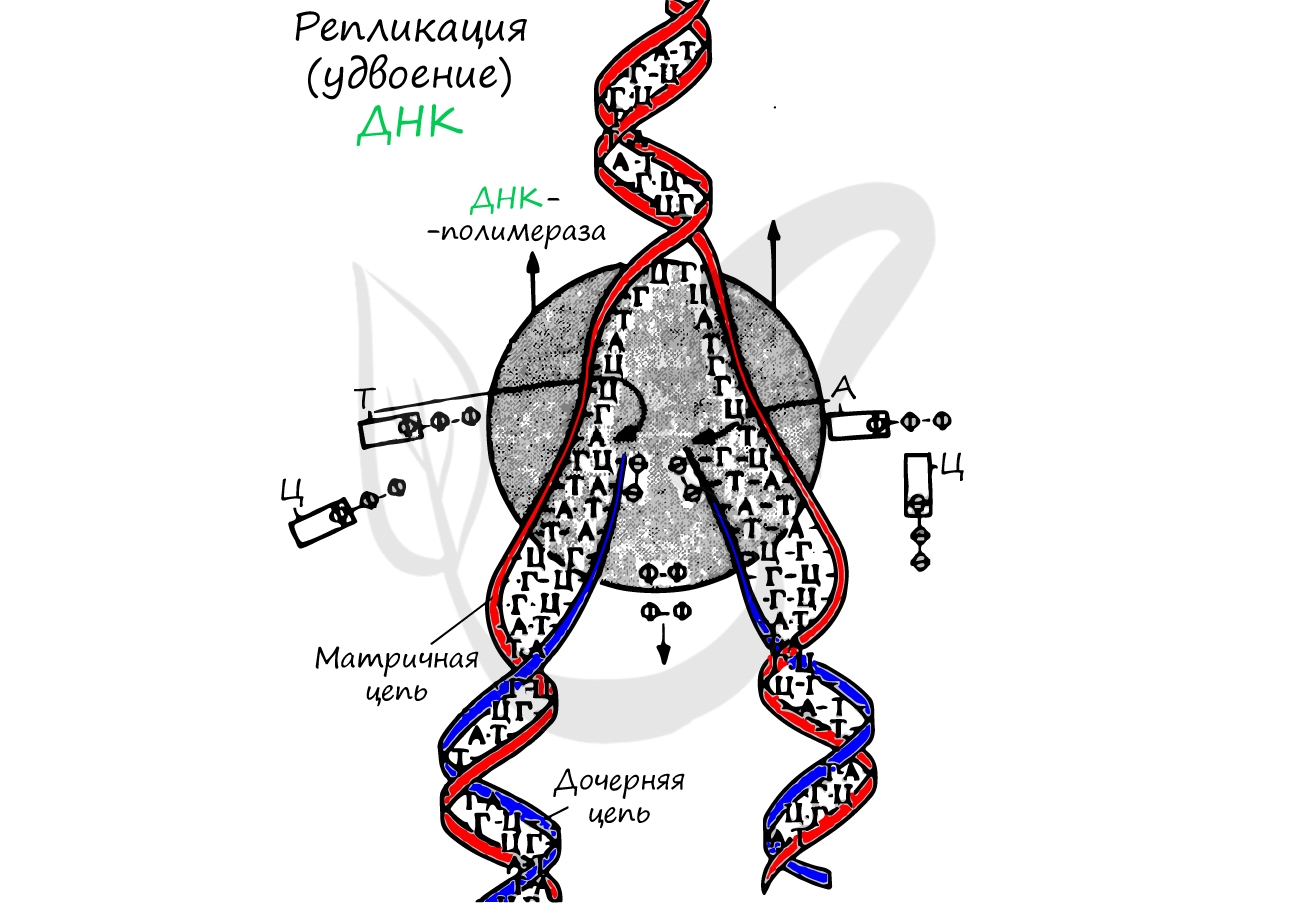
Удвоение ДНК происходит в синтетическом периоде интерфазы. При этом общее число хромосом не меняется, однако каждая из них содержит к началу деления две молекулы ДНК: это необходимо для равномерного распределения генетического материала между дочерними клетками.
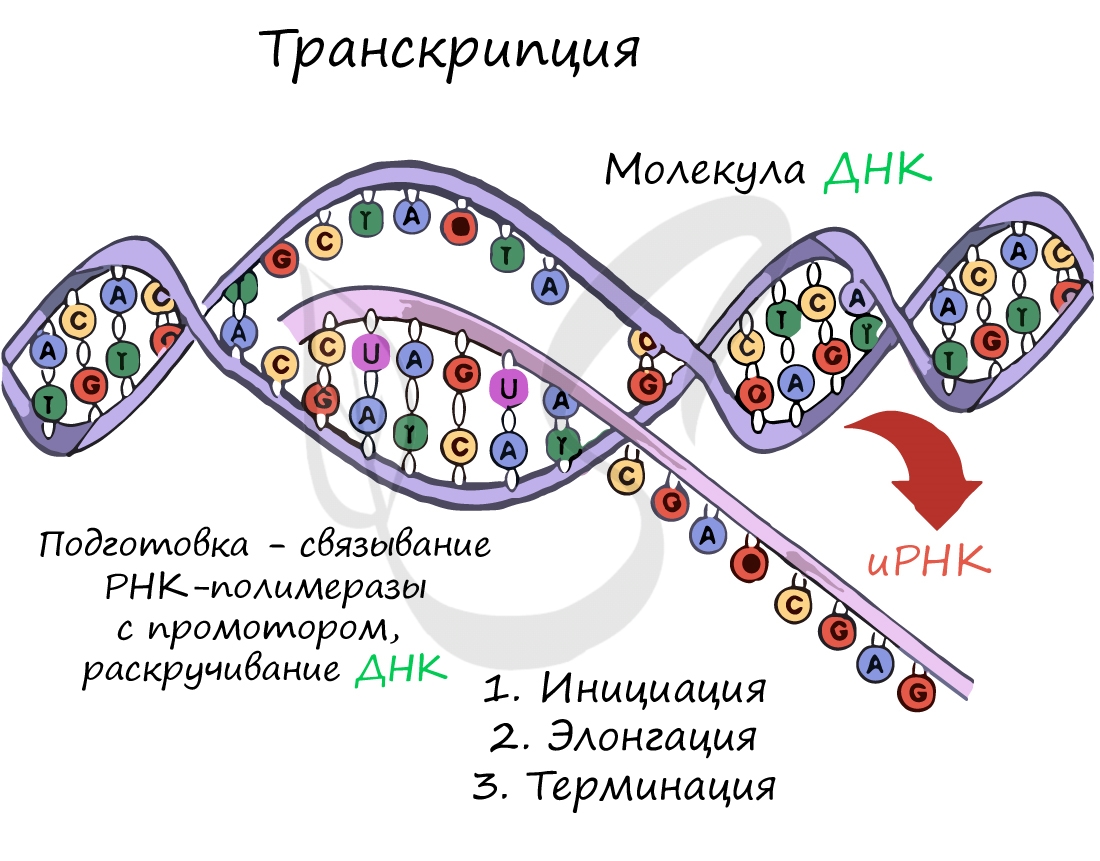
Транскрипция (лат. transcriptio — переписывание)
Транскрипция представляет собой синтез информационной РНК (иРНК) по матрице ДНК. Несомненно, транскрипция происходит в соответствии с принципом комплементарности азотистых оснований: А - У, Т - А, Г - Ц, Ц - Г (загляните в "генетический словарик" выше).
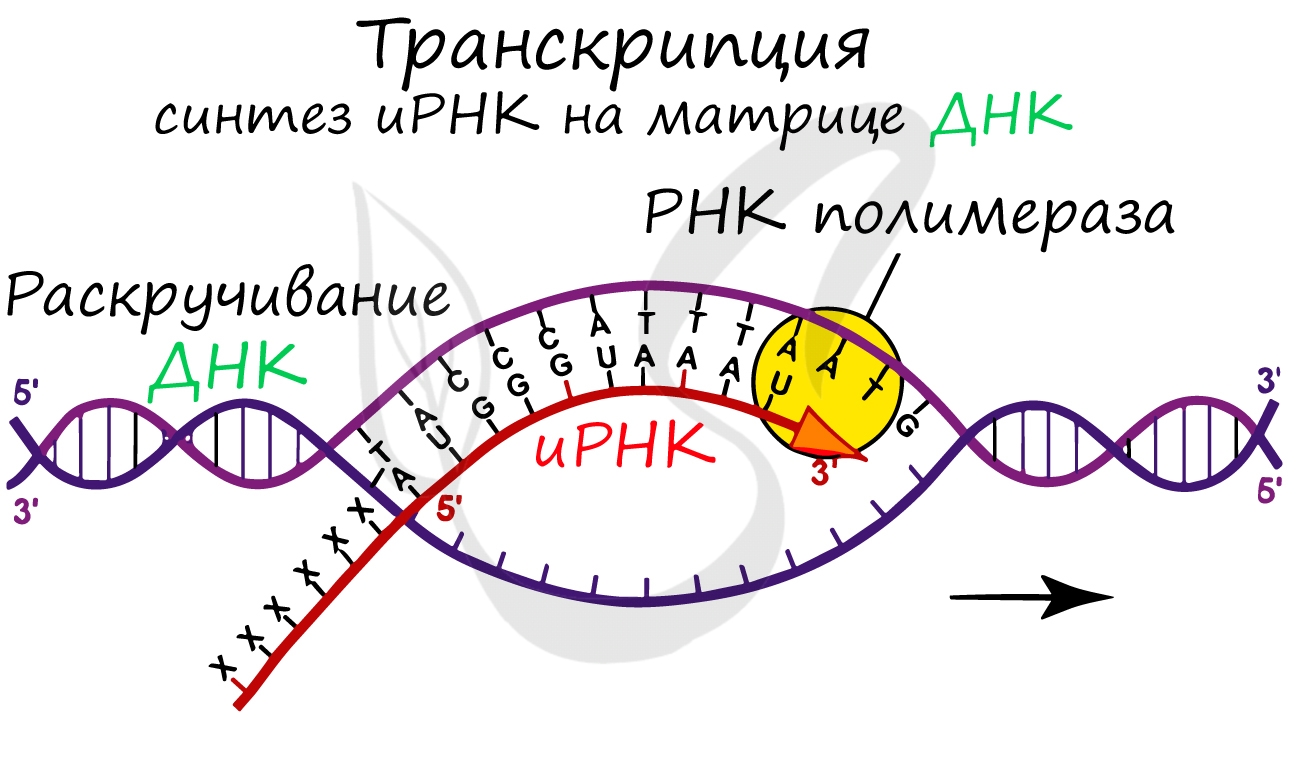
До начала непосредственно транскрипции происходит подготовительный этап: фермент РНК-полимераза узнает особый участок молекулы ДНК - промотор и связывается с ним. После связывания с промотором происходит раскручивание молекулы ДНК, состоящей из двух цепей: транскрибируемой и смысловой. В процессе транскрипции принимает участие только транскрибируемая цепь ДНК.
-
Инициация (лат. injicere — вызывать)
Образуется несколько начальных кодонов иРНК.
Нити ДНК последовательно расплетаются, освобождая место для передвигающейся РНК-полимеразы. Молекула иРНК быстро растет.
Достигая особого участка цепи ДНК - терминатора, РНК-полимераза получает сигнал к прекращению синтеза иРНК. Транскрипция завершается. Синтезированная иРНК направляется из ядра в цитоплазму.
Трансляция (от лат. translatio — перенос, перемещение)
Куда же отправляется новосинтезированная иРНК в процессе транскрипции? На следующую ступень - в процесс трансляции. Он заключается в синтезе белка на рибосоме по матрице иРНК. Последовательность кодонов иРНК переводится в последовательность аминокислот.
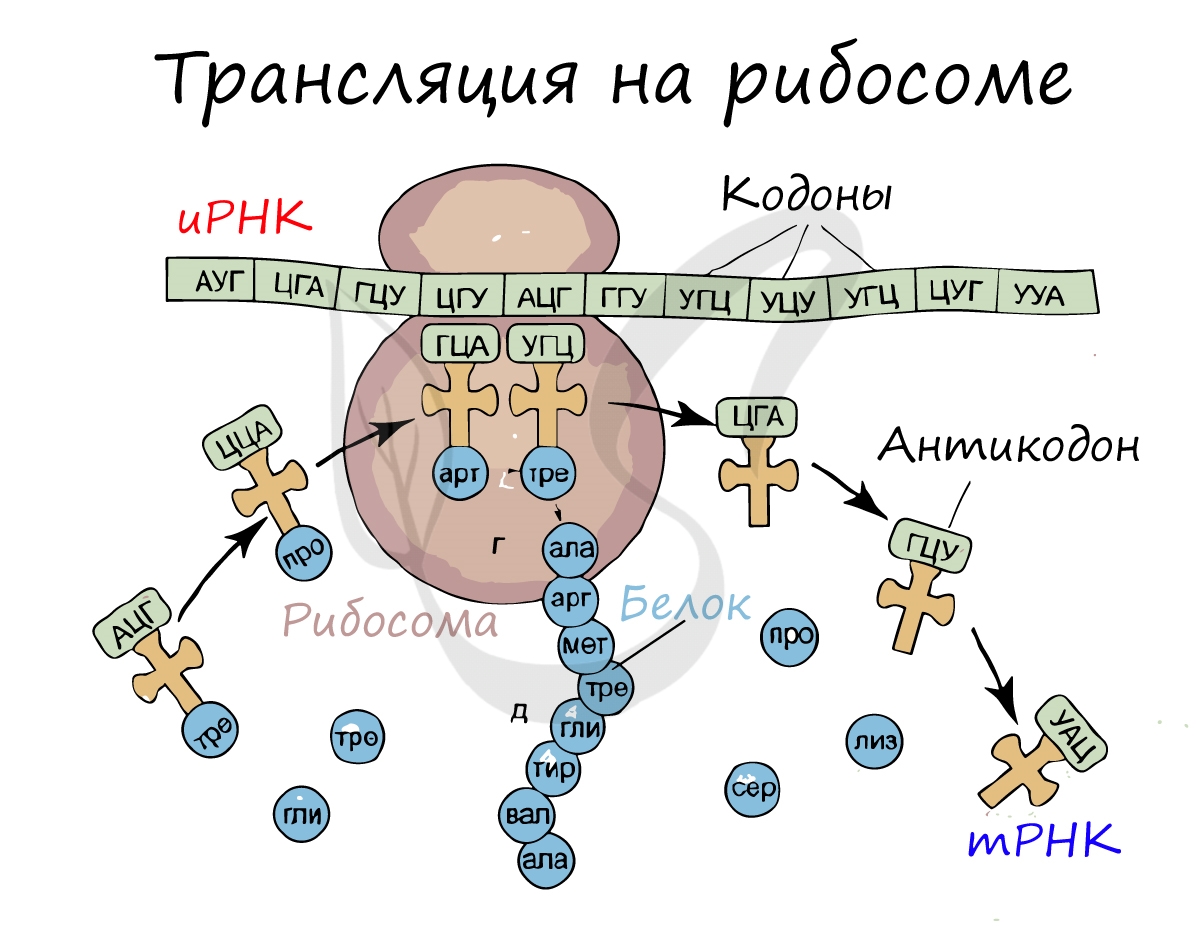
Информационная РНК (иРНК, синоним - мРНК (матричная РНК)) присоединяется к рибосоме, состоящей из двух субъединиц. Замечу, что вне процесса трансляции субъединицы рибосом находятся в разобранном состоянии.
Первый кодон иРНК, старт-кодон, АУГ оказывается в центре рибосомы, после чего тРНК приносит аминокислоту, соответствующую кодону АУГ - метионин.
Рибосома делает шаг, и иРНК продвигается на один кодон: такое в фазу элонгации происходит десятки тысяч раз. Молекулы тРНК приносят новые аминокислоты, соответствующие кодонам иРНК. Аминокислоты соединяются друг с другом: между ними образуются пептидные связи, молекула белка растет.
Доставка нужных аминокислот осуществляется благодаря точному соответствию 3 нуклеотидов (кодона) иРНК 3 нуклеотидам (антикодону) тРНК. Язык перевода между иРНК и тРНК выглядит как: А (аденин) - У (урацил), Г (гуанин) - Ц (цитозин). В основе этого также лежит принцип комплементарности.
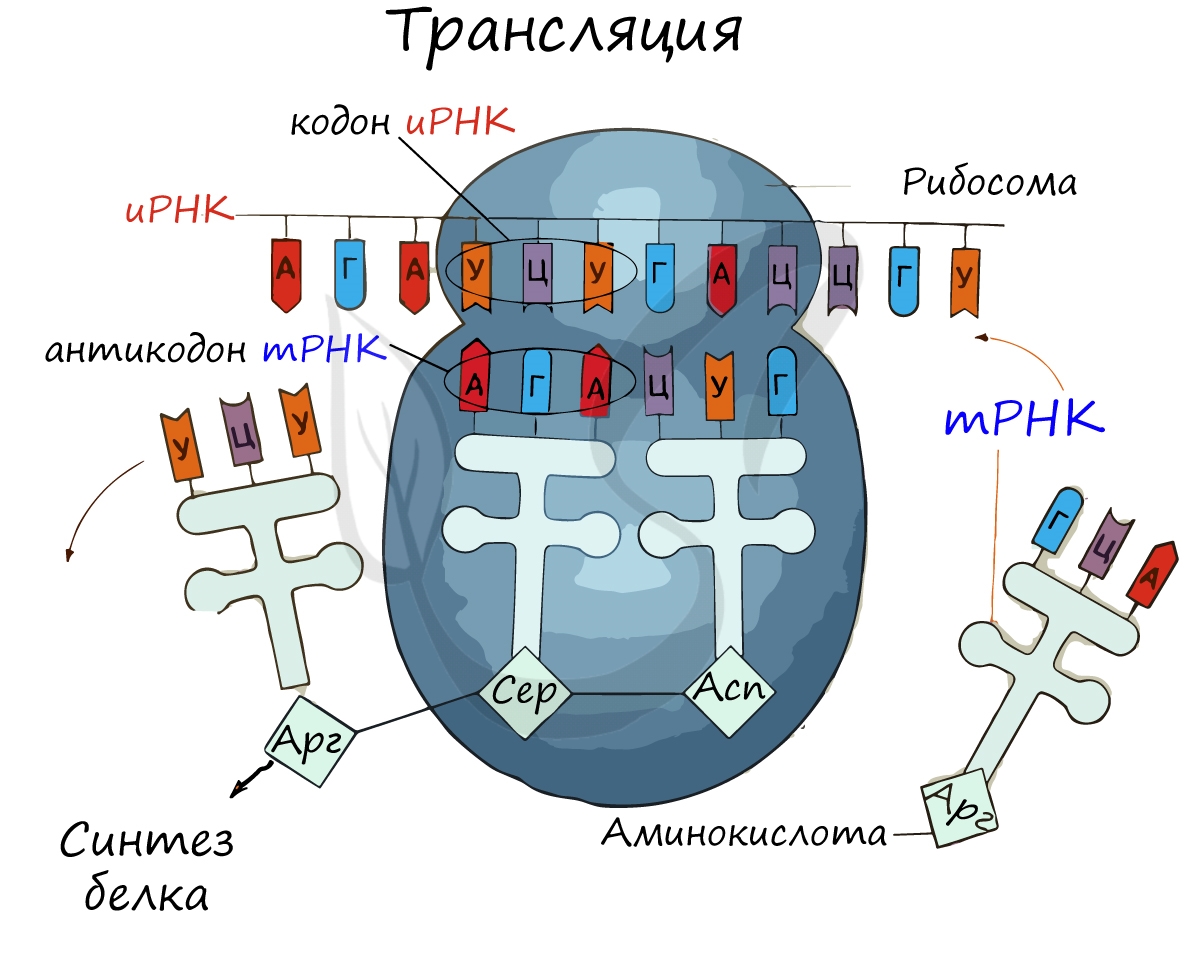
Движение рибосомы вдоль молекулы иРНК называется транслокация. Нередко в клетке множество рибосом садятся на одну молекулу иРНК одновременно - образующаяся при этом структура называется полирибосома (полисома). В результате происходит одновременный синтез множества одинаковых белков.
Синтез белка - полипептидной цепи из аминокислот - в определенный момент завершатся. Сигналом к этому служит попадание в центр рибосомы одного из так называемых стоп-кодонов: УАГ, УГА, УАА. Они относятся к нонсенс-кодонам (бессмысленным), которые не кодируют ни одну аминокислоту. Их функция - завершить синтез белка.
Существует специальная таблица для перевода кодонов иРНК в аминокислоты. Пользоваться ей очень просто, если вы запомните, что кодон состоит из 3 нуклеотидов. Первый нуклеотид берется из левого вертикального столбика, второй - из верхнего горизонтального, третий - из правого вертикального столбика. На пересечении всех линий, идущих от них, и находится нужная вам аминокислота :)
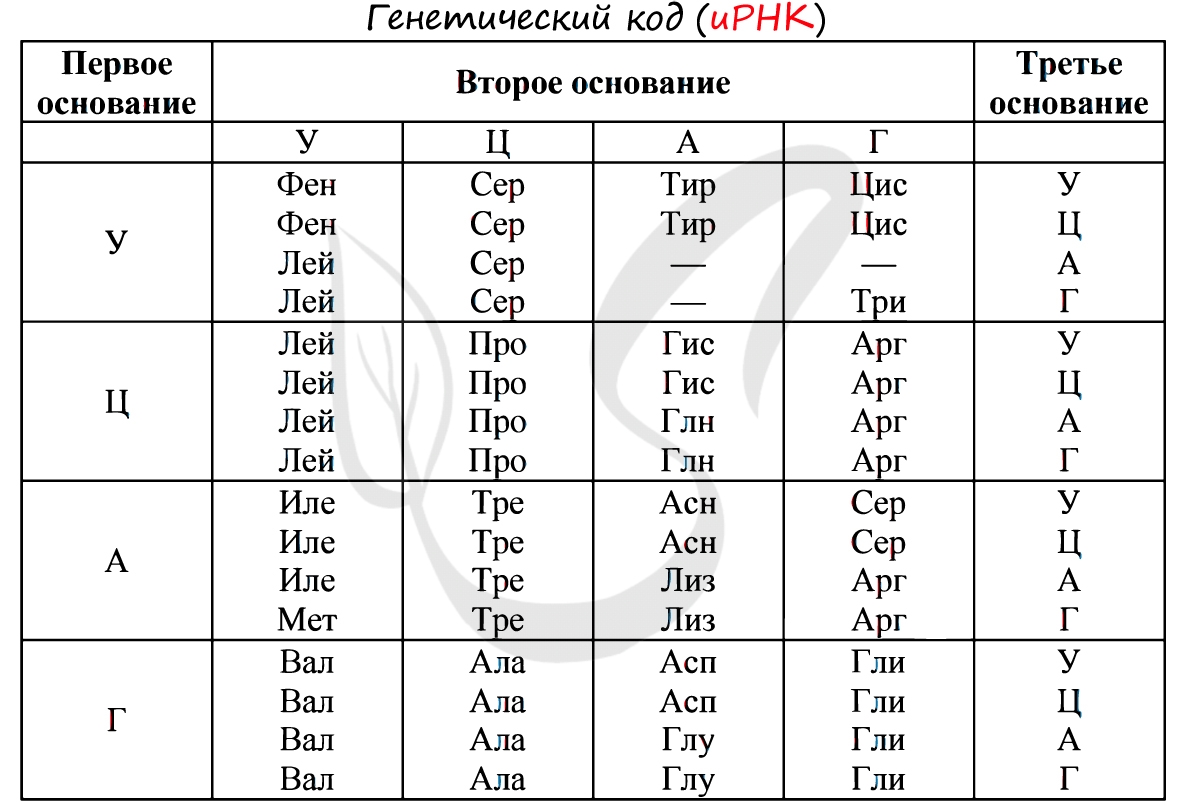
Давайте потренируемся: кодону ЦАЦ соответствует аминокислота Гис, кодону ЦАА - Глн. Попробуйте самостоятельно найти аминокислоты, которые кодируют кодоны ГЦУ, ААА, УАА.
Кодону ГЦУ соответствует аминокислота - Ала, ААА - Лиз. Напротив кодона УАА в таблице вы должны были обнаружить прочерк: это один из трех нонсенс-кодонов, завершающих синтез белка.
Примеры решения задачи №1
Без практики теория мертва, так что скорее решим задачи! В первых двух задачах будем пользоваться таблицей генетического кода (по иРНК), приведенной вверху.
"Фрагмент цепи ДНК имеет следующую последовательность нуклеотидов: ЦГА-ТГГ-ТЦЦ-ГАЦ. Определите последовательность нуклеотидов во второй цепочке ДНК, последовательность нуклеотидов на иРНК, антикодоны соответствующих тРНК и аминокислотную последовательность соответствующего фрагмента молекулы белка, используя таблицу генетического кода"

По принципу комплементарности мы нашли вторую цепочку ДНК: ГЦТ-АЦЦ-АГГ-ЦТГ. Мы использовали следующие правила при нахождении второй нити ДНК: А-Т, Т-А, Г-Ц, Ц-Г.
Вернемся к первой цепочке, и именно от нее пойдем к иРНК: ГЦУ-АЦЦ-АГГ-ЦУГ. Мы использовали следующие правила при переводе ДНК в иРНК: А-У, Т-А, Г-Ц, Ц-Г.
Зная последовательность нуклеотидов иРНК, легко найдем тРНК: ЦГА, УГГ, УЦЦ, ГАЦ. Мы использовали следующие правила перевода иРНК в тРНК: А-У, У-А, Г-Ц, Ц-Г. Обратите внимание, что антикодоны тРНК мы разделяем запятыми, в отличие кодонов иРНК. Это связано с тем, что тРНК представляют собой отдельные молекулы (в виде клеверного листа), а не линейную структуру (как ДНК, иРНК).
Пример решения задачи №2
"Известно, что все виды РНК синтезируются на ДНК-матрице. Фрагмент цепи ДНК, на которой синтезируется участок центральной петли тРНК, имеет следующую последовательность нуклеотидов: ТАГ-ЦАА-АЦГ-ГЦТ-АЦЦ. Установите нуклеотидную последовательность участка тРНК, который синтезируется на данном фрагменте, и аминокислоту, которую будет переносить эта тРНК в процессе биосинтеза белка, если третий триплет соответствует антикодону тРНК"

Обратите свое пристальное внимание на слова "Известно, что все виды РНК синтезируются на ДНК-матрице. Фрагмент цепи ДНК, на которой синтезируется участок центральной петли тРНК ". Эта фраза кардинально меняет ход решения задачи: мы получаем право напрямую и сразу синтезировать с ДНК фрагмент тРНК - другой подход здесь будет считаться ошибкой.
Итак, синтезируем напрямую с ДНК фрагмент молекулы тРНК: АУЦ-ГУУ-УГЦ-ЦГА-УГГ. Это не отдельные молекулы тРНК (как было в предыдущей задаче), поэтому не следует разделять их запятой - мы записываем их линейно через тире.
Третий триплет ДНК - АЦГ соответствует антикодону тРНК - УГЦ. Однако мы пользуемся таблицей генетического кода по иРНК, так что переведем антикодон тРНК - УГЦ в кодон иРНК - АЦГ. Теперь очевидно, что аминокислота кодируемая АЦГ - Тре.
Пример решения задачи №3
Длина фрагмента молекулы ДНК составляет 150 нуклеотидов. Найдите число триплетов ДНК, кодонов иРНК, антикодонов тРНК и аминокислот, соответствующих данному фрагменту. Известно, что аденин составляет 20% в данном фрагменте (двухцепочечной молекуле ДНК), найдите содержание в процентах остальных нуклеотидов.

Один триплет ДНК состоит из 3 нуклеотидов, следовательно, 150 нуклеотидов составляют 50 триплетов ДНК (150 / 3). Каждый триплет ДНК соответствует одному кодону иРНК, который в свою очередь соответствует одному антикодону тРНК - так что их тоже по 50.
По правилу Чаргаффа: количество аденина = количеству тимина, цитозина = гуанина. Аденина 20%, значит и тимина также 20%. 100% - (20%+20%) = 60% - столько приходится на оставшиеся цитозин и гуанин. Поскольку их процент содержания равен, то на каждый приходится по 30%.
Теперь мы украсили теорию практикой. Что может быть лучше при изучении новой темы? :)
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
нования пяти типов, пентозы двух типов и остатки фосфорной кислоты.
Азотистые основания в нуклеиновых кислотах могут быть пуриновыми (аденин,
гуанин) и пиримидиновыми (цитозин, урацил, тимин).
В зависимости от строения углевода выделяют рибонуклеиновые кислоты – содержат рибозу ( РНК ), и дезоксирибонуклеиновые кислоты – содержат дезоксирибо-
О СНОВНОЙ ПОСТУЛАТ МОЛЕКУЛЯРНОЙ БИОЛОГИИ
В подавляющем большинстве случаев передача наследственной информации
от материнской клетки к дочерней осуществляется при помощи ДНК. Для использо-
вания генетической информации самой клеткой необходимы РНК, образуемые на матрице ДНК. Далее РНК непосредственно участвуют на всех этапах синтеза белко-
вых молекул, обеспечивающих структуру и деятельность клетки.
На вышесказанном основана центральная догма молекулярной биологии , со-
гласно которой перенос генети-
ческой информации осуществля-
ется только от нуклеиновой кислоты (ДНК и РНК). Получателем
информации может быть другая
нуклеиновая кислота (ДНК или РНК) и белок.
С ТРОЕНИЕ ДЕЗОКСИРИБОНУКЛЕИНОВОЙ КИСЛОТЫ
ДНК – наиболее важная часть
хромосом: две двухцепочечные
молекулы ДНК образуют одну хромосому. Наиболее хорошо
хромосомы видны перед митозом
и во время его. В покоящихся клетках хромосомный материал
выглядит нечетко и распределен
по всему ядру. В таком состоя-
нии он получил название "хрома-
тин". В составе хроматина выде-
ляют 60% белка (гистоны и кислые белки), 35% ДНК и около 5% РНК.
Хроматин уложен в виде сферических частиц – нуклеосом, соединенных друг с
другом нитью ДНК. Нуклеосома представляет собой комплекс участка молекулы ДНК и восьми молекул гистонов. В составе нуклеосомы находятся по 2 молекулы гисто-
нов Н2α, Н2β, Н3, Н4. Нить ДНК последовательно контактируя с гистонами Н2α, Н2β,
Н4, Н3, Н3, Н4, Н2β, Н2α, наматывается на гистоновое ядро, которое "маскирует" 146 пар оснований ДНК. Гистон Н1 связывается с нуклеосомой на участке входа и
выхода ДНК, "склеивая" 2 оборота и "маскируя" еще 20 пар оснований. Всего за-
маскировано 166 пар оснований. Кроме нуклеосом, в ядре присутствуют еще 2
структуры: фибриллы диаметром 10 нм, состоящие из цепочки нуклеосом, и волокна, диаметром 30 нм, образующиеся при закручивании фибрилл в спираль. На виток спирали приходится 6-7 нуклеосом. Участок ДНК между нуклеосомами называется спейсерным (англ: space – пространство), его длина варьирует в зависимости от вида организма и типа клеток. У человека она составляет около 50 пар нуклеотидов.
Благодаря наличию нуклеосом достигается уменьшение размеров хромосомы в 7 раз, далее происходит укладка в суперспираль и „суперсуперспираль". Таким образом, благодаря гистонам размеры ДНК уменьшаются в тысячи раз: если длина ДНК достигает 6-9 см (10 -1 ), то размеры хромосом – всего несколько микрометров (10 -6 ).
Хроматин может быть активным (эухроматин) и неактивным (гетерохроматин). Активный хроматин содержит активные гены, т.е. те гены, с которых считывается информация. В активном хроматине нуклеосомная структура изменена или вообще отсутству-
ет, благодаря чему ДНК становится доступной для со-
Рибонуклеиновая кислота (РНК) представляет со-
бой последовательность рибонуклеозидмонофосфа-
тов, связанных друг с другом 5’-3’-фосфодиэфирными связями. РНК отличается от ДНК однонитевой струк-
турой, наличием урацила вместо тимина и рибозы вместо дезоксирибозы
В клетке присутствует четыре типа РНК:
Рибосомальные РНК (рРНК) у прокариот и эукариот различны и отличаются величиной седиментации (S, величиной скорости оседания молекулы). У прокариот три
разновидности рРНК: 5S, 16S и 23S. У эукариот четыре разновидности: 5S, 5,8S, 18S
и 28S. Рибосомальные РНК участвуют в построении рибосом, внутриклеточных белоксинтезирующих органелл.
Рибосомы состоят из двух неравных субчастиц, малой и большой.
• малую (30S) субчастицу образуют белки, 23S-рРНК и 5S-рРНК;
• большую (50S) – белки и 16S-рРНК.
• малую (40S) субчастицу образуют белки и 18S-рРНК,
• большую (60S) – белки и 5S-, 5,8S-, 28S-рРНК.
Матричные РНК (мРНК) представляют собой линейную последовательность нуклеотидов. К 5’-концу молекулы присоеди-
нен метилгунозиндифосфат, на 3’-конце име-
ется полиадениловая последовательность. Их функция – информационная, т.е. перенос информации о структуре белков от ДНК к месту
Транспортные РНК (тРНК) бактерий и эу-
кариот включают 73-93 нуклеотида. Они пере-
носят аминокислоты из цитозоля к рибосомам. Вторичная структура тРНК напоминает кле-
верный лист, а третичная – латинскую букву L .
ет собственную функцию: антикодоновый,
псевдоуридиловый, дигидроуридиловый, акцепторный. На 5’-конце тРНК находится гуани-
ловый нуклеотид, на 3’-конце – триплет Ц-Ц-А.
• • Малые РНК – используются для созревания мРНК и некоторых других клеточ-
Синтез ДНК в клетке происходит не беспорядоч-
но, а в строго определенный период жизни клетки.
Всего выделяют 4 фазы: митоз (М), синтетическую (S), пресинтетическую (G1, от англ. gap - интервал),
Важное участие в регуляции смены фаз клеточного цикла занимают циклины – белки массой
35-90 кДа, уровень которых меняется в ходе клеточ-
ного цикла. По функции циклины – это активаторные
субъединицы ферментов циклин-зависимых киназ (ЦЗК) . Активные комплексы циклин-ЦЗК фосфорили-
руют внутриклеточные белки, изменяя их активность. Этим обеспечивается продвижение по клеточному циклу.
Синтез (репликация, удвоение) ДНК про-
исходит в S-фазу клеточного цикла.
Механизм репликации, как установили эксперименты Мэтью Мезельсон и Франклин
Сталь в 1957 г, полуконсервативный, т.е.
на каждой нити материнской ДНК синтезируется дочерняя копия.
Весь процесс репликации идет в S-фазу
клеточного цикла, в то время, когда клетка готовится к делению.
Как матричный биосинтез, репликация
требует наличия нескольких условий:
• Матрица – в ее роли выступает материнская ДНК;
• Растущая цепь – дочерняя ДНК;
• Субстраты для синтеза – dАТФ, dГТФ, dЦТФ, ТТФ;
• Источник энергии – dАТФ, dГТФ, dЦТФ, ТТФ;
Синтез ДНК начинается в определенных участках, получивших название точка o r i (англ. origin - начало). На каждой ДНК млекопитающих точек o r i насчитывается
около 100. Репликация распространяется от этих участков в обе стороны по нитям ДНК с образованием " репликативных пузырей" . В каждом таком "пузыре" имеются
две " репликативные вилки ", в которых происходит расплетание, раскручивание и
непосредственный синтез ДНК. Репликативные вилки удаляются друг от друга. В целом вся репликация ДНК у эукариот заканчивается за 9 часов.
В каждой репликативной вилке идет синтез ДНК в направлении от 5'-конца к
3'-концу, т.е. 5'-конец новой ДНК остается свободным, следующие нуклеотиды при-
соединяются к 3'-гидроксильной группе предыдущего нуклеотида. Поскольку нити ДНК антипараллельны, то непрерывно синтезируется только одна нить, а именно та, на которой направление движения репликативной вилки совпадает с направлением
По мере расплетания и движения репликативной вилки на нити открываются
участки, где возможен синтез новой нити в направлении 5' → 3'.
Направление 5' → 3' другой материнской нити ДНК совпадает с направлением движения репликативной вилки. Поэтому синтез дочерней нити (в направлении 5' → 3') возможен только после расплетания части ДНК и освобождения участка для синтеза.
Таким образом, синтез дочерней ДНК на одной из нитей материнской ДНК идет фрагментарно. По имени японского исследователя синтезируемые на отстающей цепи отрезки ДНК назвали фрагменты Оказаки.
РАЗДЕЛ 4. БИОСИНТЕЗ НУКЛЕИНОВЫХ КИСЛОТ И БЕЛКОВ (МАТРИЧНЫЕ БИОСИНТЕЗЫ). ОСНОВЫ МОЛЕКУЛЯРНОЙ ГЕНЕТИКИ
II. Репликация
Живые организмы в течение S-фазы клеточного цикла, которая предшествует делению клетки, удваивают содержание ДНК таким образом, что каждая дочерняя клетка после деления получает набор хромосом, идентичный родительской клетке. Процесс удвоения хромосом называют репликацией (редупликацией).
Рис. 4-14. Полуконсервативная репликация.

Ферменты и белки, участвующие в репликации, должны работать быстро и точно. Эти условия выполняются с помощью особого мультиферментного комплекса.
Репликацию можно разделить на 4 этапа: образование репликативной вилки (инициация), синтез новых цепей (элонгация), исключение праймеров, завершение синтеза двух дочерних цепей ДНК (терминация).
А. Иницифция репликации
Синтез ДНК у эукариотов происходит в S-фазу клеточного цикла. Инициацию репликации регулируют специфические сигнальные белковые молекулы — факторы роста.Факторы роста связываются рецепторами мембран клеток, которые передают сигнал, побуждающий клетку к началу репликации (см. раздел 11).
Синтез новых одноцепочечных молекул ДНК может произойти только при расхождении родительских цепей. В определённом сайте (точка начала репликации) происходит локальная денатурация ДНК, цепи расходятся и образуются две репликативные вилки, движущиеся в противоположных направлениях.
В образовании репликативной вилки принимает участие ряд белков и ферментов. Так, семейство ДНК-топоизомераз (I, II и III), обладая нуклеазной активностью, участвует в регуляции суперспирализации ДНК. Например, ДНК-топоизомераза I разрывает фосфоэфирную связь в одной из цепей двойной спирали и ковалентно присоединяется к 5'-концу в точке разрыва (рис. 4-15). По окончании формирования репликативной вилки фермент ликвидирует разрыв в цепи и отделяется от ДНК.
Рис. 4-15. Участие ДНК-топоизомеразы I в образовании репликативной вилки. 1 — фермент расщепляет одну цепь ДНК; между остатком тирозина молекулы фермента и фосфорным остатком цепи образуется ковалентная связь; 2 — происходит локальное раскручивание двойной спирали при участии ДНК-хеликазы; ДНК-топоизомераза I восстанавливает фосфоэфирную связь.

Разрыв водородных связей в двухцепочечной молекуле ДНК осуществляет ДНК-хеликаза. Фермент ДНК-хеликаза использует энергию АТФ для расплетения двойной спирали ДНК.
Репликация ДНК осуществляется ДНК-зависимыми ДНК-полимеразами (рис. 4-16). Субстратами и источниками энергии для синтеза продукта служат 4 макроэргических соединения — дезоксирибонуклеозидтрифосфаты дАТФ, дГТФ, дЦТФ и дТТФ, для активации которых необходимы ионы магния. Нейтрализуя отрицательный заряд нуклеотидов, они повышают их реакционную способность. Ферменты проявляют каталитическую активность только в присутствии предварительно раскрученной матричной двухцепочечной ДНК. Синтез цепей ДНК происходит в направлении 5' —> 3' растущей цепи, т. е. очередной нуклеотид присоединяется к свободному 3'-ОН-концу предшествующего нуклеотидного остатка. Синтезируемая цепь всегда антипараллельна матричной цепи. В ходе репликации образуются 2 дочерние цепи, представляющие собой копии матричных цепей.
Рис. 4-16. Репликация.

В синтезе эукариотических ДНК принимают участие 5 ДНК-полимераз (α, β, y, δ, ε). ДНК- полимеразы различают по числу субъединиц, молекулярной массе, ассоциации с разными вспомогательными белками, ускоряющими процесс биосинтеза ДНК, и функциональному назначению. ДНК-полимеразы α (альфа), β (бета), δ (дельта), ε (эпсилон) участвуют в синтезе ДНК в ядре клеток, ДНК-полимераза у (гамма) — в репликации митохондриальной ДНК.
Олигонуклеотид, синтезированный ДНК-полимеразой α и образующий небольшой двухцепочечный фрагмент с матрицей, позволяет присоединиться ДНК-полимеразе 5 и продолжить синтез новой цепи в направлении от 5'- к 3'- концу по ходу раскручивания репликативной вилки.
ДНК-полимераза 5 последовательно наращивает цепь, шаг за шагом присоединяя к ней соответствующие дезоксинуклеотиды. Выбор ДНК-полимеразой 5 очередного нуклеотида определяется матрицей. Включение дезоксирибонуклеозидмонофосфатов в растущую цепь ДНК сопровождается гидролизом макроэргических связей, соответствующих нуклеозидтрифосфатов и отщеплением пирофосфата (Н4Р2О7). Энергия макроэргических связей расходуется на образование 3',5'-фосфодиэфирной связи между последним нуклеотидом растущей цепи ДНК и присоединяемым нуклеотидом. Включение нуклеотида в синтезируемую цепь ДНК невозможно без предварительного связывания азотистого основания водородными связями с комплементарным нуклеотидом матричной цепи. ДНК-полимеразы (α, β, y, δ, ε) могут синтезировать нуклеотидную цепь только в направлении 5' —> 3', матричная цепь всегда считывается в направлении 3' —> 5'.
Фермент ДНК-лигаза катализирует образование фосфодиэфирной связи между 3'-ОН-группой дезоксирибозы одного фрагмента цепи ДНК и 5'-фосфатом следующего фрагмента. Реакция протекает с затратой энергии АТФ. Таким образом, из множества фрагментов Оказаки образуется непрерывная цепь ДНК.
В. Ориджины репликации
Рис. 4-17. Образование двух репликативных вилок, перемещающихся в противоположных направлениях от ориджина.

После завершения репликации происходит метилирование нуклеотидных остатков вновь образованных цепей ДНК. Метальные группы присоединяются ко всем остаткам аденина в последовательности -GATC-, при этом образуется N6-метиладенин, а также возможны метилирование цитозина в последовательности -GC- и образование N5-метилцитозина. Количество метилированных оснований равно примерно 1 — 8%. Модификация происходит при участии ферментов, использующих в качестве источника метальных групп S-аденозилметионин (SAM) (см. раздел 9). Присоединение метильных групп к остаткам аденина и цитозина не нарушает комплементарности цепей (рис. 4-18).
Рис. 4-18. Метилирование остатков аденина в последовательности -GАТС-. В течение нескольких минут после репликации, пока не произошло метилирование, новая цепь ДНК отличается от матричной цепи.

Наличие метальных групп в цепях ДНК необходимо для формирования структуры хромосом, а также для регуляции транскрипции генов. В течение непродолжительного времени в молекуле ДНК последовательности -GATC- метилированы по аденину только в матричной, но не в новой цепи. Это различие используется ферментами репарации для исправления ошибок, которые могут возникать при репликации.
Г. Строение 3' и 5'-концов цепей ДНК. Теломерная ДНК
На каждом конце хромосомы присутствует специфическая нуклеотидная последовательность. Она представлена многочисленными повторами (сотни или даже тысячи раз) олигонуклеотидов -GGGТТА-, называемых теломерной последовательностью, или просто теломерной ДНК. Наличие теломер необходимо для завершения репликации концевых информативных последовательностей хромосом, т. е. для сохранения генетической информации.
После завершения репликации хромосомы 5'-концы дочерних цепей ДНК недостроены, так как после удаления праймеров эти фрагменты оказываются недореплицированными. Это происходит потому, что ДНК-полимераза β, отвечающая за заполнение бреши, образованной после удаления праймера, не может вести синтез цепи ДНК от 3'- к 5'-концу (рис. 4-19, А). Таким образом, в ходе каждого цикла репликации 5'-концы синтезированных цепей укорачиваются. Но такие потери не представляют опасности для генетической информации хромосом, потому что укорочение ДНК идёт за счёт теломер. Во время следующего цикла репликации 5-концы цепей ДНК опять остаются недостроенными. Таким образом, с каждым клеточным делением ДНК хромосом будут последовательно укорачиваться. Укорочение теломер в большинстве клеток по мере их старения — важный фактор, определяющий продолжительность жизни организма.
Рис. 4-19. Синтез теломерной ДНК. А - на рисунке показано укорочение вновь синтезированных цепей ДНК после удаления праймеров; Б - в состав теломеразы входит короткая молекула РНК, содержащая в активном центре последовательность нуклеотидов, комплементарную теломерному повтору; 1 - фермент прикрепляется за счёт взаимодействия РНК с существующей теломерой и добавляет последовательно по одному нуклеотиду фрагмент -GGGТТА-. Матрицей служит простетическая группа теломеразы — фрагмент РНК; 2 - фермент перемещается по нити ДНК таким образом, что РНК-матрица в составе теломеразы постоянно комплементарно связана с концом вновь синтезированного теломерного повтора. Заново синтезированная теломерная ДНК служит матрицей для удлинения второй цепи ДНК, но уже в ходе следующего цикла клеточного деления. Теломерный повтор на рисунке взят в квадратные скобки -[GGGТТА]-.

Однако в эмбриональных и других быстро- делящихся клетках потери концов хромосом недопустимы, потому что укорочение ДНК будет происходить очень быстро. В эукариотических клетках имеется фермент теломераза (нуклеотидилтрансфераза), обеспечивающий восстановление недореплицированных 5'-концов. К особенностям этого фермента относят присутствие в качестве простетической группы РНК. Фрагмент РНК в активном центре теломеразы служит матрицей при синтезе теломерных повторов хромосом.
С помощью РНК фермент комплементарно прикрепляется к 3'-концу недостроенной дочерней цепи ДНК. Теломераза по принципу комплементарности последовательно удлиняет 3'-конец цепи ДНК на один гексануклеотид -GGGТТА-. Синтез всегда идёт от 5'- к 3'-концу. Затем теломераза смещается по цепи ДНК на один теломер и начинает синтез нового фрагмента -GGGТТА- (рис. 4-19, Б).
В большинстве соматических клеток теломераза неактивна, так как соматическая клетка имеет длину теломерной ДНК, достаточную для времени жизни клетки и её потомства. Однако небольшую активность теломеразы обнаруживают в клетках с высокой скоростью обновления, таких как лимфоциты, стволовые клетки костного мозга, клетки эпителия, эпидермиса кожи и др.
Д. Клеточный цикл и его регуляция
Рис. 4-20. Фазы клеточного цикла. После фазы М, в ходе которой происходит деление ядра (митоз) и цитоплазмы (цитокинез), дочерние клетки вступают в интерфазу нового цикла. Интерфаза начинается с фазы в ходе которой активно происходят биосинтетические процессы, резко замедленные во время митоза. Фаза S — период синтеза ДНК; она заканчивается, когда содержание ДНК в ядре удвоится и хромосомы полностью реплицируются. Затем наступает фаза G2, в ходе которой происходят деление митохондрий и увеличение энергетических запасов клетки. Фаза G2 продолжается до начала митоза, т. е. фазы М. В фазе М ядерная оболочка разрушается, формируются два новых ядра, цитоплазма делится с образованием двух дочерних клеток, имеющих по одному ядру. На рисунке представлен 24-часовой цикл.

Все фазы клеточного цикла G1, S, G2, М могут различаться по длительности, но в особенности это касается фазы G1, длительность которой может быть равна практически нулю или быть столь продолжительной, что может казаться, будто клетки вообще прекратили деление. В этом случае говорят, что клетки находятся в состоянии покоя (фаза G0). Так, нейроны взрослого человека не делятся вообще. Клетки эпителия кишечника делятся на протяжении всей жизни человека, но даже у этих быстропролиферирующих клеток подготовка к делению занимает 24 ч. Клетки лёгких, почек, печени во взрослом организме начинают делиться только лишь в ответ на повреждение органов.
Внешние сигналы могут стимулировать или ингибировать прохождение клетки через цикл. Пролиферативные сигналы очень разнообразны, они зависят от типа клетки, стадии развития и других факторов. Такими сигналами могут быть факторы роста, интерлейкины, гормоны, способные поддерживать или индуцировать пролиферацию определённых типов клеток. Сигнальные молекулы связываются специфическими мембранными рецепторами, активируют внутриклеточные пути передачи сигналов от рецептора к ядру и таким образом индуцируют транскрипцию определённых генов. Одними из первых активируются гены, кодирующие белки циклины. Белки были названы циклинами, потому что их концентрация в клетке периодически меняется по мере прохождения клеткой разных фаз клеточного цикла.
Таблица 4-2. Циклины и циклинзависимые киназы, регулирующие прохождение клеточного цикла
Читайте также: