
Маршанция жизненный цикл кратко
Обновлено: 30.06.2024
Мхи или моховидные – отдел высших растений, объеденных в классы: печёночные мхи (6 000–8 000 видов), антоцеротовые мхи (100–200 видов) и листостебельные мхи (10 000 видов). Мхи это, как правило, мелкие растения, длина которых редко превышает 50 мм. Отличаются от других высших растений тем, что в их жизненном цикле взрослые растения представлены разнополыми гаметофитами, а спорофит развивается на женском гаметофите. Размножаются бесполым путём – спорами и вегетативно частями таллома.
Цикл развития печёночного мха на примере маршанции изменчивой
В цикле развития взрослые (многолетние) растения маршанции полиморфной представлены разнополыми (двудомными) мужскими и женскими гаметофитами. Вегетативное тело растения – пластинчатый таллом, имеет вид мясистого лопастного, дихотомически разветвленного слоевища длиной до 10-12 см и шириной до 3 см, образованного множеством клеточных слоев (около 30). Специализированных проводящих тканей таллом не имеет. На нижнем эпидермисе образуются два типа выростов:
- ризоиды – бесцветные простые и язычковые одноклеточные выросты, которые выполняют функцию закрепления растения в субстрате. Воду маршанция способна поглощать всем телом,
Различить мужской и женский гаметофиты возможно во время развития на поверхности талломов специальных органов (подставок для гаметангиев) – мужских антеридиефоров и женских архегониефоров.
Антеридиефор представляет собой ножку с расположенным сверху восьмиугольным диском (шапкой). С верхней стороны шапки в антеридиальных камерах находятся мужские гаметангии – антеридии. В антеридиях образуются мужские двужгутиковые гаметы – сперматозоиды.
Архегониефоры развиваются на женских гаметофитах в виде девятилучевого щитка (звездочки) на ножке. Во время развития архегониефора, архегонии (женские гаметангии) закладываются с морфологически верхней стороны подставки. В силу неравномерного роста обеих сторон ложа подставки (щитка) они смещаются между лучами щитка на его нижнюю сторону, где располагаются группами. Вокруг каждой группы архегониев образуется специальная защитная обертка. Архегонии представляют собой кувшиноподобные камеры, направленные горловиной вниз, в брюшке которых образуется по одной яйцеклетке.
Половой процесс в цикле развития маршанции происходит посредством воды преимущественно во время дождя. Капли дождя выбивают из антеридиев зрелые сперматозоиды и вместе с ними отражаются на женские шапки архегониефоров. Сбегая между лучами женских подставок, капли воды вместе со сперматозоидами зависают в месте расположения архегониев, покрывая их. По воде мужские гаметы проникают через шейку архегония в брюшко – калиптру, где оплодотворяют яйцеклетку, образуя зиготу (первую диплоидную клетку спорофита). После оплодотворения вокруг каждого архегония из его ножки начинает образовываться индивидульная чашевидная защитная обертка – 4-5 лопастной псевдоперианций. В это время зигота делится митозом, формируя зародыш спорофита, который прикрепляется к материнскому телу (щитку архегониефора) присоской – гаусторией, и получает оттуда питание. С увеличением размеров зародыша калиптра (брюшко архегония) растягивается (увеличивается в размерах).
Взрослый спорофит, развивающийся из зародыша, включает в себя: стопу (присоску) – закрепляющую часть, ножку и спорангий (коробочку спорогона). В процессе своего роста спорофит разрывает калиптру и выносит спорангий наружу. В спорангии формируются диплоидные клетки спорогенной ткани, которые получают питание через удлиненные клетки со спирально утолщенными клеточными стенками – элатеры. В результате разделения спорогенных клеток мейозом образуются гаплоидные гетероталличные споры. При созревании спор элатеры высыхают и скручиваются спирально, разрыхляя массу спор в спорангии. Однослойная коробочка спорангия при созревании спор легко раскрывается восемью загнутыми назад зубцами, а пружинки элатер способствуют равномерному рассеиванию спор. Гетероталличные (разнополые) споры маршанции в благоприятных условиях прорастают в протонемы (передростки) – небольшие беспигментные нитчатые выросты, с верхушечной клетки которых образуются талломы соответственно мужского и женского гаметофита нового поколения.
Вегетативное размножение маршанции происходит с помощью выводковых почек, которые формируются в специальных выростах – выводковых корзинках, на верхней стороне как женских, так и мужских талломов. В результате попадания воды на таллом выводковые почки вымываются (распрыскиваются) и прорастают на почве в новые талломы соответствующего пола. Маршанция распространена в увлажненных местах на берегах водоемов, под покровом леса.
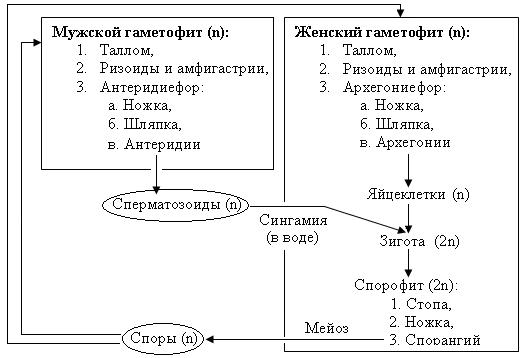
Рис. 30. Схема жизненного цикла мха маршанция
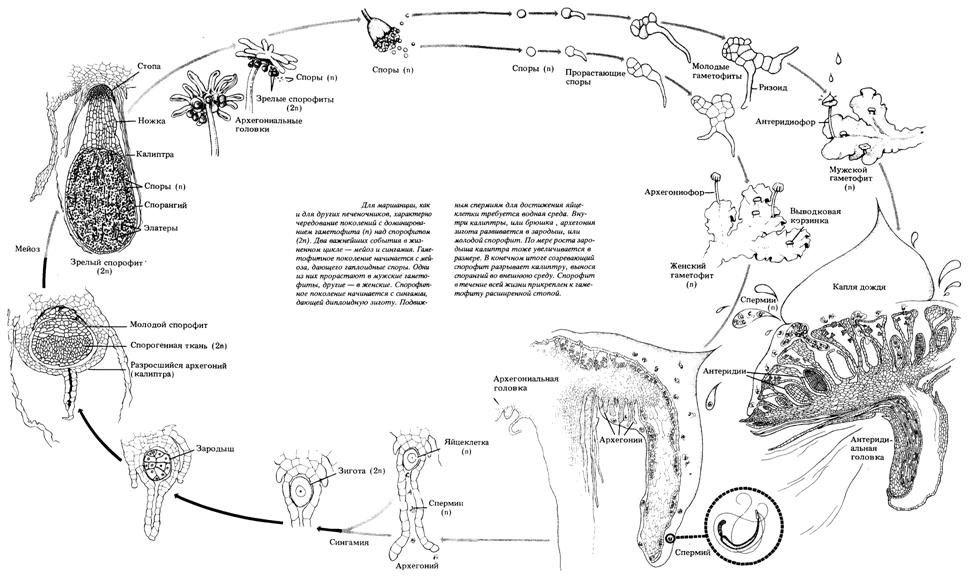
Рис. 31. Жизненный цикл мха маршанция
Цикл развития листостебельного мха на примере мха кукушкин лен
В цикле развития взрослые растения мха представлены раздельнополыми гаметофитами. Растения кукушкин лен имеют вертикальный стебель, длиной до 40 см, со спирально расположенными в три ряда редуцированными листьями, закрепленный в почве ризоидами. В центре стебля проходит концентрический проводящий тяж, среднюю часть которого занимают водопроводные клетки – гидроиды (восходящий поток), а периферийную – липтоиды – проводящие клетки органических соединений (нисходящий поток). На верхушке мужских растений находятся антеридии, в которых происходит формирование сперматозоидов, на верхушках женских – архегонии с яйцеклетками. Оплодотворение происходит с помощью капель воды. Выбивая из антеридиев сперматозоиды, капли дождя отражаются на архегонии соседних растений. По воде сперматозоиды проникают в брюшко архегониев, где оплодотворяют яйцеклетки. Из оплодотворенной яйцеклетки развивается спорофит, который своей стопой врастает в гаметофит. Ножка спорофита в процессе роста вытягивается в длину на 15-20 см, на верхушке которой образуется коробочка со спорангиями, закрытая крышкой. Край коробочки часто имеет зубчатый край, называемый перистомом. Вызревание спорофита длится до 6 месяцев (у других мхов до 18). В спорангиях спорофита происходит формирование спор из клеток спорогенной ткани, путем деления последних мейозом. После созревания спор крышка коробочки открывается, и споры рассеиваются через перистом. В благоприятных условиях споры прорастают в протонемы – нитевидные хлорофилльные выросты (таллом). Из пазушных почковидных структур протонем вырастают гаметофиты следующего поколения соответствующего пола.

Рис. 32. Схема жизненного цикла мха кукушкин лён
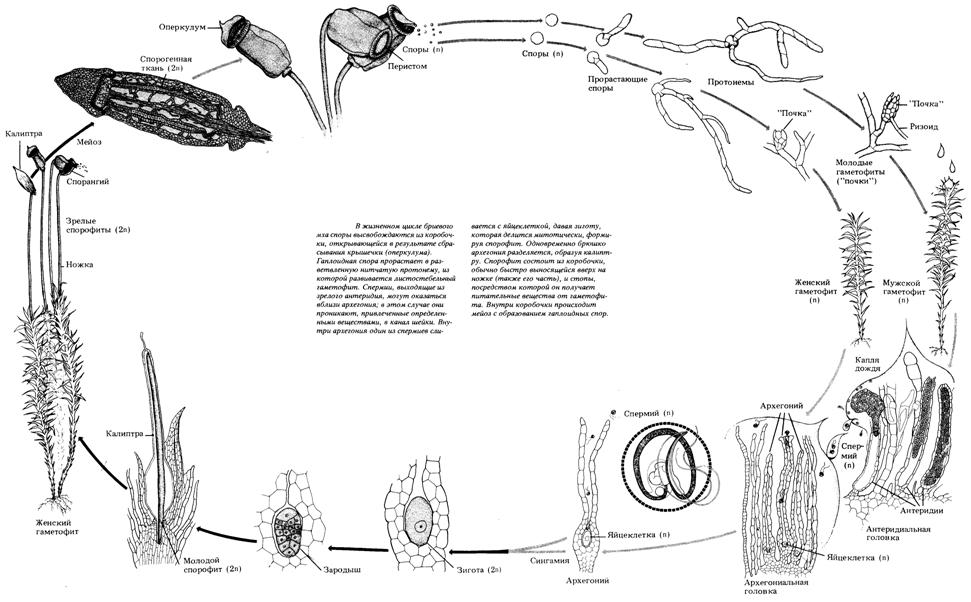
Рис. 33. Жизненный цикл мха кукушкин лён
Хвощевидные –отдел высших споровых растений, в настоящее время отнесён в подотдел Хвощёвые отдела Папоротниковидные и насчитывает 15 видов. Хвощёвые характеризуются наличием побегов, состоящих из чётко выраженных метамеров (междоузлий и узлов с мутовчатым расположением листьев). Спорофиты в анатомическом строении напоминают строение покрытосеменных злаковых растений. Бесполое размножение осуществляется разнополыми спорами, вегетативное – корневищами (подземными побегами).
Цикл развития хвощёвых на примере хвоща полевого
В цикле развития хвоща полевого преобладает спорофит – взрослое, многолетнее растение, состоящее из корневища, закрепленного в почве придаточными корнями. Весной из почек корневища на поверхность почвы вырастает спороносный, бесхлорофилльный вертикальный побег (стебель) с мутовчатым расположением редуцированных (маленьких) листьев, заканчивающийся спороносным колоском (стробилом). Сам спороносный колосок в своем строении имеет ось, на которой размещены зонтиковидные спороносные листья (щитки на ножках) – спорофиллы. С нижней стороны спорофиллов, обращенной к оси стробила, находится от 5 до 10 спорангиев. В спорангиях в результате редукционного деления мейозом клеток спорогенной ткани (2n) образуются гаплоидные, морфологически одинаковые, но разнополые споры (мужские и обоеполые). Оболочки спор имеют специальные выросты – элатеры, которые скручены спирально вокруг спор во влажном состоянии, а при подсыхании разворачиваются. Это позволяет спорам цепляться друг за друга и распространяться группами. После созревания спор спороносные листья стробила размыкаются, спорангии лопаются и споры разносятся ветром. Вследствие содержания в спорах хлоропластов они быстро (в течение 3-х недель) теряют всхожесть. Попадая на увлажненный илистый грунт, группы спор прорастают в хлорофиллоносные гаметофиты в виде лопастных пластинок, закрепляющихся в субстрате ризоидами. Гаметофиты достигают половой зрелости через 3-5 недель после прорастания. На мужских гаметофитах, меньшего размера, образуются антеридии – мужские гаметангии, в которых образуются многожгутиковые сперматозоиды. На двуполых гаметофитах, более рассеченной формы, архегонии (женские гаметангии) развиваются раньше антеридиев, что увеличивает вероятность перекрестного оплодотворения. Для достижения сперматозоидами яйцеклеток, находящихся в архегониях, необходима вода. На одном гаметофите могут быть оплодотворены сразу несколько яйцеклеток, из которых в дальнейшем развиваются зародыши, есть молодые спорофиты. Зародыши крепятся стопой к животику архегония и получают из гаметофита необходимые питательные вещества для развития, формируя зачаточный корешок, стебель и почку. После формирования, зародышевый корешок начинает расти, закрепляется в почве и молодой спорофит отсоединяется от гаметофита, который через некоторое время отмирает. После спороношения весенние (спороносные) побеги отмирают и из почек корневища вырастают зеленые ассимиляционные побеги. Ассимиляционные побеги имеют вертикальный стебель с мутовчатым расположением на нем боковых ветвей и листьев под ними. Они выполняют функцию образования органических соединений в процессе фотосинтеза и накопления их в корневище. В конце вегетационного периода ассимиляционные побеги отмирают, остается корневище, которое зимует в почве.
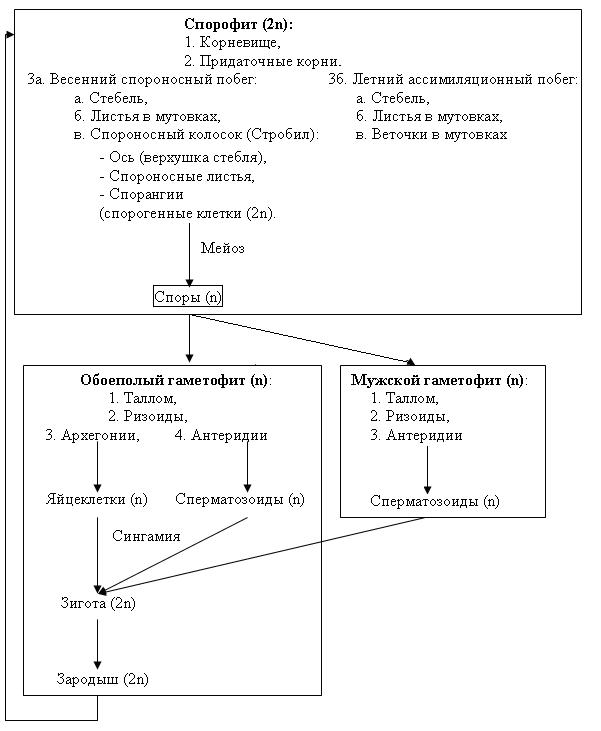
Рис. 34. Схема жизненного цикла хвоща полевого

Рис. 35. Жизненный цикл хвоща полевого
Плауновидные –отдел высших споровых растений, представлен 1200 видами. Характеризуются наличием развитой проводящей системы, корнепобеговым типом строения спорофитов и талломным строением гаметофитов, бесполым размножением спорами и вегетативным размножением надземными побегами.
Цикл развития плауновидных растений на примере плауна булавовидного
Спорофит плауна булавовидного – многолетнее вечнозеленое растение, преобладает в цикле развития. В своем составе имеет стелющийся дихотомически разветвленный стебель, покрытый спирально расположенными маленькими ланцетно-линейными листочками (микрофиллами) и закрепленный в почве дихотомически разветвленными корнями. Стеблевые побеги заканчиваются верхушечными почками или спороносными колосками (стробилами). Спороносные колоски на длинных ножках состоят из оси, на которой спирально расположены спороносные листья (спорофиллы) со спорангиями с верхней (дорсальной) стороны. После редукционного (мейотического) деления материнских диплоидных клеток образуются гаплоидные споры. Споры покрыты двойной оболочкой (эндо- и экзоспорием) и содержат до 50% масла. После созревания споры рассеиваются и в благоприятных условиях прорастают. Развитие гаметофита (заростка) из спор происходит медленно в течение 12-20 лет. Он образует ризоиды, которыми закрепляется в почве и поглощает воду, а также вступает в симбиоз с мицелием гриба, который размещается в его коровой части. Заросток развивается в почве без доступа света, поэтому не имеет хлоропластов, однако если он выходит на поверхность, они образуются. Гаметофит двуполый и напоминает луковицу, позже разрастается до 2-3 см в диаметре, и имеет блюдцеобразную форму. Антеридии и архегонии размещены рядом на верхней стороне и погружены в паренхимную ткань. Антеридии овальной формы, архегонии – колбообразные. В брюшной части архегониев содержится яйцеклетка и брюшная канальцевая клетка, в шейке – шейные канальцевые клетки. Для перемещения сперматозоидов с антеридиев к яйцеклеткам архегониев необходима вода. Заплывая в шейку архегония, сперматозоид движется в брюшко архегония, где сливается с яйцеклеткой, образуя зиготу – первую диплоидную клетку спорофита. Зародыш – молодой спорофит, медленно развивается на гаметофите, получая из него необходимые питательные вещества. Когда корни зародыша прорастают в почву, он отсоединяется от гаметофита и разрастается во взрослое растение. При постепенном созревании архегониев на гаметофите в них могут образовываться и развиваться несколько разновозрастных спорофитов одновременно.
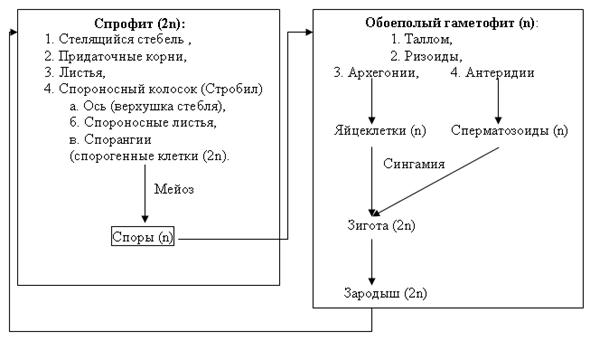
Рис. 36. Схема жизненного цикла плауна булавовидного
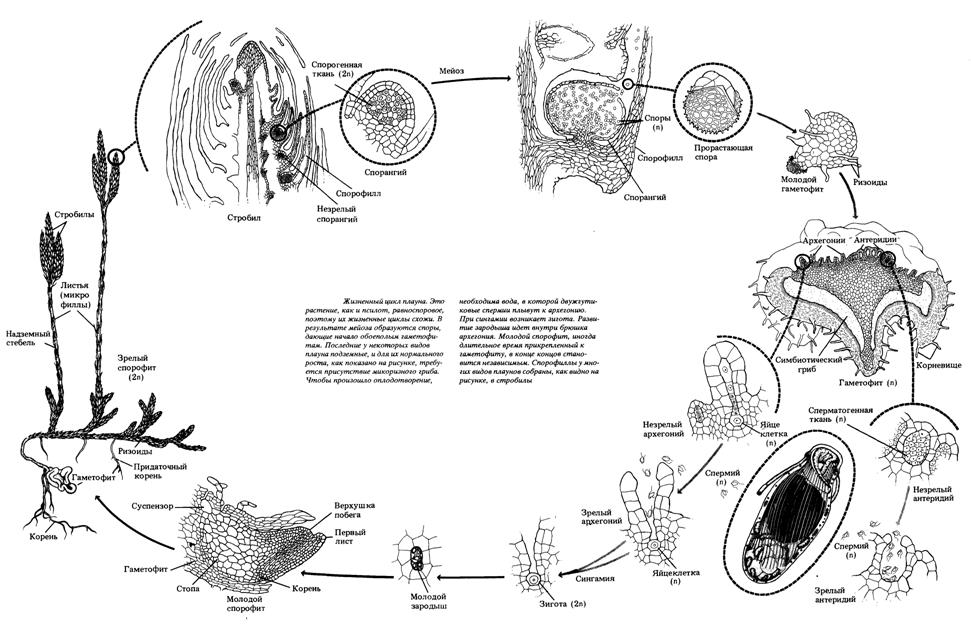
Рис. 37. Жизненный цикл плауна булавовидного
Папоротникообразные – отдел высших споровых растений, объединяющий около 11 000 видов. Папоротникообразные растения характеризуются корнепобеговым строением тела спорофитов и талломным строением тела гаметофитов. Размножаются бесполым путём – спорами и вегетативно – побегами (корневищами).
Маршанция – растение двудомное. При половом размножении на мужских талломах(рис.2 -1) развиваются выросты в виде многолопастного диска, расположенного на ножке. Это мужские антеридиальные подставки(рис.2-3). В верхней части диска находятся антеридиальные полости(рис.2-4), на дне которых находится по 1 антеридию. Антеридий(рис.2 -5)– овальное тельце с однослойной стенкой, прикрепляется ко дну полости небольшой ножкой(рис.2 -5н). Его содержимое состоит из многочисленных спермагенных клеток, в каждой из которых образуют 2 двужгутиковых сперматозоида(рис.2 -8). При созревании антеридий вскрывается наверху щелью, сперматозоиды во время дождя через канал антеридиальной полости выходят наружу и подплывают к архегонию.
На женских талломах(рис.2 -2) образуются выросты, имеющие ножку, увенчанную многолучевой звездой. Это женские, архегониальные подставки(рис.2 -6). На них находятся архегонии, сидящие группами на лучах и обращенные шейками вниз.
Каждая группа архегониев покрыта общим покровом – перихецием(рис.2 -7б), а каждый архегоний одет частным покрывалом – перианцием(рис.2 -7а), в виде кольцевидного валика у его основания. Перианций затем сильно разрастается. Перихеций и перианций защищают архегонии, а затем и спорогоны от высыхания.
Архегонии(рис.2 -7арх)– небольшие колбообразные тельца, состоят из расширенной части –брюшка и суженной –шейки. Снаружи архегоний(рис.2 -9) окружен бесплодными клетками =стенкой архегония,защищающими от высыхания содержимое. В брюшке находится неподвижная женская гамета – яйцеклетка, над которой расположена брюшная канальевая клетка. Внутри шейки расположены шейковые канальцевые клетки. К моменту созревания яйцеклетки канальцевые клетки и стенки архегония, а также верхние клетки стенки ослизняются, и архегоний на верхушке вскрывается. От его вершины к яйцеклетке ведет канал, заполненный слизью, образовавшейся из расплывшихся клеток. По этой слизи, сперматозоид проникает к яйцеклетке, производя оплодотворение. По мере созревания лучи женской подставки поднимаются, принимая сначала горизонтальное положение, а ко времени созревания спорогонов загибаются кверху. Ткань подставки имеет строение таллома, а ножка состоит из паренхимных клеток. Вдоль ножки (внутри её) протягиваются два тяжа язычковых ризоидов – от нижней стороны таллома до диска или до лучей подставки. По мере развития подставки ножка усиленно растет и выносит антеридии и архегонии высоко над талломом.
Из оплодотворенной яйцеклетки развивается бесполое поколение маршанции –спорофит или спорогон(рис.2 -10). Оплодотворенная яйцеклетка тотчас же делится поперечной перегородкой на две клетки. Из нижней клетки благодаря последующим делениям формируется ножка спорогона(рис.2-10н). Из верхней клетки развивается коробочка(рис.2 -10к). Верхняя клетка делится двумя взаимно перпендикулярными перегородками на 4 клетки, а последние, благодаря делению в тангентальном направлении, отчленяют наружные клетки, создающие стенки коробочки, и внутренние, образующие археспорий.
Археспориальные клетки делятся многократно и образуют материнские клетки спор. Одна часть материнских клеток спор приступает к делению. Первое их деление редукционное. Образовавшиеся две клетки делятся уже митотически ещё раз, и в результате образуется тетрада (четверка) спор. Таким образом, споры гаплоидны, так как образовались в результате редукционного деления, которое у всех высших растений всегда происходит при образовании спор. Споры одеты двумя оболочками – наружной–экзоспорием и внутренней, тонкой –эндоспорием. Эндоспорий облегает внутреннее содержимое споры, состоящее из плазмы и ядра.
Другая часть материнских клеток спор вытягивается и благодаря неравномерному утолщению оболочки спирально закручиваются. Каждая такая клетка превращается в пружинку – элатеру.
Итак, спорогон состоит из шаровидной коробочки(рис.2 -10к,11кор), короткой ножки, расширенная нижняя часть которой называется гаусторией(рис.2 -10г). При помощи гаустории спорогон внедряется в ткань подставки (гаметофита) и получает из нее питательные вещества. Коробочка содержит споры (рис.2 -12сп) и пружинки – элатеры(рис.2 -12 эл). Пружинки представляют собой удлиненные мертвые клетки, тонкие стенки которых имеют спиральные утолщения в виде одной или двух спиралей, идущих навстречу друг другу. Функция зрелых элатер, подвижных вследствие гигроскопичности спиральных утолщений, сводится к выбрасыванию спор из коробочки или разрыхлению их в ней.

Первоначально спорогон находится в брюшке архегония(рис.2 -10бр), стенки которого разрастаются и образуют колпачок(рис.2 -11к). Снаружи спорогон окружен также сильно разросшимся перианцием(рис.2 -10п,11п). При созревании коробочки ножка спорогона удлиняется, в результате коробочка разрывается на верхушке и выносится ножкой за пределы лучей подставки. Коробочка вскрывается створками, после чего споры высыпаются наружу. Пружинки – элатеры скручиваются в сухую погоду и раскручиваются в сырую, благодаря этому споровая масса разрыхляется и равномерно рассеивается.
При прорастании споры наружная оболочка – экзоспорий лопается, а содержимое, окруженное эндоспорием, вытягивается в короткую нить. На конце нити отчленяется клетка, которая делится в разных направлениях. В результате формируется небольшая пластинчатая протонема(рис.2 -13,14), далее развивающаяся в таллом маршанции.
Таким образом, у маршанции имеется смена поколений (генераций): половое поколение (гаметофит) сменяется бесполым (спорофитом) и т.д. В цикле развития преобладает гаметофит – растение маршанции с мужскими и женскими половыми органами. Все его клетки гаплоидны, т. к. берут начало от гаплоидной споры. Спорофит (спорогон) имеет небольшие размеры и питается за счет гаметофита. Все его клетки диплоидны, т.к. развиваются из диплоидной зиготы. Переход от спорофита к гаметофиту осуществляется за счет редукционного деления.
В народной медицине спиртовую настойку маршанции обыкновенной применяют при болезнях печени. Лекарственные свойства печёночныхмхов практически мало изучены, они ждут своего исследователя.
ПЕЛЛИЯ ЭПИФИЛЛА.
(Рисунок 3 – Пеллия эпифилла)
Отдел – МОХООБРАЗНЫЕ (Bryophyta)
Класс – ПЕЧЁНОЧНИКИ или ПЕЧЁНОЧНЫЕ МХИ (Marchantiopsida или Hepaticopsida)
Подкласс – ЮНГЕРМАННИЕВЫЕ(Jungermanniidae)
Порядок – МЕТЦГЕРИЕВЫЕ (Metzgeriales)
Семейство –Пеллиевые (Pelliaceae)
Род – Пеллия (Pellia)
Вид – Пеллия эпифилла (Pellia epiphylla)
Пеллия эпифилла широко распространена у нас по сырым лесным оврагам, около ручьёв и канав. Тонкие лентовидные узкие талломы пеллии(рис.3 -1) дихотомически ветвятся. Они светло–зелёные, в центре многослойные, по краям однослойные. Таллом имеет очень простое анатомическое строение. Он одет верхним и нижним эпидермисами и состоит из немногих слоев паренхимных клеток. От клеток нижнего эпидермиса, нерезко отграниченного от основной ткани таллома, отходят простые ризоиды(рис.3 -1а). Воздушных камер с ассимиляторами нет, а хлорофилловые зерна имеются во всех клетках таллома.
(Рисунок 2 – Маршанция обыкновенная – цикл развития)
Маршанция – растение двудомное. При половом размножении на мужских талломах(рис.2 -1) развиваются выросты в виде многолопастного диска, расположенного на ножке. Это мужские антеридиальные подставки(рис.2-3). В верхней части диска находятся антеридиальные полости(рис.2-4), на дне которых находится по 1 антеридию. Антеридий(рис.2 -5)– овальное тельце с однослойной стенкой, прикрепляется ко дну полости небольшой ножкой(рис.2 -5н). Его содержимое состоит из многочисленных спермагенных клеток, в каждой из которых образуют 2 двужгутиковых сперматозоида(рис.2 -8). При созревании антеридий вскрывается наверху щелью, сперматозоиды во время дождя через канал антеридиальной полости выходят наружу и подплывают к архегонию.
На женских талломах(рис.2 -2) образуются выросты, имеющие ножку, увенчанную многолучевой звездой. Это женские, архегониальные подставки(рис.2 -6). На них находятся архегонии, сидящие группами на лучах и обращенные шейками вниз.
Каждая группа архегониев покрыта общим покровом – перихецием(рис.2 -7б), а каждый архегоний одет частным покрывалом – перианцием(рис.2 -7а), в виде кольцевидного валика у его основания. Перианций затем сильно разрастается. Перихеций и перианций защищают архегонии, а затем и спорогоны от высыхания.
Архегонии(рис.2 -7арх)– небольшие колбообразные тельца, состоят из расширенной части –брюшка и суженной –шейки. Снаружи архегоний(рис.2 -9) окружен бесплодными клетками =стенкой архегония,защищающими от высыхания содержимое. В брюшке находится неподвижная женская гамета – яйцеклетка, над которой расположена брюшная канальевая клетка. Внутри шейки расположены шейковые канальцевые клетки. К моменту созревания яйцеклетки канальцевые клетки и стенки архегония, а также верхние клетки стенки ослизняются, и архегоний на верхушке вскрывается. От его вершины к яйцеклетке ведет канал, заполненный слизью, образовавшейся из расплывшихся клеток. По этой слизи, сперматозоид проникает к яйцеклетке, производя оплодотворение. По мере созревания лучи женской подставки поднимаются, принимая сначала горизонтальное положение, а ко времени созревания спорогонов загибаются кверху. Ткань подставки имеет строение таллома, а ножка состоит из паренхимных клеток. Вдоль ножки (внутри её) протягиваются два тяжа язычковых ризоидов – от нижней стороны таллома до диска или до лучей подставки. По мере развития подставки ножка усиленно растет и выносит антеридии и архегонии высоко над талломом.
Из оплодотворенной яйцеклетки развивается бесполое поколение маршанции –спорофит или спорогон(рис.2 -10). Оплодотворенная яйцеклетка тотчас же делится поперечной перегородкой на две клетки. Из нижней клетки благодаря последующим делениям формируется ножка спорогона(рис.2-10н). Из верхней клетки развивается коробочка(рис.2 -10к). Верхняя клетка делится двумя взаимно перпендикулярными перегородками на 4 клетки, а последние, благодаря делению в тангентальном направлении, отчленяют наружные клетки, создающие стенки коробочки, и внутренние, образующие археспорий.
Археспориальные клетки делятся многократно и образуют материнские клетки спор. Одна часть материнских клеток спор приступает к делению. Первое их деление редукционное. Образовавшиеся две клетки делятся уже митотически ещё раз, и в результате образуется тетрада (четверка) спор. Таким образом, споры гаплоидны, так как образовались в результате редукционного деления, которое у всех высших растений всегда происходит при образовании спор. Споры одеты двумя оболочками – наружной–экзоспорием и внутренней, тонкой –эндоспорием. Эндоспорий облегает внутреннее содержимое споры, состоящее из плазмы и ядра.
Другая часть материнских клеток спор вытягивается и благодаря неравномерному утолщению оболочки спирально закручиваются. Каждая такая клетка превращается в пружинку – элатеру.
Итак, спорогон состоит из шаровидной коробочки(рис.2 -10к,11кор), короткой ножки, расширенная нижняя часть которой называется гаусторией(рис.2 -10г). При помощи гаустории спорогон внедряется в ткань подставки (гаметофита) и получает из нее питательные вещества. Коробочка содержит споры (рис.2 -12сп) и пружинки – элатеры(рис.2 -12 эл). Пружинки представляют собой удлиненные мертвые клетки, тонкие стенки которых имеют спиральные утолщения в виде одной или двух спиралей, идущих навстречу друг другу. Функция зрелых элатер, подвижных вследствие гигроскопичности спиральных утолщений, сводится к выбрасыванию спор из коробочки или разрыхлению их в ней.
Первоначально спорогон находится в брюшке архегония(рис.2 -10бр), стенки которого разрастаются и образуют колпачок(рис.2 -11к). Снаружи спорогон окружен также сильно разросшимся перианцием(рис.2 -10п,11п). При созревании коробочки ножка спорогона удлиняется, в результате коробочка разрывается на верхушке и выносится ножкой за пределы лучей подставки. Коробочка вскрывается створками, после чего споры высыпаются наружу. Пружинки – элатеры скручиваются в сухую погоду и раскручиваются в сырую, благодаря этому споровая масса разрыхляется и равномерно рассеивается.
При прорастании споры наружная оболочка – экзоспорий лопается, а содержимое, окруженное эндоспорием, вытягивается в короткую нить. На конце нити отчленяется клетка, которая делится в разных направлениях. В результате формируется небольшая пластинчатая протонема(рис.2 -13,14), далее развивающаяся в таллом маршанции.
Таким образом, у маршанции имеется смена поколений (генераций): половое поколение (гаметофит) сменяется бесполым (спорофитом) и т.д. В цикле развития преобладает гаметофит – растение маршанции с мужскими и женскими половыми органами. Все его клетки гаплоидны, т. к. берут начало от гаплоидной споры. Спорофит (спорогон) имеет небольшие размеры и питается за счет гаметофита. Все его клетки диплоидны, т.к. развиваются из диплоидной зиготы. Переход от спорофита к гаметофиту осуществляется за счет редукционного деления.
В народной медицине спиртовую настойку маршанции обыкновенной применяют при болезнях печени. Лекарственные свойства печёночныхмхов практически мало изучены, они ждут своего исследователя.
ПЕЛЛИЯ ЭПИФИЛЛА.
(Рисунок 3 – Пеллия эпифилла)
Отдел – МОХООБРАЗНЫЕ (Bryophyta)
Класс – ПЕЧЁНОЧНИКИ или ПЕЧЁНОЧНЫЕ МХИ (Marchantiopsida или Hepaticopsida)
Подкласс – ЮНГЕРМАННИЕВЫЕ(Jungermanniidae)
Порядок – МЕТЦГЕРИЕВЫЕ (Metzgeriales)
Семейство –Пеллиевые (Pelliaceae)
Род – Пеллия (Pellia)
Вид – Пеллия эпифилла (Pellia epiphylla)
Пеллия эпифилла широко распространена у нас по сырым лесным оврагам, около ручьёв и канав. Тонкие лентовидные узкие талломы пеллии(рис.3 -1) дихотомически ветвятся. Они светло–зелёные, в центре многослойные, по краям однослойные. Таллом имеет очень простое анатомическое строение. Он одет верхним и нижним эпидермисами и состоит из немногих слоев паренхимных клеток. От клеток нижнего эпидермиса, нерезко отграниченного от основной ткани таллома, отходят простые ризоиды(рис.3 -1а). Воздушных камер с ассимиляторами нет, а хлорофилловые зерна имеются во всех клетках таллома.


На фото — маршанция изменчивая (Marchantia polymorpha), разросшаяся почти сплошным ковром на гари спустя несколько лет после лесного пожара. Это очень распространенный вид-космополит, заселяющий нарушенные местообитания вроде пожарищ или кострищ, обочин и выбоин дорог, любых обнажений почвы. Для успешного развития маршанции нужно одновременно и достаточно высокое содержание минеральных веществ в субстрате, и низкое число конкурентов. Растения с такой экологической стратегией называются рудералами.
Как вы видите, выглядит маршанция довольно замысловато. Но при этом у нее нет разделения тела на органы. Такой вариант организации тела растения называют талломом, или слоевищем. С нижней стороны таллома есть одноклеточные ризоиды. Часть из них прорастает в субстрат и, по-видимому, служит для закрепления таллома в субстарте и для минерального питания. Часть ризоидов ориентирована параллельно субстрату и прикрыта брюшными чешуйками. Эти ризоиды, отличающиеся внутренними утолщениями клеточной стенки, называют язычковыми. Есть предположение, что они работают как аналог проводящей системы и позволяют воде распространяться по слоевищу.
У всех мохообразных в жизненном цикле преобладает гаплоидная стадия — гаметофит. На нем образуются половые клетки, а после оплодотворения там же, на гаметофите, развивается диплоидное поколение — спорофит. У каждого гаметофита есть пол, мужской или женский, который определяется половой хромосомой, Y или X. Выросты, похожие на миниатюрные пальмы, — это так называемые подставки. На подставках образуются мужские гаметангии — антеридии, и женские — архегонии. В антеридиях развиваются сперматозоиды, в архегониях — яйцеклетки. Отличить мужскую подставку от женской легко: мужская подставка более плоская, более округлая (на верхнем фото мужские подставки имеют фиолетовый оттенок). Антеридии расположены с ее верхней стороны. Выход сперматозоидов происходит при попадании воды на мужскую подставку.
Скрещивание маршанции изменчивой в лабораторных условиях. На мужские подставки капают жидкость, антеридии открываются, и из них выходит мутная масса, содержащая сперматозоиды
Женская подставка разделена на 9–12 узких долей, загибающихся вниз. Архегонии закладываются сверху, но в ходе роста и растяжения клеток оказываются с нижней стороны, на которой развиваются многочисленные чешуйки. Женская подставка такой формы неплохо удерживает каплю воды. На ножках подставок есть ризоиды: ножка по строению соответствует нижней стороне слоевища. Для оплодотворения маршанции нужна пленка воды, по которой сперматозоиды могут доплыть до архегониев с яйцеклеткой. Вероятно, ризоиды и чешуйки с нижней стороны женской подставки способствуют образованию и сохранению этой водной пленки.

Женские подставки маршанции изменчивой. На их нижней стороне видны чешуйки, на ножке — нитевидные ризоиды. Фото © Евгения Правдолюбова, Ямало-Ненецкий округ, июль 2018 года
После оплодотворения на нижней стороне женских подставок развивается диплоидное поколение — спорофит. Спорофит состоит из коробочки, ножки и стопы — гаусториальных клеток, которые обеспечивают поглощение питательных веществ из тканей гаметофита. В коробочке происходит образование спор и специальных разрыхляющих споровую массу клеток — элатер (см. также Elater). Ножка спорофита у маршанции очень короткая, и коробочка долгое время остается скрытой в тканях материнского организма. При созревании коробочки ножка удлиняется, коробочки оказываются снаружи, раскрываются и рассеивают гаплоидные споры. Одна подставка маршанции производит около 7 миллионов спор.

Прорастание спор светозависимое и по крайней мере в лабораторных условиях требует длинного светового дня. Споры разбухают и начинают синтезировать хлорофилл, потом прорастают. Первые этапы развития известны с точностью до клетки: сначала образуется первый ризоид, потом сферическая протонема из небольшого числа клеток.
Таллом маршанции образует особые структуры для вегетативного размножения — круглые корзиночки с выводковыми тельцами. Корзиночки появляются за счет удлинения клеток эпидермы. После этого другие клетки эпидермы на дне корзиночки начинают делиться и образуют выводковые тельца — маленькие многоклеточные образования с одной или несколькими точками роста. Выводковые тельца разлетаются с каплями дождя, когда капли попадают в корзиночку.
У маршанции изменчивой подставки и корзиночки с выводковыми тельцами могут встречаться на одном талломе в одно и то же время. В лабораторных условиях можно стимулировать появление подставок длинным световым днем и дальним красным светом, а образование корзиночек — коротким световым днем, то есть, как этого и следует ожидать, фазы бесполого и полового размножения запускаются разными факторами среды. Что приводит к одновременному появлению подставок и корзиночек в природе, пока не установлено.

Маршанция, выросшая в цветочном горшке. На поверхности слоевища видны поры и корзиночки с выводковыми тельцами. Фото © Евгения Правдолюбова, Рязанская область, июль 2014 года
В талломе маршанции есть несколько типов тканей: покровная ткань — эпидерма; нефотосинтезирующая основная ткань, некоторые клетки которой несут запасы питательных веществ — зерна крахмала или масляные тельца; и способная к фотосинтезу ассимиляционная ткань. Масляные тельца, образующиеся из фрагментов эндоплазматической сети, — это уникальная органелла, известная только у печеночников. В них содержится смесь гидрофобных веществ разного состава, в основном терпеноиды, а также ароматические вещества, ненасыщенные жирные кислоты и триглицериды. Некоторые из этих веществ в экспериментах проявляют цитотоксичность по отношению к клеткам животных, антибактериальную и антигрибную активность. В масляных тельцах маршанции также обнаружены белки, родственные ферментам синтеза изопренов. Значение масляных телец точно не установлено, но можно предположить, что эти вещества могут отпугивать животных, желающих полакомиться сочным талломом, защищать талломы от ультрафиолетового излучения, подавлять развитие патогенных грибов.

Ассимиляционная ткань образует воздушные камеры, которые сообщаются с атмосферой через поры, одна пора на одну камеру. Они способствуют газообмену и благодаря водоотталкивающей кутикуле не позволяют каплям воды проникнуть в воздушные камеры. Иногда эти поры называют устьицами, но это не совсем корректно: несмотря на общую функцию, строение пор маршанции совсем непохоже на строение устьиц мхов из других отделов и на устьица сосудистых растений. Эти поры, за редкими исключениями, не способны менять свой диаметр при обезвоживании таллома (исключение — печеночники рода Preissia с особым строением пор). Удивительно и то, что они образуются у гаплоидного поколения, у всех остальных высших растений устьица есть только на спорофитах.

Схема среза воздушной камеры маршанции. Бочонкообразная структура — пора. Она состоит из 4–7 расположенных друг над другом колец, каждое из 4–5 клеток. Рисунок с сайта elte.prompt.hu
Хотя у маршанции нет корней, она может образовывать микоризоподобные симбиозы с гломусовыми грибами. Контакт клеток гриба и печеночника происходит внутри таллома. Для маршанции изменчивой образование такого симбиоза не очень характерно и отмечено только для одного подвида — Marchantia polymorpha subsp. montivagans, который растет на более бедной почве, чем остальные подвиды. Несколько видов грибов-базидиомицетов специализировались на питании маршанцией. Эти грибы относят к роду Loreleia. Точно не установлено, являются ли они сапротрофами или паразитами.

Loreleia postii — один из видов грибов, растущих на маршанциях. Точно не установлено, является ли он сапротрофом на отмерших частях таллома, или паразитом на живых тканях маршанции. Фото © Евгения Правдолюбова, Ямало-Ненецкий округ, июль 2018 года
Рудеральные растения, к которым относится маршанция, обычно легко культивировать: воссоздание нарушенных местообитаний требует минимум усилий. И как раз рудеральные виды зачастую становятся модельными объектами биологов. Арабидопсис, или резуховидка Таля, знаменитое цветковое растение, на котором изучены основные закономерности физиологии растений, — сорняк, как и маршанция. Первые лабораторные эксперименты с маршанцией датируются XIX веком. На ней проследили развитие выводковых телец и оплодотворение. В эпоху молекулярно-генетических исследований маршанция изменчивая оказалась одним из первых растений, у которых отсеквенировали геном митохондрий и пластид. Для маршанции разработаны протоколы выращивания и генетической трансформации. Недавно была установлена и проанализирована полная последовательность генома маршанции изменчивой.
Маршанция обыкновенная растет по болотам, в увлажненных местах. Лентовидное тело дихотомически ветвится, стелется по земле и достигает 5–20 см в длину и 1–2 см в ширину. Так как тело маршанции имеет очень простое строение, его называют слоевищем или талломом (рис. 1-II-1).
Верхняя сторона таллома темно–зеленого цвета. На нем заметна "средняя жилка", идущая вдоль таллома. Верхняя поверхность таллома имеет сетчатую структуру и разбита на участки – воздушные камеры(рис.1-I-1к)с темными точками в каждом и одной светлой. Темные точки –ассимиляторы(рис.1-I-1а)– ветвистые нити, составленные из округлых клеток, содержащих хлорофилловые зерна Светлые точки –устьица (рис.1-I-1у), (рис.1-I-2)
Нижняя сторона таллома буровато-красная. Вдоль средней жилки располагаются одноклеточные ризоиды (рис.1-I-3р), которые отходят в землю, многие из них отходят и в стороны от средней жилки таллома, но там они плотно к нему прижаты.
Анатомическое строение таллома таково. На поперечном срезе таллома (рис.1-I-5) видны верхний эпидермис (рис.1-I-5эп) и устьица (рис.1-I-5у). Устьица имеют бочонкообразную форму и состоят из 4-х рядов клеток (16 клеток), расположенных в 4 этажа по 4.
Под эпидермисом – воздушные камеры,ограниченные бесцветными крупными клетками, расположенными в 1–2 этаж (рис.1-I-5ст.к.- стенка камеры). Со дна воздушных камер поднимаются ветвистые нити ассимиляторы (рис.1-I-5ас), составленные округлыми клетками, содержащими хлорофилловые зерна. Это главный ассимиляционный аппарат маршанции, обусловливающий темно-зеленую окраску верхней стороны ее таллома. Ткань под воздушными камерами заполнена широкими бесцветными паренхимными клетками (рис.1-I-5пар), которые нередко заполнены крахмалом. Местами встречаются клетки, содержащие включения – масляные тельца. Изредка попадаются полости (ходы), образовавшиеся от разрушения соседних клеток. Это является особенностью всех печеночников.
Нижний эпидермис (рис.1-I-5н.эп.) ограничен от паренхимной ткани. От него отходят амфигастрии– особые выросты таллома, представленные однорядными пластиночками. Амфигастрии еще называют однослойными брюшными чешуйками (рис.1-I-3а). Амфигастрии тянутся в 2 ряда вдоль средней жилки и краев таллома, прижимая к таллому тяжи язычковых ризоидов.
Вдоль средней жилки располагаются многочисленные ризоиды 2–х типов:
1) простые ризоиды (рис.1-I-4а)– имеют широкий просвет, внутренние стенки гладкие, располагаются вдоль средней жилки. Растут вертикально вниз. Врастают в землю, всасывают воду с растворенными минеральными веществами и прикрепляют таллом маршанции к почве.
2) язычковые ризоиды (рис.1-I-4б,в) – имеют на внутренней поверхности выросты в виде сосочков – язычков. Эти ризоиды сплетаются в тяжи, которые протягиваются вдоль краев таллома и прижимаются к нему амфигастриями. По язычковым ризоидам, как по фитилю, происходит движение воды вдоль таллома.
Внутри таллома маршанции нет проводящей ткани, прохождение воды обеспечивается наружной проводящей системой в виде язычковых ризоидов.
Вегетативное размножение
Осуществляется за счет всех частей слоевища, а также специальными выводковыми почками, которые развиваются в выводковых корзиночках на верхней стороне таллома (рис.1-II-1). В такой корзиночке можно видеть большое количество почек, находящихся на разных стадиях развития (рис.1-II-2,3).Сформировавшаяся выводковая почка (рис.1-II-4, 5-поперечный разрез) имеет вид овальной пластиночки с выемками по бокам, где помещаются точки роста. От почки отходят ризоиды (рис.1-II-4р,5р). Выводковые почки расположены на 1-клеточной ножке (рис.1-II-3н), от которой легко отламываются (рис.1-II-4о-место отпадения почки). Во время дождя они вымываются из корзиночки водой и, попав на землю, нарастают при помощи точек роста, формируя новые талломы маршанции.
Вообще, другие виды Hepaticopsida образуют разнообразные органы вегетативного размножения: выводковые тельца, выводковые почки и выводковые ветви, клубеньки и другие.
Половое размножение
(Рисунок 2 – Маршанция обыкновенная – цикл развития)
Маршанция – растение двудомное. При половом размножении на мужских талломах(рис.2 -1) развиваются выросты в виде многолопастного диска, расположенного на ножке. Это мужские антеридиальные подставки(рис.2-3). В верхней части диска находятся антеридиальные полости(рис.2-4), на дне которых находится по 1 антеридию. Антеридий(рис.2 -5)– овальное тельце с однослойной стенкой, прикрепляется ко дну полости небольшой ножкой(рис.2 -5н). Его содержимое состоит из многочисленных спермагенных клеток, в каждой из которых образуют 2 двужгутиковых сперматозоида(рис.2 -8). При созревании антеридий вскрывается наверху щелью, сперматозоиды во время дождя через канал антеридиальной полости выходят наружу и подплывают к архегонию.
На женских талломах(рис.2 -2) образуются выросты, имеющие ножку, увенчанную многолучевой звездой. Это женские, архегониальные подставки(рис.2 -6). На них находятся архегонии, сидящие группами на лучах и обращенные шейками вниз.
Каждая группа архегониев покрыта общим покровом – перихецием(рис.2 -7б), а каждый архегоний одет частным покрывалом – перианцием(рис.2 -7а), в виде кольцевидного валика у его основания. Перианций затем сильно разрастается. Перихеций и перианций защищают архегонии, а затем и спорогоны от высыхания.
Архегонии(рис.2 -7арх)– небольшие колбообразные тельца, состоят из расширенной части –брюшка и суженной –шейки. Снаружи архегоний(рис.2 -9) окружен бесплодными клетками =стенкой архегония,защищающими от высыхания содержимое. В брюшке находится неподвижная женская гамета – яйцеклетка, над которой расположена брюшная канальевая клетка. Внутри шейки расположены шейковые канальцевые клетки. К моменту созревания яйцеклетки канальцевые клетки и стенки архегония, а также верхние клетки стенки ослизняются, и архегоний на верхушке вскрывается. От его вершины к яйцеклетке ведет канал, заполненный слизью, образовавшейся из расплывшихся клеток. По этой слизи, сперматозоид проникает к яйцеклетке, производя оплодотворение. По мере созревания лучи женской подставки поднимаются, принимая сначала горизонтальное положение, а ко времени созревания спорогонов загибаются кверху. Ткань подставки имеет строение таллома, а ножка состоит из паренхимных клеток. Вдоль ножки (внутри её) протягиваются два тяжа язычковых ризоидов – от нижней стороны таллома до диска или до лучей подставки. По мере развития подставки ножка усиленно растет и выносит антеридии и архегонии высоко над талломом.
Из оплодотворенной яйцеклетки развивается бесполое поколение маршанции –спорофит или спорогон(рис.2 -10). Оплодотворенная яйцеклетка тотчас же делится поперечной перегородкой на две клетки. Из нижней клетки благодаря последующим делениям формируется ножка спорогона(рис.2-10н). Из верхней клетки развивается коробочка(рис.2 -10к). Верхняя клетка делится двумя взаимно перпендикулярными перегородками на 4 клетки, а последние, благодаря делению в тангентальном направлении, отчленяют наружные клетки, создающие стенки коробочки, и внутренние, образующие археспорий.
Археспориальные клетки делятся многократно и образуют материнские клетки спор. Одна часть материнских клеток спор приступает к делению. Первое их деление редукционное. Образовавшиеся две клетки делятся уже митотически ещё раз, и в результате образуется тетрада (четверка) спор. Таким образом, споры гаплоидны, так как образовались в результате редукционного деления, которое у всех высших растений всегда происходит при образовании спор. Споры одеты двумя оболочками – наружной–экзоспорием и внутренней, тонкой –эндоспорием. Эндоспорий облегает внутреннее содержимое споры, состоящее из плазмы и ядра.
Другая часть материнских клеток спор вытягивается и благодаря неравномерному утолщению оболочки спирально закручиваются. Каждая такая клетка превращается в пружинку – элатеру.
Итак, спорогон состоит из шаровидной коробочки(рис.2 -10к,11кор), короткой ножки, расширенная нижняя часть которой называется гаусторией(рис.2 -10г). При помощи гаустории спорогон внедряется в ткань подставки (гаметофита) и получает из нее питательные вещества. Коробочка содержит споры (рис.2 -12сп) и пружинки – элатеры(рис.2 -12 эл). Пружинки представляют собой удлиненные мертвые клетки, тонкие стенки которых имеют спиральные утолщения в виде одной или двух спиралей, идущих навстречу друг другу. Функция зрелых элатер, подвижных вследствие гигроскопичности спиральных утолщений, сводится к выбрасыванию спор из коробочки или разрыхлению их в ней.
Первоначально спорогон находится в брюшке архегония(рис.2 -10бр), стенки которого разрастаются и образуют колпачок(рис.2 -11к). Снаружи спорогон окружен также сильно разросшимся перианцием(рис.2 -10п,11п). При созревании коробочки ножка спорогона удлиняется, в результате коробочка разрывается на верхушке и выносится ножкой за пределы лучей подставки. Коробочка вскрывается створками, после чего споры высыпаются наружу. Пружинки – элатеры скручиваются в сухую погоду и раскручиваются в сырую, благодаря этому споровая масса разрыхляется и равномерно рассеивается.
При прорастании споры наружная оболочка – экзоспорий лопается, а содержимое, окруженное эндоспорием, вытягивается в короткую нить. На конце нити отчленяется клетка, которая делится в разных направлениях. В результате формируется небольшая пластинчатая протонема(рис.2 -13,14), далее развивающаяся в таллом маршанции.
В народной медицине спиртовую настойку маршанции обыкновенной применяют при болезнях печени. Лекарственные свойства печёночныхмхов практически мало изучены, они ждут своего исследователя.
ПЕЛЛИЯ ЭПИФИЛЛА .
(Рисунок 3 – Пеллия эпифилла)
Отдел – МОХООБРАЗНЫЕ (Bryophyta)
Класс – ПЕЧЁНОЧНИКИ или ПЕЧЁНОЧНЫЕ МХИ (Marchantiopsida или Hepaticopsida)
Подкласс – ЮНГЕРМАННИЕВЫЕ(Jungermanniidae)
Порядок – МЕТЦГЕРИЕВЫЕ (Metzgeriales)
Семейство –Пеллиевые (Pelliaceae)
Род – Пеллия (Pellia)
Вид – Пеллия эпифилла (Pellia epiphylla)
Пеллия эпифилла широко распространена у нас по сырым лесным оврагам, около ручьёв и канав. Тонкие лентовидные узкие талломы пеллии(рис.3 -1) дихотомически ветвятся. Они светло–зелёные, в центре многослойные, по краям однослойные. Таллом имеет очень простое анатомическое строение. Он одет верхним и нижним эпидермисами и состоит из немногих слоев паренхимных клеток. От клеток нижнего эпидермиса, нерезко отграниченного от основной ткани таллома, отходят простые ризоиды(рис.3 -1а). Воздушных камер с ассимиляторами нет, а хлорофилловые зерна имеются во всех клетках таллома.
Вегетативное размножение
Вегетативное размножениепеллии осуществляется за счет выводковых почек.
Половое размножение
При половом размножении подставок не образуется, антеридии(рис.3 -1б) и архегонии(рис.3 -1в) расположены прямо на талломе.
Антеридии(рис.3 -2) расположены вдоль средней жилки таллома. Они шаровидной формы(рис.3 -2а), сидят на короткой ножке и погружены в ткань таллома. Над антеридием эпидермис прерывается и несколько возвышается, образуя отверстие, идущее в антеридиальную полость. В антеридии развиваются двужгутиковые сперматозоиды(рис.3 -4).
Архегонии(рис.3 -3) возникают группами в небольшом углублении, вблизи точки роста. Они имеют обычное строение (смотри описание Маршанции обыкновенной).
Сперматозоид проникает к яйцеклетке через канал шейки архегония(рис.3 -5). После оплодотворения образуется спорогон(рис.3 -6). На разрезе спорогона(рис.3 -7) видно, что он состоит из коробочки со спорами(рис.3 -7к), ножки(рис.3 -7н), гаустории(рис.3 -7г), которая внедряется в ткань таллома. Первоначально спорогон заключён в брюшке архегония(рис.3 -7бр). Затем, ко времени созревания спор, ножка спорогона сильно удлиняется (обычно это бывает весной следующего года) и выносит коробочку высоко над талломом. У основания ножки сохраняется колпачок – остаток брюшка архегония.
Коробочка вскрывается на верхушке четырьмя створками(рис.3 -8), и из неё высыпаются споры. Пружинки у пеллии, собраны в пучок, прикрепляющийся в центре коробочки – так называемый элатофор(рис.3 -7э). Пружинки элатофора при вскрытии коробочки раскручиваются, распрямляются (элатофор становится похожим на малярную кисточку (рис.3 -8) и мощно разбрасывают споры(рис.3 -9) из коробочки.
Спора при прорастании образует короткую протонему(рис.3 -10), вскоре развивающуюся в новый таллом пеллии.
СФАГНУМ или БЕЛЫЙ МОХ.
(Рисунок 4 - Сфагнум болотный)
Отдел – МОХООБРАЗНЫЕ (Bryophyta)
Класс – МХИ(MUSCI или BRYOPSIDA)
Подкласс – СФАГНОВЫЕ или БЕЛЫЕ или ТОРФЯНЫЕ МХИ (SPHAGNIDAE)
Порядок – СФАГНОВЫЕ (SPHAGNALES)
Семейство – Сфагновые (Sphagnaceae)
Род – Сфагнум (Sphagnum)
Вид – Сфагнум болотный (Sphagnum polustre)
Наиболее широко распространен на равнинах в умеренных и холодных областях северного полушария, образует сплошные покровы на болотах, в заболоченных лесах, за-нимает огромные пространства в Евразии и Северной Америке. В горных областях растет на влажных скалах. У нас наибольшего развития сфагны получают на севере лесной зоны. В тундре их роль резко снижается.
Сфагнум произрастает густыми, плотными дернинами. На внешний вид(рис.4 -1) имеет листостебельное строение. На стебле находятся стеблевые листья, маленькие, язычковид-ные, незаметные, в пазухах некоторых из них расположены ветви, собранные пучками (по 2–7). На ветвях расположены веточные листья, на разных участках стебля ветви имеют различный вид. На верхушке стебля – короткие веточки, плотно скрученные в головку. Листья на них сидят тесно – это верхушечные ветви (рис.4 -2ан.п.). Ниже по стеблю пучки расположены на заметном расстоянии друг от друга, с более редким расположением листьев – торчащие ветви (рис.4 -2г.п.). Остальные ветви пучка – очень длинные, с редкими листьями, свешиваются вниз вдоль стебля – свисающие ветви(рис.4 -2св.п.). В конце лета одна из верхушечных ветвей начинает расти вверх, продолжая рост сфагнума, нижний конец стебля постепенно отмирает.
Анатомическое строение
Центральная часть стебля занята сердцевиной (рис.4 -строение стебля – сц), образованной широкими паренхимными клетками. Далее идет кора. Внутрення кора представлена несколькими рядами клеток с утолщенными стенками, окрашенными в бурый или красный цвет. Это механическая ткань (рис.4 -строение стебля - м.т.) или древесный цилиндр. Он придает прочность стеблю. Внешняя кора состоит из нескольких слоев широких мертвых прозрачных клеток, стенки которых имеют спиральные гиалиновые утолщения(рис.4 -строение стебля - г) и поры (рис.4 -строение стебля -п). Снаружи стебель одет эпидермисом, клетки которого имеют такое же строение. Внешнюю кору и эпидермис часто называют многослойным эпидермисом – гиалодермисом(рис.4 -строение стебля - гд). Через поры соседние клетки гиалодермиса сообщаются между собой. Они всасывают воду, которая легко и быстро передвигается по стеблю.
Веточные листья сфагнума ланцетные или яйцевидно–ланцетные. Состоят из одного слоя клеток, не имеют средней жилки. Содержат клетки двух родов: хлорофиллоносные (рис.4 -строение листа - х.к.) и водоносные = гиалиновые (рис.4 -строение листа - г.к.). Хлорофилло-носные клетки узкие, червеобразные, содержат хлорофилловые зерна. Они зажаты между широкими бесцветными гиалиновыми клетками, которые заполнены водой. Стенки водо-носных клеток имеют спиральные утолщения(рис.4 -строение листа – г) и продырявлены отверстиями, порами (рис.4 -строение листа - п).
Поры могут быть простыми и окаймленными, их число и расположение в клетке различны. Эти признаки строго постоянны для определенных видов сфагнума. На долю водоносных клеток сфагнума приходится до 1/3 поверхности листа. Такое анатомическое строение обеспечивает сфагнуму быстрое всасывание и проведение воды. Описанным выше строением объясняется и белесоватый цвет сфагнума. Листья разных ветвей отличаются формой, величиной, количеством и расположением пор в гиалиновых клетках.
Ассимилирующими являются верхушечные и торчащие ветви, свисающие образуют вокруг стебля как бы чехол, способствуя быстрому продвижению воды вдоль стебля. На стебле сфагнума расположены стеблевые листья. Они также однослойные, без средней жилки, состоят из хлорофиллоносных и гиалиновых клеток.
Взрослые растения сфагнума лишены ризоидов, вода всасывается непосредственно его стеблем и листьями. Дернины сфагнума ежегодно отмирают в нижней части, и одна из верхушечных ветвей продолжает рост стебля вверх. Отмершие части сфагнума вследствие обводнения и нехватки кислорода не разлагаются до конца, а образуют слои полуразложившихся остатков в виде торфа. При этом образуются органические кислоты, которые ещё больше угнетают бактериальную флору и задерживают разложение торфа.
Читайте также: