
Гипотезы происхождения эукариотической клетки кратко
Обновлено: 02.07.2024
Именно появление эукариотической клетки является одним из самых значимых событий биологической эволюции. Отличие эукариотических организмов от прокариотических состоит в более совершенной системе регуляции генома. А благодаря этому возросла приспособляемость одноклеточных организмов, их способность адаптироваться к меняющимся условиям среды без внесения наследственных изменений в геном. Благодаря возможности адаптироваться эукариоты смогли стать многоклеточными – в многоклеточном организме клетки с одним и тем же геномом, в зависимости от условий, образуют совершенно разные по морфологии и по функции ткани.
Этот ароморфоз произошел на рубеже Архея и Протерозоя (2,6 – 2,7 млрд. лет назад), что определили по биомаркерам – остаткам стероидных соединений, свойственных только эукариотическим клеткам. Появление эукариот совпадает по времени с кислородной революцией.
Считается общепризнанным, что эукариоты появились в результате симбиоза нескольких разновидностей прокариот. Видимо, митохондрии произошли от альфа-протеобактерий (аэробных эубактерий), пластиды – от цианобактерий, а цитоплазма – от неизвестной архебактерии. Пока еще нет общепринятой теории возникновения ядра, цитоскелета, а также жгутиков. Гипотезы возникновения жизни на Земле не внесли ясности в вопрос о возникновении клетки. Если о происхождении прокариотов практически нет никаких гипотез, правдоподобно описывающих их возникновение, то что касается происхождения эукариотических клеток, есть несколько точек зрения.
Основные гипотезы происхождения эукариот:
1. Симбиотическая гипотеза основана на двух концепциях. Согласно первой из этих концепций, самое фундаментальное разграничение в живой природе – это разграничение между бактериями и организмами, состоящими из клеток с истинными ядрами – протистами, животными, грибами и растениями. Вторая концепция состоит в том, что источником некоторых частей эукариотических клеток была эволюция симбиозов – формирование постоянных ассоциаций между организмами разных видов. Предполагается, что три класса органелл – митохондрии, реснички и фотосинтезирующие пластиды произошли от свободно живущих бактерий, которые в результате симбиоза были включены в состав клеток прокариот-хозяев. Эта теория в большой мере опирается на неодарвинистские представления, развитые генетиками, экологами, цитологами, которые связали Менделевскую генетику с Дарвиновской идеей естественного отбора. Она опирается также на молекулярную биологию, особенно на данные о структуре белков и последовательности аминокислот, на микропалеонтологию, изучающую наиболее ранние следы жизни на Земле, и на физику и химию атмосферы, поскольку эти науки имеют отношение к газам биологического происхождения.
2. Инвагинационная гипотеза говорит, что предковой формой эукариотической клетки был аэробный прокариот. Внутри находилось несколько геномов, прикреплявшихся к клеточной оболочке. Корпускулярные органеллы и ядро, возникли путем впячивания и отшнуровывания участков оболочки с последующей функциональной специализацией в ядро, митохондрий, хлоропласты. Потом в процессе эволюции произошло усложнение ядерного генома и появилась система цитоплазматических мембран. Эта гипотеза объясняет наличие в оболочках ядра, митохондрий, хлоропластов, двух мембран. Но она встречается с трудностями в объяснении различий в деталях процесса биосинтеза белка в корпускулярных органеллах и цитоплазме эукариотической клетки. В митохондриях и хлоропластах этот процесс в точности соответствует таковому в современных прокариотических клетках.
Происхождение эукариотической клетки согласно симбиотической (I) и инвагинационной (II) гипотезам:
1 — анаэробный прокариот (клетка-хозяин), 2 — прокариоты, имеющие митохондрии, 3 — сине-зеленая водоросль (презумптивный хлоропласт), 4 — сиирохетообразная бактерия (презумпгивный жгутик), 5 — примитивный эукариот со жгутиком, 6 — растительная клетка, 7 — животная клетка со жгутиком, 8 — аэробный прокариот (презумптивная митохондрия), 9 — аэробный прокариот (клетка-родоначальница согласно гипотезе II), 10 — инвагинации клеточной оболочки, давшие ядро и митохондрии, 11 — примитивный эукариот 12 — впячивание клеточной оболочки, давшее хлоропласт, 13 — растительная клетка; а —ДНК прокариотической клетки, б — митохондрия, в —ядро эукариотической клетки, г — жгутик, д — хлоропласт.
Имеющихся данных пока недостаточно для того, чтобы отдать предпочтение какой-то одной из гипотез или выработать новую, которая устроила бы большинство ученых, но в последние годы удалось убедительно доказать симбиогенетическую теорию происхождения эукариотической клетки.
Эволюционные возможности клеток эукариотического типа выше, чем прокариотического. Ведущая роль здесь принадлежит ядерному геному эукариот, который превосходит по размерам геном прокариот. Важные отличия заключаются в диплоидности эукариотических клеток благодаря наличию в ядрах двух комплектов генов, а также в многократном повторении некоторых генов.
Аэробное дыхание также послужило предпосылкой для развития многоклеточных форм. Сами эукариотические клетки появились на Земле после того, как концентрация O2 в атмосфере достигла 1% (точка Пастера). А эта концентрация –необходимое условием аэробного дыхания.
Известно, что каждая эукариотическая клетка содержит геномы различного происхождения: в клетках животных и грибов это геномы ядра и митохондрий, а в клетках растений – также и пластид. Небольшая кольцевая ДНК содержится и в базальном тельце жгутиков эукариотических клеток.
Согласно методу молекулярных часов эукариоты возникли тогда же, когда и прокариоты. Но очевидно же, что в течение значительной части истории Земли на ней доминировали прокариоты. Первые клетки, которые соответствуют эукариотическим размерам (акритархи), имеют возраст 3 млрд. лет, но их природа все еще остается неясной. Почти несомненные остатки эукариот имеют возраст около 2 млрд. лет. И лишь после кислородной революции на поверхности планеты сложились благоприятные условия для эукариот (около 1 млрд. лет назад).
Скорее всего, основным предком эукариотических клеток стали архебактерии, которые перешли к питанию путем заглатывания пищевых частиц. Изменение формы клетки, необходимое для такого заглатывания, обеспечивал состоящий из актина и миозина цитоскелет. Наследственный аппарат такой клетки переместился вглубь от ее изменчивой поверхности, сохранив при этом свою связь с мембраной. А уже это послужило причиной возникновения ядерной оболочки с ядерными порами.
Поглощенные клеткой-хозяином бактерии могли продолжить свое существование внутри нее. Так, предками митохондрий стала группа фотосинтезирующих бактерий – пурпурные альфапротеобактерии. Внутри клетки-хозяина они утратили способность к фотосинтезу и приняли на себя окисление органических веществ. Благодаря им эукариотические клетки стали аэробными. Симбиозы с другими фотосинтезирующими клетками стали причиной приобретения растительными клетками пластид. Возможно, жгутики эукариотических клеток произошли в результате симбиоза клеток-хозяев с бактериями, которые были способны к извивающимся движениям.
Наследственный аппарат эукариотических клеток был устроен примерно так же, как у прокариот. Но в связи с необходимостью управления более крупной и сложной клеткой, позже изменилась организация хромосом, а ДНК оказалась связана с белками-гистонами. Прокариотическая организация сохранилась у геномов внутриклеточных симбионтов.
В результате различных актов симбиогенеза возникли различные группы эукариотических организмов: эукариотической клетка + цианобактерия = красные водоросли; эукариотическая клетка + бактерия-прохлорофита = зеленые водоросли. Даже хлоропласты золотистых, диатомовых, бурых и криптомонадовых водорослей возникли в результате двух последовательных симбиозов, о чем говорит наличие у них 4 мембран.
Появление эукариот было приурочено к такому периоду истории биосферы, когда условия были особенно нестабильны и непредсказуемы, когда приспособительная стратегия прокариот (быстрое мутирование, горизонтальный обмен генами и отбор устойчивых клонов) оказалась слишком расточительной и недостаточно эффективной. В такой ситуации большое преимущество могла получить принципиально более универсальная и экономичная адаптивная стратегия, основанная на развитии целесообразной модификационной изменчивости.
На примере эукариот отчетливо показано, что появление новых форм жизни следует рассматривать не как результат эволюции каких-то отдельных филетических линий или клад, а как закономерный и неизбежный эффект развития систем высшего порядка – сообществ, биосферы и, возможно, всей планеты как единого целого.
Используемые источники:
А. В. Марков, А. М. Куликов. Происхождение эукариот как результат интеграционных процессов в микробном сообществе
А. В. Марков. Проблема происхождения эукариот
М. В. Ларина. Гипотезы происхождения эукариотических клеток. Возникновения многоклеточности
2.Инвагинационная. Инвагинационная гипотеза происхождения эукариотической клетки исходит из того, что предковый формой был аэробный прокариот. Он содержал несколько геномов, каждый из которых прикреплялся к клеточной оболочке. Корпускулярные органеллы, имеющие ДНК, а также ядро, возникли в результате впячивания и отшнуровки фрагментов оболочки вместе с геномом с последующей функциональной специализацией в ядро, митохондрии, хлоропласты, усложнением ядерного генома, развитием цитоплазматических мембран. Эта гипотеза удовлетворительно объясняет наличие 2 мембран в оболочке ядра, митохондрий и хпоропластов. Она встречается с трудностями в объяснении различий в деталях процесса биосинтеза белка в корпускулярных органеллах и цитоплазме эукариотической клетки. В митохондриях и хлоропластах этот процесс в точности соответствует таковому в современных прокариотических клетках. Симбиотическая и инвагинационная гипотезы не исчерпывают все точки зрения на происхождение эукариотического типа клеточной организации.
Клетка – элементарная биологическая система. Клеточная теория Т. Шванна и М. Шлейдена, история, ее основные положения. Современное состояние клеточной теории. Значение клеточной теории.
Клеточная теория — одно из общепризнанных биологических обобщений, утверждающих единство принципа строения и развития мира растений, животных и остальных живых организмов с клеточным строением, в котором клетка рассматривается в качестве общего структурного элемента живых организмов.
Клеточная теория
1.Все организмы состоят из клеток.
2.Все клетки развиваются по единому плану.
3.Свойства многоклеточного организма сводятся к арифметической сумме свойств тех клеток, которые его слагают.
Шлейден предложил считать ядро наиболее постоянной структурой клетки. Многие положения оказались неверными (положение 3). Клетка стала изучаться. Клеточная теория оказала большое влияние на биологию и медицину.

Современная клеточная теория
1.Жизнь существует только в форме клеток.
2.В основе непрерывности жизни лежит клетка.
3.Принцип комплиментарности (связь между структурой и функцией).
Активность организма зависит от клетки, в клетке хранится, перерабатывается наследственная информация. Клетка – основная единица, через которую проходит, запасается и перерабатывается энергия, вещество. Клетка простейшего практически бессмертна. При половом размножении вечны гаметы. В клетке клеточные структуры связаны между собой, все биохимические процессы происходят в соответствующей структуре.
Современная клеточная теория исходит из того, что клеточная структура является главнейшей формой существования жизни, присущей всем живым организмам, кроме вирусов. Совершенствование клеточной структуры явилось главным направлением эволюционного развития как у растений, так и у животных, и клеточное строение прочно удержалось у большинства современных организмов.
Вместе с тем должны быть подвергнуты переоценке догматические и методологически неправильные положения клеточной теории:
Клеточная структура является главной, но не единственной формой существования жизни. Неклеточными формами жизни можно считать вирусы. Правда, признаки живого (обмен веществ, способность к размножению и т.п.) они проявляют только внутри клеток, вне клеток вирус является сложным химическим веществом. По мнению большинства учёных, в своём происхождении вирусы связаны с клеткой, являются частью её генетического материала, "одичавшими" генами.
Выяснилось, что существует два типа клеток - прокариотические (клетки бактерий и архебактерий), не имеющие отграниченного мембранами ядра, и эукариотические (клетки растений, животных, грибов и протистов), имеющие ядро, окружённое двойной мембраной с ядерными порами. Между клетками прокариот и эукариот существует и множество иных различий. У большинства прокариот нет внутренних мембранных органоидов, а у большинства эукариот есть митохондрии и хлоропласты. В соответствии с теорией симбиогенеза, эти полуавтономные органоиды - потомки бактериальных клеток. Таким образом, эукариотическая клетка - система более высокого уровня организации, она не может считаться целиком гомологичной клетке бактерии (клетка бактерии гомологична одной митохондрии клетки человека). Гомология всех клеток, таким образом, свелась к наличию у них замкнутой наружной мембраны из двойного слоя фосфолипидов (у архебактерий она имеет иной химический состав, чем у остальных групп организмов), рибосом и хромосом - наследственного материала в виде молекул ДНК, образующих комплекс с белками. Это, конечно, не отменяет общего происхождения всех клеток, которое подтверждается общностью их химического состава.
Клеточная теория рассматривала организм как сумму клеток, а жизнепроявления организма растворяла в сумме жизнепроявлений составляющих его клеток. Этим игнорировалась целостность организма, закономерности целого подменялись суммой частей.
Догматическая клеточная теория игнорировала специфичность неклеточных структур в организме или даже признавала их, как это делал Вирхов, неживыми. В действительности, в организме кроме клеток есть многоядерные надклеточные структуры (синцитии, симпласты) и безъядерное межклеточное вещество, обладающее способностью к метаболизму и потому живое. Установить специфичность их жизнепроявлений и значение для организма является задачей современной цитологии. В то же время и многоядерные структуры, и внеклеточное вещество появляются только из клеток. Синцитии и симпласты многоклеточных - продукт слияния исходных клеток, а внеклеточное вещество - продукт их секреции, т.е. образуется оно в результате метаболизма клеток.
Целостность организма есть результат естественных, материальных взаимосвязей, вполне доступных исследованию и раскрытию. Клетки многоклеточного организма не являются индивидуумами, способными существовать самостоятельно (так называемые культуры клеток вне организма представляют собой искусственно создаваемые биологические системы). К самостоятельному существованию способны, как правило, лишь те клетки многоклеточных, которые дают начало новым особям (гаметы, зиготы или споры) и могут рассматриваться как отдельные организмы. Клетка не может быть оторвана от окружающей среды (как, впрочем, и любые живые системы). Сосредоточение всего внимания на отдельных клетках неизбежно приводит к унификации и механистическому пониманию организма как суммы частей.
Очищенная от механицизма и дополненная новыми данными клеточная теория остается одним из важнейших биологических обобщений.
2.Инвагинационная. Инвагинационная гипотеза происхождения эукариотической клетки исходит из того, что предковый формой был аэробный прокариот. Он содержал несколько геномов, каждый из которых прикреплялся к клеточной оболочке. Корпускулярные органеллы, имеющие ДНК, а также ядро, возникли в результате впячивания и отшнуровки фрагментов оболочки вместе с геномом с последующей функциональной специализацией в ядро, митохондрии, хлоропласты, усложнением ядерного генома, развитием цитоплазматических мембран. Эта гипотеза удовлетворительно объясняет наличие 2 мембран в оболочке ядра, митохондрий и хпоропластов. Она встречается с трудностями в объяснении различий в деталях процесса биосинтеза белка в корпускулярных органеллах и цитоплазме эукариотической клетки. В митохондриях и хлоропластах этот процесс в точности соответствует таковому в современных прокариотических клетках. Симбиотическая и инвагинационная гипотезы не исчерпывают все точки зрения на происхождение эукариотического типа клеточной организации.
Клетка – элементарная биологическая система. Клеточная теория Т. Шванна и М. Шлейдена, история, ее основные положения. Современное состояние клеточной теории. Значение клеточной теории.
Клеточная теория — одно из общепризнанных биологических обобщений, утверждающих единство принципа строения и развития мира растений, животных и остальных живых организмов с клеточным строением, в котором клетка рассматривается в качестве общего структурного элемента живых организмов.
Клеточная теория
1.Все организмы состоят из клеток.
2.Все клетки развиваются по единому плану.
3.Свойства многоклеточного организма сводятся к арифметической сумме свойств тех клеток, которые его слагают.
Шлейден предложил считать ядро наиболее постоянной структурой клетки. Многие положения оказались неверными (положение 3). Клетка стала изучаться. Клеточная теория оказала большое влияние на биологию и медицину.
Современная клеточная теория
1.Жизнь существует только в форме клеток.
2.В основе непрерывности жизни лежит клетка.
3.Принцип комплиментарности (связь между структурой и функцией).
Активность организма зависит от клетки, в клетке хранится, перерабатывается наследственная информация. Клетка – основная единица, через которую проходит, запасается и перерабатывается энергия, вещество. Клетка простейшего практически бессмертна. При половом размножении вечны гаметы. В клетке клеточные структуры связаны между собой, все биохимические процессы происходят в соответствующей структуре.
Современная клеточная теория исходит из того, что клеточная структура является главнейшей формой существования жизни, присущей всем живым организмам, кроме вирусов. Совершенствование клеточной структуры явилось главным направлением эволюционного развития как у растений, так и у животных, и клеточное строение прочно удержалось у большинства современных организмов.
Вместе с тем должны быть подвергнуты переоценке догматические и методологически неправильные положения клеточной теории:
Клеточная структура является главной, но не единственной формой существования жизни. Неклеточными формами жизни можно считать вирусы. Правда, признаки живого (обмен веществ, способность к размножению и т.п.) они проявляют только внутри клеток, вне клеток вирус является сложным химическим веществом. По мнению большинства учёных, в своём происхождении вирусы связаны с клеткой, являются частью её генетического материала, "одичавшими" генами.
Выяснилось, что существует два типа клеток - прокариотические (клетки бактерий и архебактерий), не имеющие отграниченного мембранами ядра, и эукариотические (клетки растений, животных, грибов и протистов), имеющие ядро, окружённое двойной мембраной с ядерными порами. Между клетками прокариот и эукариот существует и множество иных различий. У большинства прокариот нет внутренних мембранных органоидов, а у большинства эукариот есть митохондрии и хлоропласты. В соответствии с теорией симбиогенеза, эти полуавтономные органоиды - потомки бактериальных клеток. Таким образом, эукариотическая клетка - система более высокого уровня организации, она не может считаться целиком гомологичной клетке бактерии (клетка бактерии гомологична одной митохондрии клетки человека). Гомология всех клеток, таким образом, свелась к наличию у них замкнутой наружной мембраны из двойного слоя фосфолипидов (у архебактерий она имеет иной химический состав, чем у остальных групп организмов), рибосом и хромосом - наследственного материала в виде молекул ДНК, образующих комплекс с белками. Это, конечно, не отменяет общего происхождения всех клеток, которое подтверждается общностью их химического состава.
Клеточная теория рассматривала организм как сумму клеток, а жизнепроявления организма растворяла в сумме жизнепроявлений составляющих его клеток. Этим игнорировалась целостность организма, закономерности целого подменялись суммой частей.
Догматическая клеточная теория игнорировала специфичность неклеточных структур в организме или даже признавала их, как это делал Вирхов, неживыми. В действительности, в организме кроме клеток есть многоядерные надклеточные структуры (синцитии, симпласты) и безъядерное межклеточное вещество, обладающее способностью к метаболизму и потому живое. Установить специфичность их жизнепроявлений и значение для организма является задачей современной цитологии. В то же время и многоядерные структуры, и внеклеточное вещество появляются только из клеток. Синцитии и симпласты многоклеточных - продукт слияния исходных клеток, а внеклеточное вещество - продукт их секреции, т.е. образуется оно в результате метаболизма клеток.
Целостность организма есть результат естественных, материальных взаимосвязей, вполне доступных исследованию и раскрытию. Клетки многоклеточного организма не являются индивидуумами, способными существовать самостоятельно (так называемые культуры клеток вне организма представляют собой искусственно создаваемые биологические системы). К самостоятельному существованию способны, как правило, лишь те клетки многоклеточных, которые дают начало новым особям (гаметы, зиготы или споры) и могут рассматриваться как отдельные организмы. Клетка не может быть оторвана от окружающей среды (как, впрочем, и любые живые системы). Сосредоточение всего внимания на отдельных клетках неизбежно приводит к унификации и механистическому пониманию организма как суммы частей.
Очищенная от механицизма и дополненная новыми данными клеточная теория остается одним из важнейших биологических обобщений.
Спонтанное образование органических молекул и продолжительная эволюция дали начало прокариотическим клеткам, которые появились на планете примерно 3,5 млрд. лет назад. Такую гипотезу выдвинул А. И. Опарин.
Решающим этапом в этом процессе стало появление ферментативных (каталитических) молекулярных механизмов.
Для первых клеток было характерным использование каталитических свойств белков и РНК. При этом, РНК у них выступала в качестве вещества наследственности. Постепенно структура и функции клеток становились сложнее, накапливались дополнительные каталитические белки. В итоге место молекулы РНК заняла двухцепочная ДНК, взявшая на себя функцию сохранения генетической информации.
Симбиотическая гипотеза объясняет, как появились эукариотические клетки. Согласно этой гипотезы, клеткой-хозяином выступал анаэроб. Аэробное дыхание возникло в связи с тем, что аэробные бактерии проникли в клетку-хозяина и стали сосуществовать с ней в виде митохондрий.
Благодаря наличию в зеленых растениях хлоропластов, они способны к фотосинтезу. Хлоропласты, по мнению многих, возникли из прокариотических сине-зеленых водорослей: они были симбионтами клетки-хозяина.
Состоятельность симбиотической гипотезы подтверждается следующим аргументом: в митохондриях и хлоропластах есть собственная ДНК.
Система внутриклеточных мембран, образующая гладкую и зернистую эндоплазматические сети (ЭПС), ядерную оболочку и комплекс Гольджи, является производной внешней мембраны клетки, которой характерно ветвление.
Происхождение генетического материала ядра является довольно сложным вопросом. Есть предположения, что он тоже образовался из симбиотических прокариот. Скорее всего, что увеличение числа ядерной ДНК происходило постепенно — по мере перемещения генетического материала из геномов симбионтов в участок клетки, ограниченный мембраной.
Появление митоза, как механизма, равномерно распределяющего генетический материал и воспроизводящего клетки, также было очень важным. В процессе эволюции возник еще одни механизм деления клеток — мейоз. Это решило проблему, связанную с размножением многоклеточных организмов.
В связи с переходом к половому размножению возникла комбинаторная изменчивость, а также заметно увеличилась скорость эволюции.
Все эти процессы на протяжении 1 млрд лет эволюции позволили эукариотическому типу клеточной эволюции обусловить многообразие живых организмов и пройти путь от простейших до человека.
Эволюция клетки прокариот
Ископаемые останки бактерий и цианобактерий, изученные учеными, дают основания предполагать, что начальная клеточная форма — это примитивная прокариотическая клетка, возникшая 3,5 млрд. лет назад.
Чтобы обеспечить собственное существования, клетки этого типа использовали органические молекулы небиологического происхождения. Образование мембраны, окружающей клеточное вещество, стало первым шагом на пути формирования примитивной клетки.
Позже в клетках примитивных прокариот возникли механизмы синтеза и энергетического обеспечения. У первых прокариотических клеток были простейшие каталитические системы. Поэтому основой получения ими энергии было брожение.
Затем клетки отдельных видов прокариот сменили процесс брожения на дыхание, в результате чего получение энергии стало более эффективным.
Эволюция клетки эукариот
Клетки эукариот в ходе эволюции менялись за счет увеличения разнообразия форм, структуры и функций, а также компартментализации биохимических систем и сохранения общего для всех аэробного метаболизма, происходящих одновременно.
Возникновение эукариотической клетки из прокариотической произошло примерно менее 1 млрд лет назад.
Есть 4 гипотезы, объясняющие происхождение эукариотической клетки.
Первая гипотеза происхождения эукариот
Одна из гипотез возникновения эукариот — гипотеза клеточного симбиоза. Эта гипотеза имеет наибольшее число сторонников. Согласно ей, эукариотическая клетка — это симбиотическая структура, состоящая из нескольких клеток различного типа, которые окружены общей для всех мембраной.
Многие считают, что пластиды клеток современных зеленых растений возникли из бактерий, предшественников современных цианобактерий, которые способны к фотосинтезу. Митохондрии эукариотических клеток появились из аэробных бактерий — последние вступали в симбиоз с примитивными анаэробными клетками, у которых была способность к фотосинтезу. В результате образовались клетки, которые могли существовать в кислородной атмосфере и использовать кислород в процессе дыхания.
Считается, что ядро — это рудимент какого-либо внутриклеточного симбионта, потерявшего свою цитоплазму после того, как был включен в исходную клетку.
Подтверждением этого являются данные о временных симбиотических связях отдельных организмов.
К примеру, одноклеточная зеленая водоросль хлорелла проживает в цитоплазме зеленого лишайника парамеции. Поскольку для этой водоросли характерен фотосинтез, то она является для парамеции поставщиком питательных веществ.
Пластиды и митохондрии содержат свою систему генетической информации о синтезе белков. Она представлена в виде ДНК, тРНК, мРНК и соответствующих ферментов.
Прокариоты митохондрий и хлоропластов характеризуются похожими репродуктивными способами — они размножаются с помощью простого деления пополам.
Вторая гипотеза происхождения эукариот
Последователи второй гипотезы утверждают, что эукариотическая клетка образовалась от клетки прокариот и содержала несколько геномов, которые были прикреплены к клеточной мембране.
Впячивание клеточной мембраны внутрь цитоплазмы привело к образованию мезосом, у которых в первое время была способность к фотосинтезу. Потом произошла специализация этих органелл: одна из них потеряла способность к дыханию и фотосинтезу и преобразовалась в ядро, а другие — развили эти опции, дав начало митохондриям и пластидам.
Подтверждение этой гипотезы — наличие двойного строения мембран ядра, пластид и митохондрий.
Третья гипотеза происхождения эукариот
В ее основе — мысль о том, что все живые формы произошли от предковых анаэробных гетеротрофов. Эукариоты — это сублиния бесстеночных анаэробных прокариот, у которых есть способность к эндоцитозу.
Четвертая гипотеза происхождения эукариот
Возникновение клеток эукариот произошло из прокариот — изначально они содержали множество распадающихся на части геномов, давших начало структурам с различными функциями. Завершающим этапом было клонирование структур с подобными функциями. Далее они покрывались двойными мембранами, происходило образование ядра, митохондрий и мембранной сети.
Также можно утверждать о схожести генетического кода, который находится в ядерной и митохондриальной ДНК, и регуляции дыхательной функции ядра и митохондрий.
Из всех перечисленных первая гипотеза самая популярная. При этом важно обозначить, что, несмотря на похожесть митохондрий и хлоропластов на временные бактерии-аэробы и цианобактерии, между ними существуют заметные различия. Одно из таких различий заключается в том, что митохондрии и хлоропласты содержат меньше ДНК, чем клетки бактерий.
Эволюционируя, митохондрии и хлоропласты постоянно меняли свои размеры.
Геном эукариот развивался при помощи объединения молекул ДНК и белков. Хроматин и хромосомы в этом случае формировались разной формы и в различном количестве. Отмечалась специализация хроматина: формировались эухроматин и гетерохроматин, аутосомы и половые хромосомы.
Пока не до конца понятно, почему в ходе эволюции у конкретных организмов сформировалось именно такое количество хромосом. Зачастую у примитивных организмов их больше, чем в клетках более эволюционно развитых.
Но что точно ясно, так это то, что в образовании новых видов большую роль играли структурные и количественные изменения в кариотипах. Кроме того, в это же время усложнялась структура и функции компонентов клетки, происходило развитие регуляторных механизмов.
Большое значение также имело эволюционное развитие митоза. Важное условие, обеспечивающее многоклеточность — точное распределение хромосом в процессе митоза. Но логичных объяснений происхождения самого митоза нет. Предполагают, что он развивался из примитивного митоза, в ходе которого при расхождении реплицировавшихся хромосом не происходило разрушения ядерной мембраны.
Эукариоты, возможно, существуют на Земле около 2,7 млрд лет. Эта дата определена благодаря обнаружению в древних отложениях остатков стероидных соединений, встречающихся только в мембранах эукариотических клеток. Однако только примерно 1–1,5 млрд лет назад эукариоты стали превалирующим компонентом биосферы, потеснив прокариот. Появление эукариот – событие планетарного масштаба, так как именно они дали начало огромному многообразию организмов, в том числе ландшафтообразующих. Не случайно этот период в истории Земли называют неопротерозойской революцией.
Гипотезы о происхождении эукариот
С момента обнаружения существенных различий в строении клеток бактерий и простейших, в первую очередь отсутствия или присутствия ядра, встал вопрос о выяснении происхождения и степени родства этих групп. Были выдвинуты три основные гипотезы происхождения эукариот и их органоидов – аутогенная, химерная и симбиогенная, которые и в настоящее время все еще остаются гипотезами.
Согласно аутогенной (от греч. autos – сам, собственный и genesis – происхождение, возникновение), или сукцессионной, гипотезе эукариотическая клетка самостоятельно развилась из прокариотической в результате впячивания плазматической мембраны с ее последующей перестройкой. Таким образом, все мембранные органоиды, в том числе митохондрии и пластиды, ведут свое начало от имеющихся у прокариот внутриплазматических мембранных структур, подобных мезосомам. В настоящее время аутогенная гипотеза считается устаревшей.
Химерная гипотеза трактует образование клетки эукариот путем слияния целых клеток различных прокариот. Предложено несколько вариантов химерного происхождения эукариотической клетки от архей: слияние с фототрофной протеобактерией; слияние архебактерии с эубактерей-бродильщиком, обладающей гликолизом; слияние архебактерии со спирохетой, приведшее к образованию жгутика.
Согласно симбиогенной гипотезе, или гипотезе симбиогенеза, первичная клетка крупной прокариотической бактерии вступала в симбиоз с другими прокариотами и в результате постепенно приобретала различные органоиды: из симбионтных цианобактерий образовались пластиды, из гетеротрофных бактерий – митохондрии, из каких-то иных симбионтов – центриоли и жгутики.
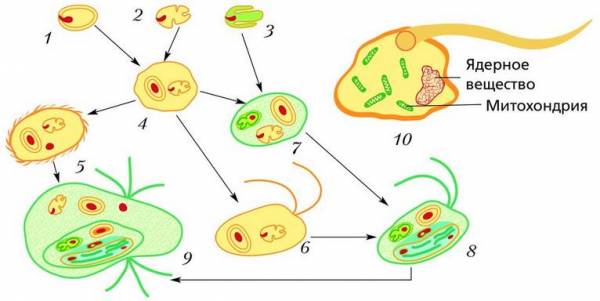
Схема возникновения эукариот согласно симбиогенной гипотезе: 1 – прокариота – будущее ядро; 2 – прокариота – будущая митохондрия; 3 – прокариота – будущий хлоропласт; 4 – первая эукариота с ядром и митохондрией; 5–9 – возможные пути дальнейшего усложнения структуры клетки эукариот; 10 – бактерия-симбионт с митохондриями (первая эукариотическая клетка)
Некоторые сторонники этой гипотезы предполагали даже возможность симбиотического происхождения ядра.
Все эти предположения породили ряд гипотез, объясняющих вероятность подобных явлений. Но в настоящее время ученые еще так и не пришли к единому мнению о происхождении многих внутриклеточных структур эукариот. Однако сама дискуссия вызвала массу интересных работ по изучению митохондрий и пластид, что обеспечило более глубокое и детальное выяснение свойств этих органоидов и других внутриклеточных структур.
Следует отметить, что впервые идею о симбиотическом происхождении сложно устроенной клетки высказал в конце 60-х годов XIX века русский ученый-ботаник Андрей Сергеевич Фаминцын (1835–1918) на основе изучения лишайников. В начале XX века (1905–1909) биолог Константин Сергеевич Мережковский (1855–1921) выдвинул идею о симбиотическом происхождении носителей пигментов в растительных клетках, которая в 1921 – 1924 годах ботаником Борисом Михайловичем Козо-Полянским (1890– 1957) была развита в симбиотическую гипотезу происхождения клеток растений и животных. В 1983 году сотрудницей Массачусетского университета Линн Маргулис (Саган) гипотеза симбиогенеза была изложена в виде целостной концепции. В последующие годы благодаря новым методам исследований и открытиям симбиогенная гипотеза получила многочисленные подтверждения, и иногда ее называют уже не гипотезой, а теорией.
Гипотезы симбиогенеза по-разному отображают и начало самого процесса, и путь возникновения сложных клеток эукариот.
В настоящее время общепризнанной считается гипотеза симбиотического происхождения эукариотической клетки от прокариоты, вероятнее всего – от архебактерии, хотя не исключено, что предком эукариот была вымершая группа каких-то прокариот, одинаково далекая и от эубактерий, и от архебактерий.
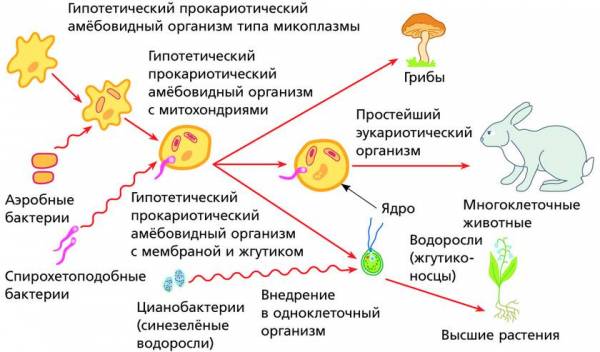
Схема возможного происхождения эукариот от гипотетического прокариотического организма
Получено достаточное количество данных, доказывающих, что митохондрии происходят от симбиотических альфапротеобактерий (близких к современным пурпурным бактериям), а пластиды растений и большинства водорослей – от симбиотических фотосинтезирующих цианобактерий (синезеленых водорослей). Однако до сих пор не прекращаются дискуссии по поводу природы клетки-хозяина, и главное – относительно происхождения ядра.
Например, согласно одной из гипотез образование ядерной мембраны могло стать побочным результатом избыточной мембранообразующей деятельности эубактерий, поселившихся в цитоплазме архебактерий. Согласно другой гипотезе клеточное ядро сформировалось у архей в результате вирусной инфекции. Существует множество других гипотез возникновения ядра, но ни одна пока не доказана.
Один из наиболее вероятных вариантов происхождения ядра – появление его вследствие впячивания участков плазматической мембраны с находящимися на них точками прикрепления хромосом внутрь полиплоидной (имеющей несколько хромосом) прокариотной клетки. Таким путем первичные эукариоты могли получить ядро, ограниченное двойной ядерной оболочкой, но у них еще сохранялось кольцевое строение хромосом и отсутствовали гистоны. В какой-то мере этот эволюционный путь подтверждается тем, что в ядре большинства эукариот хромосомы также прикрепляются к субмембранным структурам внутренней ядерной мембраны и в ядре всегда имеются плазмиды (внехромосомные молекулы ДНК, часто представляющие собой кольцевые двухцепочечные структуры), а у динофлагеллят (панцирных жгутиконосцев) хромосомы содержат кольцевые молекулы ДНК, практически лишенные гистонов.
Читайте также: