
Генетический аппарат бактерий кратко
Обновлено: 03.07.2024
Генетическая система бактерий имеет по крайней мере четыре особенности, присущие только этим организмам:
1. Хромосомы бактерий (и соответственно плазмид) располагаются свободно в цитоплазме, не отграничены от нее никакими мембранами, но связаны с определенными рецепторами на цитоплазматической мембране. Поскольку длина хромосомы (у E. coli около 1,6 мм) во много раз превышает длину бактериальной клетки (1,5 – 3,0 мкм в среднем), хромосома особым компактным образом в ней упакована: молекула хромосомной ДНК находится в суперспирализованной форме и свернута в виде петель, число которых составляет 12 – 80 на хромосому. Петли в центре нуклеоида объединяются за счет связывания ДНК с сердцевинной структурой, представленной молекулами особого класса РНК – 4,5S РНК. Такая упорядоченная упаковка обеспечивает постоянную транскрипцию отдельных оперонов хромосомы и не препятствует ее репликации. Возможно, что петли упакованной хромосомы способствуют компартментализации рибосом.
2. Хотя бактерии являются гаплоидными организмами, т. е. имеют один набор генов, содержание ДНК у них непостоянно, оно может при благоприятных условиях достигать значений, эквивалентных по массе 2, 4, 6 и даже 8 хромосомам. У всех прочих живых существ содержание ДНК постоянное, и оно удваивается (кроме вирусов и плазмид) перед делением.
3. У бактерий в естественных условиях передача генетической информации происходит не только по вертикали, т. е. от родительской клетки дочерним, но и по горизонтали с помощью различных механизмов: конъюгации, сексдукции, трансдукции, трансформации.
4. У бактерий очень часто помимо хромосомного генома имеется дополнительный плазмидный геном, наделяющий их важными биологическими свойствами, нередко – специфическим (приобретенным) иммунитетом к различным антибиотикам и другим химиопрепаратам.
Содержание ДНК у бактерий зависит от условий их роста: при благоприятных условиях оно возрастает до величин, соответствующих массе нескольких хромосом. Это уникальное свойство бактериального генома. Биологическое значение его состоит в том, что, регулируя содержание копий своих генов (а оно будет определяться количеством копий синтезируемых хромосом), бактерии одновременно приспосабливают скорость своего размножения к условиям роста. Наряду с увеличением содержания ДНК у бактерий в этом случае существенно возрастает и количество рибосом. Благодаря этому создаются необходимые условия для транскрипции и трансляции (а у бактерий они происходят одновременно) нескольких копий генов одновременно, возрастает суммарная скорость биосинтеза всех субклеточных и клеточных структур и соответственно скорость размножения бактерий. Время клеточного цикла бактерий сокращается от нескольких часов до 20 – 30 мин. Скорость размножения определяет возможность накопления в окружающей среде большого запаса клеток данного вида. Это и является причиной существования бактерий в природе многие миллионы лет. Возможность регулировать скорость собственного размножения – одно из главных условий, обеспечивающих выживание бактерий в окружающей среде, а следовательно, и сохранение вида в природе.
Генетический аппарат бактерий характеризуется определенными особенностями:
1) в нем отсутствует ядерная оболочка,
2) отсутствует ядрышко,
3) ДНК находится непосредственно в цитоплазме;
4) нет деления на хромосомы,
5) ДНК бактерий аналог хромосомы эукариот;
6) в клетке может находиться несколько копий ДНК,
7) отсутствуют митоз и мейоз.
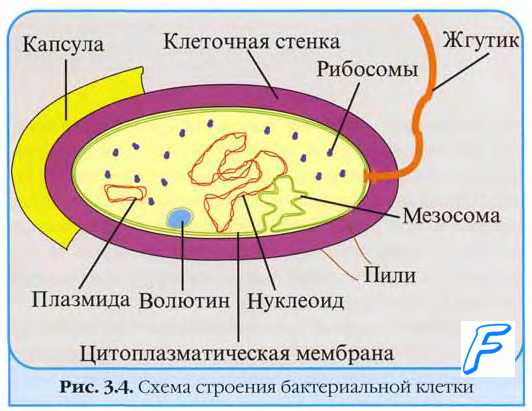
Ядерный аппарат бактерий представлен нуклеоидом - замкнутой в кольцо двуцепочечной молекулой ДНК с молекулярной массой 10 9 -3•10 9 Д, и длиной от 0,25 до 3 мм. Бактериальная хромосома никогда не связана с гистонами. На клетку может приходиться до 40 копий бактериальной хромосомы.
Морфологические формы бактерий
По морфологии бактерии делятся на группы:
1. Шаровидные (кокки). В зависимости от количества плоскостей деления и расположение бактериальных клеток после деления коки подразделяют на следующие виды:
- Микрококки. Разделение происходит в одной или нескольких плоскостях. Клетки располагаются одиночно. Это, как правило, сапрофиты, типичная микрофлора почвы, воды, воздуха. Представители: Micrococcus luteus,
- Диплококки. Клетки делятся в одной плоскости, располагаются попарно. Представители: Azotobacter chroococcus,
- Стрептококки. Клетки делятся в одной плоскости, образуют цепочки различной длины. Представители: Streptococcus lactis;
- Тетракокки. Клетки делятся в двух плоскостях, образуют тетрады. Представитель: Deinococcus proteolytics;
- Сарцины. Деление клеток происходит в трех взаимно перпендикулярных плоскостях с образованием группировок в виде пакетов с 8, 16, 32 клеток. Представитель; Sarcina flava;
- Стафилококки. Клетки делятся в нескольких плоскостях с образованием характерных скоплений, подобных виноградных гроздьев. Представители; Staphylococcus aureus,
2. Палочковидные. Существует несколько классификаций цилиндрических бактерий:
- За спорообразованием: спорообразующие (бациллы) p. Bacillus и p.Clostridium и неспорообразующие (бактерии) - p. Рseudomonas
- Длиной: длинные (большие 3 мкм) Bacillus megaierium; короткие (около 1 мкм) – E. Colli; очень короткие (менее 1 мкм) - Yersinia pestis;
- Поперечным диаметром: тонкие - Mycobacterium tuberculosis; толстые-Bacillus subtilis;
- Формой концов: округлые - Yersinia pestis; со срезанными концами - Salmonella typhi; утолщенные - Corynebacierium diphteriae;
- Расположением: единичные - Serratia marcescens; попарные p. Pseudomonas; в цепочках - p. Bacillus; расположены под углом - Corynehacterium diphteriae
- Вибрионы. Имеют вид комы. Представитель: Vibrio cholerae;
- Спириллы. Спирально изогнутые клетки со значительным поперечным диаметром и малым количеством завитков. Представитель: Spirillum volutans
- Спирохеты. Спирально изогнутые клетки с малым поперечным диаметром и большим количеством завитков. Представитель: Treponema pallidum.
4. Нитчатые - палочковидные одноклеточные и многоклеточные организмы. Нити образуются многими клетками, которые соединяются между собой с помощью чехлов и могут быть простыми или разветвленными. Представители: Beggiatoa,
5. Бактерии необычной формы - в виде колец, звезд, почек, серповидные т.д.
Методы исследования м/о
Основным инструментом для изучения строения бактериальной клетки является световой микроскоп. Общее увеличение микроскопа определяется как произведение увеличения объектива и увеличение окуляра. Световой микроскоп дает максимальное увеличение изображения в 1 000 раз.
Основные элементы его конструкции - механическая часть (штатив, подставка, предметный столик, тубус, тубусодержатель, револьвер), оптическая часть (окуляр, объектив) и система освещения (зеркало, конденсор, диафрагма). Зеркало необходимо для отражения лучей в направлении конденсора. Конденсор - это система короткофокусных линз под предметным столиком. Конденсор собирает световые лучи, отраженные от зеркала, и направляет их через диафрагму, которая служит для регулирования количества лучей, на объект.
Лучи, прошедшие сквозь объект, попадают в объектив, а потом - к окуляру. Объектив представляет собой систему двояковыпуклых линз в металлической оправе Передняя - фронтальная линза - главная, увеличительная. Внутренние-корректирующие - предназначены для устранения аберраций. В микробиологии для исследования микропрепаратов используют иммерсионная систему (Имерсионний объектив и иммерсионная масло). Заменяя воздушную среду между линзой объектива и объектом на иммерсионная вещество (показатели преломления иммерсионной масла и стекла почти одинаковые), можно предотвратить преломления лучей и увеличить разрешение микроскопа. При работе с иммерсионной системой на объект наносят иммерсионное масло и погружают в него фронтальную линзу иммерсионного объектива. Объект фокусируют с помощью микрометрической винта.
В основе метода микроскопии в темном поле лежит явление освещения объекта косыми високоапертурнимы лучами света. Это достигается использованием конденсора с затемненной центральной частью, которая задерживает пучок лучей, и они не попадают в объектив. Поле остается темным. Боковые лучи, проходя сквозь кольцевую щель конденсора, расположенную между его центральной частью и краем, попадают на боковую поверхность линзы, отражаются от нее, направляются под углом к плоскости препарата и фокусируются на нем, если в препарате содержатся какие-либо частицы (объект), то косые лучи, отраженные конденсором, сталкиваются с ними и преломляются на них образуя волны дифрагированного света, которые попадают в объектив. В результате на темном фоне поля зрения видно частицы, которые светятся. Темнопольная микроскопия позволяет наблюдать за движением микробов, изучать микроорганизмы, размеры которых лежат за пределами разрешающей способности светового микроскопа.
Электронная микроскопия основана на том, что электромагнитное поле влияет на пучок электронов, который выходит из электронной пушки и проходит через ряд электромагнитных линз. Конденсорная линза собирает пучок электронов к препарату, а несколько увеличительных линз создают увеличенное изображение, которое проецируется на экран. Электронный микроскоп позволяет получать трехмерное изображение поверхности объектов. Контрастность объекта повышают с помощью напыления солей тяжелых металлов.
Бактериологические методы – это выделение чистых культур микроорганизмов. Чистая культура – это популяция, клеток – потомков одной родительской клетки. Осуществляется путем посева на питательные среды. Для получения чистых культур используют селективные среды, и различные способы посева (истончающийся штрих).
Смотри 3
14. Классификация бактерий.
Класифицируют так же по отношению к О2 (аеробы анаеробы микроаерофилы), к температуре (термофилы нейтрофилы психрофилы), к рН (ацидофилы, алкалофилы), по способам получения энергии и др.
15 Микоплазмы относят к отделу Tenericuies. Микоплазмы – это бактерии, лишенные клеточной стенки. Гр- Для них хар-рен полиморфизм, в культурах одного вида могут встреч. крупные шаровидн.тела, мелкие зерна, диско-, элипсо-, палочко-, нитевидные кл., котор. могут обр. мицелиеподобные структуры. Размеры различны – от 0,1-0,2 мкм (на границе разреш.способн.свет.микроск., это самые мелкие клеточные организмы, способные к самовоспроизвелению) до 10 мкм. Разные способы размножения: фрагментация, бинарное деление, почкование. При делении полученные клетки не равноценны по размеру, часто одна из них даже нежизнеспособна. Кл. микоплазм окружены ЦПМ, снаружи – капсулоподобн.слой. В цитоплазме – нуклеоид, рибосомы, производные ЦПМ. Никогда нет мезосом. ЦПМ стабильна и эластична (холестерин – осн. компонент мембранных липидов микоплазм).Не чувствительны к антибиот. пенициллинового ряда. На пит.средах обр. колонии в виде яичницы-глазуньи. Разнообразны по требованиям к средам: некотор. растут на искусствен. пит. средах, некотор. – только внутри организма хозяина или в культурах кл. Если в среде есть холистерин, глицерин – мицеллярный рост, на плотных пит.средах больше сферич.тел. Получение энергии – за счет окисления или сбражив. органич. соедин. или окислен. неорган.соед. Строгие аэробы и облигатн. анаэр., ацидофилы, термофилы. Паразиты.
Литотрофные микроорганизмы
Используют неорганич. вещества в качестве окисляемых субстратов — доноров электронов. Различают фото- и хемолитотрофные микроорганизмы. У фотолитотрофных микроорганизмов окисляемый субстрат — мол. водород, соединения серы (пурпурные и зелёные бактерии, нек-рые цианобактерии) или вода (цианобактерии, микроформы водорослей) — служит источником восстановителя, а энергию они получают в результате поглощения света. У хемолитотрофньгх микроорганизмов окисляемый неорганич. субстрат — источник и энергии, и восстановителя. Ими могут служить мол. водород (водородные бактерии), окись углерода (карбоксидобактерии), восстановленные соединения серы (тио-новые бактерии), соединения азота (нитрифицирующие бактерии). Окислитель во всех перечисленных случаях — мол. кислород. В анаэробных условиях окислителем могут быть нитрат, нитрит и окислы азота (денитрифицирующие бактерии), сера и (или) сульфат (сульфат-восстанавливающие бактерии), углекислота (метан- и ацетатобразующие бактерии), фумарат и нек-рые др. соединения. Л. м. играют важнейшую роль в природе, замыкая циклы биогенных элементов. Большое значение Л. м. имеют в геологии. Напр., тионовые бактерии обусловливают выщелачивание металлов из горных пород, сульфатвосстанавливающие бактерии участвуют в формировании осадочных отложений серы.
Генетика бактерий. Геном бактерии. Генотип бактериальной клетки. Фенотип бактериальной клетки. Генетический материал бактерий.
В наши дни приоритетным направлением естествознания можно считать молекулярную биологию. Она тесно связана с микробиологией и в известном смысле является её детищем, так как в качестве основных моделей использует бактерии и вирусы, а одно из основных направлений молекулярной биологии — молекулярная генетика — долгое время являлась не чем иным, как генетикой бактерий и бактериофагов.
Изучение генетики бактерий имеет также и несомненный прикладной интерес, например в плане установления механизмов передачи патогенных свойств и устойчивости к лекарственным препаратам.
Бактерии — удобная модель для генетических исследований. Их отличает: относительная простота строения генома, позволяющая выявлять мутанты с частотой 10 -9 и ниже; гаплоидность, исключающая явление доминантности; половая дифференциация в виде донорских и реципиентных клеток; наличие обособленных, и интегрированных фрагментов ДНК (плазмид, транспозонов и т.д.); лёгкость культивирования и возможность получения популяций, содержащих миллиарды микробных тел.
Как и у других организмов, совокупность генов бактериальной клетки — геном — определяет её свойства и признаки (генотип). Фенотип бактериальной клетки — результат взаимодействий между бактерией и окружающей средой — также контролирует геном (так как сами признаки закодированы в бактериальных генах).
Генетический материал бактерий
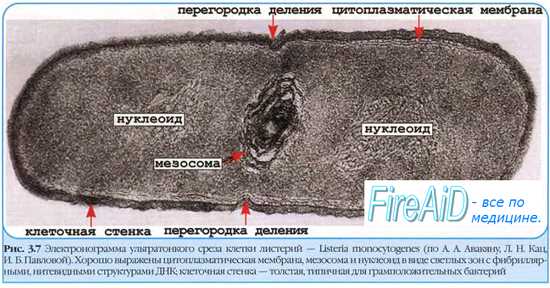
Ядерные структуры бактерий имеют характерное строение, отличающее их от ядер эукарио-тических клеток; их образуют так называемые хроматиновые тельца, или нуклеоиды, лишённые оболочки и включающие в себя почти всю ДНК бактерии.
• Ядерные структуры можно наблюдать в фазово-контрастный микроскоп, где они выглядят как менее плотные участки цитоплазмы. Для их выявления в фиксированных мазках предложена реакция Фёльгена-Россенбёка.
• В растущих бактериальных клетках нуклеоиды активно делятся, их количество иногда достигает 2-4.
Прокариотический геном
У бактерий обычно имеется одна замкнутая кольцевидная хромосома, содержащая до 4000 отдельных генов, необходимых для поддержания жизнедеятельности и размножения бактерий, то есть бактериальная клетка гаплоидна, а удвоение хромосомы обычно сопровождается её делением.
• Некоторые виды (например, Brucella melitensis) стабильно содержат две кольцевые хромосомы, другие (Leptospira interrogans) — одну кольцевую хромосому и одну большую плазмиду, третьи — одну линейную хромосому (Streptomyces ambofaciens), то есть обладают сложными геномами.
• Бактериальная хромосома содержит до 5*10 6 пар оснований. Для сравнения: геном человека составляет 2,9*10 9 пар оснований. Длина бактериальной хромосомы в развёрнутом состоянии составляет около 1 мм (Escherichia coli).
Некоторые бактерии содержат внехромосомные молекулы ДНК (плазмиды) и мобильные элементы (либо плазмидные, либо хромосомные).
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Генетический аппарат бактерий представлен хромосомными (нуклеоид) и внехромосомными (плазмиды, инсерционные последовательности, транспозоны) структурами.
Нуклеоид
не имеет ядерной мембраны и не связан с гистонами;
одна непарная суперспирализованная хромосома состоит из двунитевой молекулы ДНК (кольцевой или линейной) размером от 3х10 8 до 2,9х10 9 Д и содержит до 4600 генов;
один конец бактериальной хромосомы связан с мезосомой;
некоторые бактерии имеют сложные геномы, состоящие из двух или нескольких репликонов.
Структура ДНК и генетический код. Материальной основой наследственности, определяющей генетические свойства всех организмов, является ДНК. Исключение составляют только РНК–содержащие вирусы, у которых генетическая информация закодирована в РНК.
ДНК состоит из последовательности химически связанных нуклеотидов и имеет структуру правильной двойной спирали из закрученных одна вокруг другой двух полинуклеотидных цепей. Каждый нуклеотид состоит из азотистого основания, сахара дезоксирибозы и фосфатной группы. Азотистые основания представлены пуринами (аденин, гуанин) и пиримидинами (тимин, цитозин). ДНК содержит А, С, G, Т; РНК — А, С, G, U.
Каждый нуклеотид обладает полярностью. У него имеются дезоксирибозный 3'–конец и фосфатный 5'–конец. Нуклеотиды соединяются в полинуклеотидную цепочку фосфодиэфирными связями между 5'–концом одного нуклеотида и 3'–концом другого. Соединение между двумя цепочками обеспечивается водородными связями комплементарных азотистых оснований: аденина с тимином, гуанина с цитозином. Нуклеотидные цепи антипараллельны: на каждом конце линейной молекулы ДНК расположены 5'–конец одной цепи и 3'–конец другой цепи. Последовательность нуклеотидов ДНК определяет последовательность аминокислотных остатков в молекуле белка.
Каждому белку соответствует свой ген — уникальная структурная единица наследственности. Ген — фрагмент полинуклеотидной цепи молекулы ДНК, отличающийся числом и специфичностью последовательности нуклеотидов и кодирующий синтез одного пептида. В ДНК содержатся структурные и регуляторные гены.
Регуляторные гены регулируют транскрипцию структурных генов.
Хромосома состоит из особых функциональных единиц — оперонов. Оперон — совокупность промотора, оператора и структурных генов — является функциональной генетической единицей, регулирующей экспрессию одного или группы генов.
Основные этапы развития генетической системы: кодон ген оперон геном вирусов и плазмид хромосома прокариот (нуклеоид) хромосомы эукариот (ядро).
Регуляция выражения генетической информации у бактерий. Бактериальная клетка способна запустить или прекратить синтез фермента в зависимости от присутствия соответствующего субстрата. Для этого бактериальные гены объединены в группы так, что все ферменты, необходимые для осуществления биосинтеза, детерминируются генами, сцепленными друг с другом. Вся группа генов может транскрибироваться в одну полицистронную мРНК, которая последовательно транслируется рибосомами с образованием каждого из белков.
Экспрессия генов у прокариот регулируется на уровне транскрипции. Роль сигнальных веществ для запуска транскрипции играют низкомолекулярные соединения, которые являются либо субстратом для фермента, либо продуктом ферментативной деятельности. Индукция и репрессия представляют собой разные стороны одного и того же явления. Малые молекулы, индуцирующие образование ферментов, способных метаболизировать их, называются индукторами. Те же, которые предотвращают образование ферментов, способных синтезировать их, —корепрессорами.
Молекулы-эффекторы не могут вступать в прямое взаимодействие с ДНК, посредником для них служит специальный регуляторный белок. Регуляторный белок, который связывается с ДНК в отсутствии индуктора, называется репрессором.
За синтез регуляторных белков ответственны регуляторные гены. В присутствии белка-репрессора транскрипция блокирована; его удаление обусловливает доступ РНК-полимеразы к генам и запуск транскрипции. Прекращение синтеза фермента при помощи белка-репрессора получило название репрессии. Репрессия позволяет бактериальной клетке избежать перевода своих ресурсов на ненужную в данный момент синтетическую активность. Если индуктор присутствует в клетке в высокой концентрации, то в результате специфического присоединения к регуляторному белку он изменяет его конформацию и способность связываться с ДНК.
Контроль транскрипции достигается взаимодействием регуляторного белка с регуляторным сайтом (оператором), который расположен между структурными генами и промотором (участком, распознаваемым ДНК-зависимой РНК-полимеразой). Промотор служит местом связывания РНК-полимеразы, и от него начинается транскрипция.
Ген имеет три фундаментальные функции.
1. Непрерывность наследственности — обеспечивается полуконсервативным механизмом репликации ДНК: каждая из двух цепочек ДНК хромосомы или плазмиды служит матрицей для синтеза комплементарной дочерней цепочки ДНК.
Репликация начинается с расплетения двунитевой структуры ДНК ферментом ДНК-гидролазой. При этом формируются две репликативные вилки, которые двигаются в противоположных направлениях, пока не встретятся. Формирование новой дочерней цепи осуществляется ферментом ДНК-полимеразой, присоединяющей комплементарные матрице нуклеотиды к свободному 3'–концу растущей цепочки. Для осуществления реакции полимеризации нуклеотидов на матрице родительской цепочки ДНК-полимеразе требуется затравка, которая называется праймером (англ. primer - запал). Праймер — короткая нуклеотидная цепочка, комплементарная матричной цепочке со свободным 3'–концом.
Две цепи двойной спирали ДНК комплементарны друг другу. На каждой цепи из структурных элементов ДНК (дезоксирибонуклеозидтрифосфатов) синтезируется новая цепь. При этом с каждым из оснований спаривается комплементарное ему основание, так что каждая из двух новых цепей будет комплементарна родительской цепи. Обе новые двойные цепи состоят из одной родительской и одной вновь синтезированной цепи. Такая точная репликация ДНК гарантирует сохранение генетической информации. ДНК бактерий, будучи носителем наследственной информации, сама не служит матрицей для синтеза полипептидов. Биосинтез белков происходит на рибосомах, которые непосредственно с ДНК не соприкасаются. Передачу записанной в ДНК информации к местам синтеза белка осуществляет одноцепочечная мРНК. По последовательности оснований цепь мРНК комплементарна цепи ДНК и отличается от цепи ДНК тем, что тимин в РНК заменен урацилом.
Транскрипция — синтез мРНК на одной из цепей ДНК, начиная с 5'–конца; механизм этого процесса сходен с механизмом репликации ДНК.
Трансляция — перевод нуклеотидной последовательности мРНК в последовательность аминокислот. Аминокислоты затем собираются в полипептидную цепь, т. е. синтезируется белок (рис. 72).

Рис. 72. Биосинтез белка
Последовательность аминокислот определяет пространственную структуру белка — конформацию. По мере продвижения рибосомы вдоль мРНК полипептидная цепь растет, закручивается и свертывается в клубок. В результате возникает структура, обусловливающая специфические свойства и функцию данного белка. К мРНК обычно прикрепляется несколько рибосом, так что на одной и той же матрице одновременно синтезируется несколько полипептидных цепей. На конце мРНК находится кодон, от которого зависит отделение сформированной полипептидной цепи от рибосомы.
2. Взаимоотношения между последовательностью нуклеотидов в гене и последовательностью аминокислот в белке устанавливаются с помощью генетического кода из четырех оснований. Код триплетный, поскольку кодон (функциональная единица, кодирующая аминокислоту) состоит из трех оснований. Последовательности кодонов считываются непрерывно, начиная с фиксированной стартовой точки на одном конце гена и заканчивая точкой терминации на другом конце гена. Это значит, что различные части гена не могут читаться независимо.
Читайте также: