
Азотфиксация это кратко и понятно
Обновлено: 05.07.2024
Знаете ли вы что такое азотфиксация?
Вчера мы говорили о фотосинтезе — процессе, обеспечивающем все живое на Земле такими органическими веществами как углеводы .
Прозорливый ум вправе спросить меня, репетитора по биологии, а откуда же берется на планете для жизни четвертый элемент-органоген азот N ?
Без азота ведь не обеспечишь клетки ни белками, ни нуклеиновыми кислотами — этими биополимерными молекулами, составляющими основу жизни . Немного терпения и мы скоро все узнаем.
Значит, наряду с фотосинтезом на нашей планете должен существовать не менее глобальный процесс, связанный с появлением азота N в живых системах. Да, должен существовать. И он существует! Это биологическая азотфиксация .
Пожалуй нет в биологической науке большего парадокса, чем ее отношение к азоту. Давно известно, что в большом количестве азот на Земле находится лишь в воздухе в виде молекулярного азота N2. В атмосферном воздухе азота содержится около 80%.
Уже в XX веке было показано, что действительно азот может быть пагубен для жизни. Но, конечно, не инертный молекулярный азот воздуха, а нитратный азот NO3, вносимый человеком в почву в виде удобрений под растения в избыточном количестве.
Избыточные нитраты в растениях могут превращаться в нитриты NO2. А нитриты — это действительно сильный яд… Так ли уж был не прав Лавуазье?
Так что же такое азотфиксация
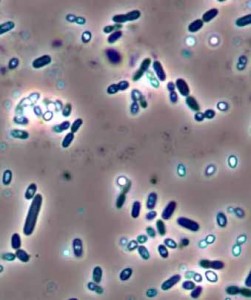
Азотфиксация — это процесс связывания молекулярного азота атмосферы N2 некоторыми бактериями-азотфиксаторами, живущими в почве (на фото клетки бактерии азотобактер). Таким образом, весь связанный почвенный азот, а значит и азот всей биомассы ныне живущих существ на Земле — это результат их многомиллионолетней деятельности.
Как восполнить дефицит белковой пищи? Изобрел человек тогда процесс производства азотных удобрений. А из чего их создавать, где азота больше всего на планете? Конечно из воздуха… Но, что это за процесс? Человек научился связывать молекулярный азот с водородом для производства аммиака NH3 по реакции Габера (изобретенной еще в начале XX века).
Осуществление этого процесса требует затрат огромного количества энергии для поддержания давления в реакционной смеси в 500-1000 атм. и температуры 500 градусов. Габер, великий Габер, заслуженно получивший за свое изобретение Нобелевскую премию, знал бы он, что через 50 лет будет описан процесс азотфиксации, осуществляемый непосредственно в природе бактериями…
Оказалось, что азотфиксирующие бактерии, имеющие специальный фермент нитрогеназу, осуществляют ту же реакцию образования аммиака из молекулярного азота и водорода, но при обычных земных условиях, при давлении 1 атм. и температуре 15-25 градусов.
Это наглядный пример, показывающий насколько рациональна жизнь, насколько биохимические клеточные реакции совершеннее химического катализа.
Связь двух глобальных биосферных процессов фотосинтеза и азотфиксации
Не случайно я вчерашнюю тему по фотосинтезу объединяю с темой азотфиксации, потому что эти два процесса в природе тесно сопряжены.
Оказалось, что частью своих продуктов фотосинтеза, растения делятся с бактериями-азотфиксаторами, поселяющимися в их корневой зоне. За счет энергии углеводов (а получается за счет солнечной энергии) бактерии связывают азот воздуха и кормят им растения (ассоциативная азотфиксация).
Таким образом, растительные белки и нуклеиновые кислоты имеют в своем составе N, полученный бактериями из воздуха.
Всё — круговорот замкнулся. Фотосинтез поставляет в экосистемы углеводы (а это значит С,О и Н), а бактерии азотфиксаторы, особенно активно работающие вблизи растений, доставляют еще и N.

Кроме такой ассоциативной азотфиксации в природе существует и более тесная симбиотическая азотфиксация.
Такой способ существования характерен для бобовых растений и некоторых древесных пород растений. У них азотфиксирующие бактерии поселяются не только вблизи корней, но способны проникать в корни, образуя клубеньки (на фото клубеньки с бактериями на корнях сои).
Именно в клубеньках достигается наибольшая сопряженность процессов азотфиксации и фотосинтеза — углеводы растений не растрачиваются куда попало, а все доходят до адресата, клубеньковых бактерий.
А что в будущем
К сожалению, в своей практической деятельности человек до конца еще не может использовать весь природный потенциал, заложенный в механизме естественной азотфиксации. Здесь, отчасти, и косность мышления, и верность традициям (надо же продолжать нефтегазодобывающим монополиям вбухивать энергетические ресурсы в работу химических комбинатов по производству азотных удобрений, что бы не разориться).
Поэтому человек продолжает плодить по всей Земле заводы-монстры по производству азотных удобрений. Заводы, загрязняющие всё вокруг себя и требующие огромных энергетических затрат невосполнимых природных ресурсов. Но думается, что будущее за энергосберегающими и не наносящими вреда окружающей среде технологиями.
Мне, как репетитору по биологии, бывшему заведующему группой биологическоЙ азотфиксации Института биологии БНЦ УрО АН СССР,
очень хочется верить, что может быть кому то из вас посчастливится найти способ использования в бОльших масштабах природный процесс биологической азотфиксации и научить человечество как избавиться от гигантских заводов-поработителей.
Ваше будущее — в ваших руках, не правда ли?!
У кого есть вопросы по статье к репетитору биологии по Скайпу, замечания, пожелания — прошу в комментарии.
Азотфикса́ция, или азотофиксация — фиксация молекулярного атмосферного азота, диазотрофия. Процесс восстановления молекулы азота и включения её в состав своей биомассы прокариотными микроорганизмами. Важнейший источник азота в биологическом круговороте. В наземных экосистемах азотфиксаторы локализуются в основном в почве.
Содержание
Биохимия
Атомы в молекуле азота связаны прочной тройной ковалентной связью, из-за чего он практически не вступает в реакции окисления-восстановления в нормальных условиях без применения катализаторов и не может использоваться растениями и животными. Микроорганизмы для восстановления азота используют целую серию ферментов (ферредоксин, гидрогеназа), важнейшим из которых является нитрогеназа. За её синтез ответственны так называемые nif-гены, широко распространенные у прокариот (в том числе архебактерий), но не встречающиеся у эукариот. Процесс азотфиксации достаточно энергоёмкий, для ассимиляции 1 молекулы азота требуется не менее 12 молекул АТФ, то есть для использования 1 мг азота анаэробным микроорганизмам требуется около 500 мг сахарозы.
Защита от кислорода
Нитрогеназа блокируется молекулярным кислородом, поэтому азотфиксация в основном анаэробный процесс. Однако ряд аэробных бактерий выработал механизмы защиты нитрогеназы от блокирования:
- Механизм повышенного уровня дыхания. Azotobacter chroococcum при азотфиксации окисляет часть органического вещества, не запасая выделившейся энергии, а только лишь удаляя этим кислород.
- Механизм локализации азотфиксации в гетероцистах характерен для цианобактерий, способных к фотосинтезу с выделением кислорода. Для защиты нитрогеназы от кислорода они имеют особые, лишенные хлорофиллаклетки - гетероцисты. Некоторые цианобактерии, не образующие гетероцисты, также способны к азотфиксации. Нитчатая цианобактерия Plectonema boryanum фиксирует азот в микроаэробных условиях (1.5% содержания кислорода в темноте и 0.5% кислорода на свету), нитчатые цианобактерии Symploca и Lyngbya majuscula, а также одноклеточные цианобактерии родов Gloeothece и Cyanothece способны к азотфиксации при отсутствии освещения.
- Механизм симбиотической защиты характерен для клубеньковых бактерий. В корнях бобовых продуцируется легоглобин, выполняющий функции защиты от избытка кислорода.
Экологические аспекты азотфиксации
Различают три типа азотфиксации:
- Свободноживущими бактериями самых разнообразных таксономических групп.
- Ассоциативная азотфиксация бактериями, находящимися в тесной связи с растениями (в прикорневой зоне или на поверхности листьев) и использующие их выделения (корневые выделения составляют до 30 % продукции фотосинтеза) как источник органического вещества. Азотфиксаторы живут в кишечнике многих животных (жвачные, грызуны, термиты) и человека (род Escherichia).
- Симбиотическая. Наиболее известен симбиоз клубеньковых бактерий (сем. Rhizobiaceae) с бобовыми растениями. Обычно происходит корневое заражение, но известны растения, образующие клубеньки на стеблях и листьях.
История изучения
Первые диазотрофные бактерии были выделены С. Н. Виноградским в 1898 году и названы в честь Луи Пастера Clostridium pasterianum. В 1901 Бейеринк выделил первый аэробный азотфиксатор Azotobacter chroococcum. С. П. Костычев в 1926 на примере азотобактера и растений табака показал существование ассоциативной азотфиксации.
См. также
Литература
Ссылки
- Физиология
- Микробиология
- Биологический цикл азота
Wikimedia Foundation . 2010 .
Полезное
Смотреть что такое "Азотфиксация" в других словарях:
азотфиксация — азотфиксация … Орфографический словарь-справочник
АЗОТФИКСАЦИЯ — связывание молекулярного азота атмосферы и перевод его в азотистые соединения. Биологическая азотфиксация осуществляется клубеньковыми бактериями, живущими в симбиозе с высшими растениями (симбиотическая азотфиксация), а также свободноживущими… … Большой Энциклопедический словарь
АЗОТФИКСАЦИЯ — АЗОТФИКСАЦИЯ, включение азота воздуха в соединение, доступное для использования растениями и другими организмами. Биологическая азотфиксация осуществляется азотфиксирующими микроорганизмами, многие из которых живут в симбиозе с высшими растениями … Современная энциклопедия
Азотфиксация — АЗОТФИКСАЦИЯ, включение азота воздуха в соединение, доступное для использования растениями и другими организмами. Биологическая азотфиксация осуществляется азотфиксирующими микроорганизмами, многие из которых живут в симбиозе с высшими растениями … Иллюстрированный энциклопедический словарь
АЗОТФИКСАЦИЯ — см. в ст. Фиксация азота. Экологический энциклопедический словарь. Кишинев: Главная редакция Молдавской советской энциклопедии. И.И. Дедю. 1989. Азотфиксация усвоение организмами азота и включение его з азотистые соединения. Осущест … Экологический словарь
АЗОТФИКСАЦИЯ — АЗОТФИКСАЦИЯ, в биологии связывание атмосферного азота в химические соединения для потребления живыми организмами. Азотфиксирующие микроорганизмы (главным образом, БАКТЕРИИ и ЦИАНОБАКТЕРИИ) погдощают азот из воздуха, почвы или из воды и… … Научно-технический энциклопедический словарь
АЗОТФИКСАЦИЯ — биологическая, усвоение мол. азота воздуха азотфиксируюшими бактериями с образованием соединений азота, доступных для использования др. организмами. Один из важнейших процессов в круговороте азота в природе; показатель плодородия почвы. А.… … Биологический энциклопедический словарь
азотфиксация — ДИАЗОТРОФЙЯ – усвоение молекулярного азота воздуха азотфиксирующими прокариотными организмами с образованием соединений азота, доступных для использования др. организмами. Осуществляется как свободноживущими азотфиксирующими бактериями… … Словарь микробиологии
азотфиксация — сущ., кол во синонимов: 2 • азотофиксация (1) • усвоение (29) Словарь синонимов ASIS. В.Н. Тришин. 2013 … Словарь синонимов
АЗОТФИКСАЦИЯ — Процесс химического превращения атмосферного газообразного азота в нитраты или аммиак, которые могут использоваться растениями для синтеза аминокислот и других азотсодержащихорганических молекул Словарь бизнес терминов. Академик.ру. 2001 … Словарь бизнес-терминов
азотфиксация — Усвоение молекулярного атмосферного азота микроорганизмами. [ГОСТ 20432 83] Тематики удобрения … Справочник технического переводчика
Азотфиксация, или микробиологическая фиксация атмосферного азота — процесс поглощения микроорганизмами почвы азота атмосферы и трансформация его в органические и минеральные вещества.
Изучением азотфиксации занимались Ж. Буссенго, М. Бейерник, Г. Гельригель, Г. Вильфорт, М.С. Воронин, С.Н. Виноградский, В.Л. Омелянский, Д.Н. Прянишников, Д.И. Менделеев, К.А. Тимирязев.
Навигация
Значение азотфиксации
На долю азота в атмосферном воздухе приходится 78,09%. Над 1 гектаром суши или водной поверхности Земли содержится около 80 тыс. т азота, который недоступен большинству высших растений.
Атомы азота в молекуле N2 соединены очень прочной тройной связью N≡N, поэтому разрыв этой связи сопряжен с большими затратами энергии. В промышленности этот процесс с образованием аамиака происходит при высоких температурах и давлении, тогда как в биологических системах — при нормальном атмосферном давлении и температуре.
В зависимости от источников энергии азотфиксирующие микроорганизмы относят к: автотрофам и гетеротрофам.
По оценкам, суммарный объем азотфиксации в год в наземных экосистемах составляет 175-190 млн т азота, 90-110 млн т из которых приходятся на почвы сельскохозяйственных угодий (Мишустин, 1983). При этом ежегодный вынос азота из почвы с сельскохозяйственной продукцией составляет 110 млн т.
Интенсивность азотфиксации
Опыт, проведенный Б.А. Ягодиным совместно с Ю.Я. Мазелем и Ю.Г. Сазоновым в 1981 г. показал зависимость симбиотической азотфиксации от обеспеченности растений азотом и интенсивности фотосинтеза. В этом опыте, люпин сорта Быстрорастущий 4 выращивали при разных уровнях обеспеченности азота и 1-, 3- и 6-суточном затенении. Освещенность изменялась в 1000 раз. Затенение приводило к снижению азотфиксации, в большей степени — при высоком содержании минерального азота. После 6-суточного затенения азотфиксация в варианте без азота снизилась в 40 раз, в варианте с половинной дозой — полностью прекратилась, при двукратных дозах — азотфиксация остановилась уже после 3-суточного затенения.
Максимум интенсивности азотфиксации отмечался в фазе цветения в вариантах без азота и половинной дозой. В фазе бутонизации при половинной дозе она была больше, чем в варианте без азота. Это объясняется тем, что небольшая стартовая доза азота способствует лучшему развитию клубеньков на ранних этапах развития. В фазе цветения в варианте без азота этот показатель был выше, чем в вариантах с азотом.
В фазе бутонизации максимум азотфиксации в дневном цикле приходился на утренние часы (8 ч), причем в варианте с половинной дозой фиксация проходила быстрее, чем в варианте без азота. В фазе цветения максимум приходился на полдень. В этом случае она была наибольшей в варианте без азота. При повышенной дозе азота этот показатель уменьшался во все фазы развития.
Более интенсивное поступление продуктов, меченных 14 С, отмечалось в варианте без азота. При двойной дозе оно было на 20% меньше. Через 30 мин после экспозиции метка обнаруживалась в клубеньках обоих вариантов (0,37 и 0,07 соответственно, от общей активности). За 2,5 ч в варианте без азота в клубеньки поступило в 7 раз больше продуктов, чем в варианте с азотом, в корни — в 5 раз, в стебли — в 2 раза больше.
Неодинаковая скорость поступления продуктов фотосинтеза в корневые клубеньки при разных уровнях азотного питания повлияла на интенсивность азотфиксации. Вследствие накопления продуктов фотосинтеза в варианте с азотом затенение в течение 3 суток подавило азотфиксацию клубеньков.
Таким образом, затенение люпина приводит к снижению фиксации азота, но в варианте на фоне минерального азота это снижение больше, чем без азота.
Коэффициент азотфиксации составляет от 0,3 до 0,85.
Интенсивность азотфиксации свободноживущими бактериями зависит от запасом легкодоступных органических веществ, служащих источником энергии. Например, активность азотфиксации в прикорневой зоне растений за счет ассоциативной азотфиксации в 3-200 раз больше, чем в почвах междурядий. Поэтому растения является главным фактором деятельности диазотрофных бактерий в ризосфере благодаря корневой экссудации и корнеопада, объем которых составляет от 25 до 50% продукции фотосинтеза.
Интенсивность фиксации азота диазотрофов определяется выделительной деятельностью корневых систем растений, то есть, в конечном счете от фотосинтетической активности.
Высокая активность в ризосфере многих тропических растений связана со способность использовать при фотосинтезе путь С-4-дикарбоновых кислот. Растениям этого типа требуют интенсивного освещения, а максимальная скорость фотосинтеза у них значительно выше, чем у растений, использующих цикл Кальвина (С-3-тип). Так как растениями с С-4-типом расходуется меньшее количество углеводов на фотодыхание, их часть используется для роста корней и корневой экссудации.
Несимбиотическая азотфиксация изучалась многими исследователями, однако о ее масштабах в различных почвенно-климатических зонах информации мало, в связи с тем, что в природных условиях этот процесс зависит от ряда динамичных факторов среды.
Так, согласно ряду исследований плодородных почв рисовых полей показано, что в результате несимбиотической фиксации под рисом накапливаться 60-70 кг/га азота в год. Причем в затопляемых почвах фиксируется 57-63 кг/га азота, а в незатопляемых — 3-7 кг/га, без растений в затопленных почвах — 23-28 кг/га азота.
За 3 месяца вегетации азотфиксация в почвах рисовых полей Краснодарского края составляла 9-27 кг. Внесение соломы в почву способствует размножению различные группы азотфиксирующих бактерий и росту азотфиксации до 20-40 кг/га в месяц. Влажность также способствует усилению активности при разложении соломы и целлюлозы. В интразональных почвах избыточного увлажнения, то есть пойменных, болотных почвах и рисовых плантациях, активность наиболее высока — от 16,5 до 67,5 кг/га в месяц. В почвах тропической зоны несимбиотическая азотфиксация в среднем составляет 200 кг/га в год, достигая иногда 600 кг/га в год.
Активность несимбиотической азотфиксации зависит также: влажности, температуры, гранулометрического состава почвы, степени аэрированности корнеобитаемого слоя, содержания углекислого газа, наличия макро- и микроэлементов. Минеральные удобрения, известкование, воздушный режим также влияют на интенсивность, но, высокая эффективность отмечается, когда влажность, температура и органическое вещество не лимитируют азотфиксацию. Внесение в дерново-подзолистые почвы растительных остатков позволяет увеличить азотфиксирующую активность в 2-5 раз при условии достаточного увлажнения.

Посещение при помощи электронного микроскопа в передаче поперечного сечения клубеньковых бактерий эндофита в корень из сои . Бактерии Bradyrhizobium japonicum (ен) уже решен внутри корней сои и установил симбиотические азотфиксирующие. Это изображение (при большом увеличении) показывает эндоплазматический ретикулум , диктиосому и клеточную стенку .
Это процесс ферментативного восстановления N 2 ( молекулярного азота ) до аммиачного азота или аммиака (NH 3 ): эта комбинированная форма N, называемая ключевым промежуточным соединением, представляет собой конец реакции связывания и начало включения закрепил азот в углеродном скелете. В биологической системе, фиксирующей N 2, оптимальные условия для биологического катализа соответствуют давлению от 0,2 до 1,0 атм N 2 и температуре 30-35 ° C , а условия химического катализа суровые: давление 250-1000 ° C. атм N 2 и температуру 450 ° С .
Резюме
Общий
Этот процесс сравним с фотосинтезом, при котором углеводные вещества образуются из углекислого газа (CO 2 ) в атмосфере. Но в то время как фотосинтез - это работа всех растений (кроме сапрофитных ), фиксация окружающего азота осуществляется только некоторыми видами бактерий, такими как cyanophyceae . Однако многие растения , в основном семейства Fabaceae ( бобовые ), производят его косвенно, в симбиозе с бактериями из их ризосферы , которые обычно расположены в клубеньках, расположенных на их корнях.
До недавнего времени считалось, что микоризные грибы могут фиксировать N 2 . Теперь известно, что свойство фиксировать N 2 строго ограничено прокариотами и никогда не проявлялось у мицелиальных грибов. Фиксацию N 2, измеренную по восстановлению ацетилена ( C 2 H 2 ) до этилена (C 2 H 4 или CH 2 = CH 2 ) микоризным корнем, следует приписывать не самому грибу , а бактериям, связанным с микоризосферой. . Более того, эта связывающая активность N 2 в лесной подстилке даже более важна, чем в самой микоризосфере .
N 2 + 16 АТФ + 8 e - + 8 H + → 2 NH 3 + H 2 ↑ + 16 АДФ + 16 Pi
Ингибирование нитрогеназы O 2 , прямо или косвенно через окислительные радикалы, является сильным регулятором содержания кислорода в атмосфере в кислородном цикле за счет отрицательной обратной связи по фиксации азота. Ингибирование азотфиксации привело бы к падению первичной продуктивности, как непосредственно для диазотрофных цианобактерий, так и косвенно для всей первичной продуктивности гидросферы , вызывая в долгосрочном плане, для круговорота углерода , снижение скорости захоронения. органический углерод. Без захоронения нет чистого образования O 2. биосферой, а избыток кислорода очень реактивными формами постепенно исчезает из-за окисления поверхности планеты .
Азотфиксирующие бактерии
На Земле есть свободные бактерии, которые живут в почве и обеспечивают фиксацию азота либо по отдельности, либо в симбиозе с другими бактериями. В основном это:
Другие бактерии живут в симбиозе с корневой системой растений:
- Rhizobium : бобовые (Fabaceae);
- актиномицетовFrankia : различные виды покрытосеменных, главным образом , деревьев и кустарников, в том числе ольхи , в крушины , в казуариновые и Myrica буре .
В океанах , это цианобактерии , которые фиксируют азот. Есть два типа:
Случай с бобовыми
Различные виды бактерий rhizobium , которые способны инфицировать корни бобовых ( семейство Fabaceae ), специфичны для определенных растений-хозяев. Таким образом, Rhizobium phaseoli поражает бобы ( Phaseolus sp).
В глобальном масштабе зафиксированная таким образом масса азота оценивается в 100 миллионов тонн в год, что по порядку величины соответствует производству азота в химической промышленности.
Fabaceae-rhizobiaceae диалог
Формирование корневых клубеньков включает сложный молекулярный диалог между растением-хозяином и ризобиальными .
Растение выделяет такие сигналы, как флавоноиды , бетаины . Восприятие этих сигналов бактерией вызывает экспрессию множества бактериальных генов, из которых известно около сотни. Наиболее изучены гены nod (или nol и noe). Гены Nod D экспрессируют регуляторные белки в ответ на индукторы растений. После активации белки Nod D связываются с последовательностями узлового бокса, промоторами структурных генов узлов и активируют их транскрипцию. Эти гены продуцируют важные факторы Nod в установлении симбиоза .
Эти факторы NOD активация сигнального пути с участием ряда генов растений - хозяина, и в конечном счете вызывают морфологические изменения и цитологический корень, что приводит к образованию узелков .
Сигнальный путь NOD
Многие гены были идентифицированы у гороха ( Pisum sativum ) , трилистника ( Lotus japonica ) и трилистника люцерны ( Medicago truncatula ) . Все они представляют собой рецепторы SYMRK (рецептор симбиоза, подобный киназе), обладающий киназным доменом на внутриклеточной стороне (RLK, Receptor like kinase) и мотивами, богатыми LysmM (взаимодействие с факторами NOD) на внеклеточной стороне. Эти гены называются Nfr (рецептор фактора Nod) или Sym (гены симбиоза). Рецепторов может быть двух типов: один контролирует заражение корня бактериями, а другой обеспечивает искривление корневых волосков .
Через несколько десятков секунд после применения факторов Nod, быстрый приток кальция наблюдается , который индуцирует деполяризацию в плазматической мембране , ведущей к кривизне и деформации абсорбирующих волосков .
Через десять минут периодические и переходные колебания цитозольного кальция концентрации происходят вблизи перинуклеарной области. Эти кальциевые колебания ( скачки кальция) происходят из-за двух мембранных белков DMI1 и DMI2. Последний представляет собой рецептор типа RKL (рецептор-подобную киназу), представляющий три домена, богатых лейцином (LRR).
Ответ на эти колебания обусловлен геном DMI3, кодирующим кальмодулин и кальций-зависимую протеинкиназу . Этот белок чувств изменения кальция концентрации и путем связывания с кальций- связанных calmodulins и свободный кальций . DMI3 активируется аутофосфорилированием его киназного домена. Кроме того, DMI3 оказывает отрицательную обратную связь на DMI1 и DMI2 и, следовательно, на колебания кальция.
DMI3 активирует экспрессию генов ENOD (ранние гены нодулина), белки , продуцируемых растения в течение первых этапов симбиоза, (такие , как легоглобин) , участвующий в инфекции и органогенезе из узелков активирующих разделения корковых клеток . Однако промежуточные субстраты DMI3 еще не определены.
Эволюция азотфиксирующих симбиозов
Происхождение и внешний вид
Похоже, что rhizobiaceae задействовали микоризный сигнальный путь и что органогенез клубенька происходит из генетической программы вторичного органогенеза корня. Эндосимбиозы, связывающие азот, должны были появиться 65 миллионов лет назад [ ref. желаемый] . У предка, общего для всех этих видов, должен был развиться бактериальный эндосимбиоз благодаря определенной предрасположенности к узловому органогенезу (образование клубеньков является отклонением генетической программы образования вторичных корней). Вероятно, способность образовывать узелки возникала несколько раз и независимо в процессе эволюции . Эта способность также несколько раз терялась, например, при адаптации к более плодородным почвам. Это помогает объяснить присутствие клубеньковых растений в кладе клубеньковых растений.
Специфика
- Первая из этих сил, положительная, - это увеличение количества азота, фиксируемого растением-хозяином, колонизированным азотфиксирующим микроорганизмом.
- Второй, отрицательный, - это заражение растения-хозяина непродуктивным патогеном.
Низкая специфичность, по-видимому, обнаруживается у родов примитивных растений, в то время как специфичность симбионтов увеличивается в процессе эволюции.
На молекулярном уровне во время диалога растения-хозяина ризобий можно определить три уровня специфичности. Гены Nod переносятся бактериальной плазмидой. Затем может происходить горизонтальный перенос генов, который также передает спектры хозяина. Это объясняет взаимодействие между эволюционно далекими видами бактерий и ближайшими растениями-хозяевами, и наоборот. Таким образом, происходит коэволюция между генами симбиоза и растениями-хозяевами, а не между rhizobiaceae и растениями-хозяевами.
Другие растения с ризосферой, фиксирующей азот
У многих растений есть похожие ассоциации:
- Allocasuarina, а также другие роды семействаCasuarinaceae
- Alnus или ольха (семействоBetulaceae )
- Азолла (семейство Azollaceae )
- Casuarina(семействоCasuarinaceae)
- Ceanothus (семейство Rhamnaceae )
- Церкокарпус (семействорозоцветных )
- Cycadacée (семейство Cycadaceae )
- Elaeagnus umbellata (семействоElaeagnaceae )
- Gunnera (семействоGunneraceae )
- Лобария - Lobaria cuneifolia (L.) Haw.; выбрано название: Saxifraga cuneifolia subsp. cuneifolia , Painter's Despair, камнеломка клиннолистная (семействоSaxifragaceae ) с некоторыми лишайниками .
- Myrica (семействоMyricaceae )
- Purshia tridentata (англ. Antelope bitterbrush) (семействорозоцветные )
- Robinia pseudoacacia , Leucaena leucocephala (семействоFabaceae )
- Shepherdia argentea (англ. Серебряный буйвол) (семействоElaeagnaceae )
С 2000 по 2015 год было обнаружено несколько видов растений, демонстрирующих азотфиксирующие эндофиты, в том числе:
- Azoarcus sp (травянистые);
- Сахарный тростник ;
- дикий рис ;
- кукуруза ,
- Сорго халепенсное ;
- Мискантус ;
- Кактус Mammillaria fraileana ;
- Сладкий картофель ;
- бореальные лесные мхи ;
- дюнные травы ( Ammophila arenaria и Elymus mollis ;
- кофейные растения Coffea arabica L .;
- хвойные растения ( Pinus flexilis и Picea engelmannii ), инокулированные Paenibacillus polymyxa .
Способность к связыванию азота была напрямую определена количественно у некоторых трав (сахарного тростника, пшеницы и риса).
Измерение биологической азотфиксации
Это делается двумя способами:
Некоторые исследования в качестве меры предосторожности используют оба этих теста одновременно, например, исследование 2016 года, которое впервые показало существование диазотрофных бактерий-симбионтов первых видов в ветвях деревьев (цветение ив или первых тополей в среде с высокий дефицит азота).
Читайте также: