
Аэробный путь ресинтеза атф кратко
Обновлено: 05.07.2024
После расщепления полимерных липидных молекул полученные мономеры всасываются в верхнем отделе тонкого кишечника в начальные 100 см. В норме всасывается 98% пищевых липидов.
1. Короткие жирные кислоты (не более 10 атомов углерода) всасываются и переходят в кровь без каких-либо особенных механизмов. Этот процесс важен для грудных детей, т.к. молоко содержит в основном коротко- и среднецепочечные жирные кислоты. Глицерол тоже всасывается напрямую.
2. Другие продукты переваривания (длинноцепочечные жирные кислоты, холестерол, моноацилглицеролы) образуют с желчными кислотами мицеллы с гидрофильной поверхностью и гидрофобным ядром. Их размеры в 100 раз меньше самых мелких эмульгированных жировых капелек. Через водную фазу мицеллы мигрируют к щеточной каемке слизистой оболочки. Здесь мицеллы распадаются и липидные компоненты диффундируют внутрь клетки, после чего транспортируются в эндоплазматический ретикулум.
Желчные кислоты также здесь могут попадать в энтероциты и далее уходить в кровь воротной вены, однако бóльшая их часть остается в химусе и достигает подвздошной кишки, где всасывается при помощи активного транспорта.
Аэробный путь ресинтеза АТФ

Аэробный путь ресинтеза АТФ иначе называется тканевым дыханием – это основной способ образования АТФ, протекающий в митохондриях мышечных клеток. В ходе тканевого дыхания от окисляемого вещества отнимаются два атома водорода и по дыхательной цепи передаются на молекулярный кислород, доставляемый в мышцы кровью, в результате чего возникает вода. За счет энергии, выделяющейся при образовании воды, происходит синтез молекул АТФ из АДФ и фосфорной кислоты. Обычно на каждую образовавшуюся молекулу воды приходится синтез трех молекул АТФ.
Чаще всего водород отнимается от промежуточных продуктов цикла трикарбоновых кислот (ЦТК). ЦТК – это завершающий этап катаболизма в ходе которого происходит окисление ацетилкофермента А до углекислого газа и воды. В ходе этого процесса от перечисленных выше кислот отнимается четыре пары атомов водорода и поэтому образуется 12 молекул АТФ при окислении одной молекулы ацетилкофермента А.
В свою очередь ацетилкофермент А может образовываться из углеводов, жиров аминокислот, то есть через это соединение в ЦТК вовлекаются углеводы, жиры и аминокислоты.
Скорость аэробного обмена АТФ контролируется содержанием в мышечных клетках AДФ, который является активатором ферментов тканевого дыхания. При мышечной работе происходит накопление AДФ. Избыток AДФ ускоряет тканевое дыхание, и оно может достигнуть максимальной интенсивности.
Другим активатором ресинтеза АТФ является углекислый газ. Избыток этого газа в крови активирует дыхательный центр головного мозга, что в итоге приводит к повышению скорости кровообращения и улучшению снабжения мышцы кислородом. Максимальная мощность аэробного пути составляет 350 -450 кал/мин-кг. По сравнению с анаэробными путями ресинтеза АТФ тканевое дыхание облает более низкими показателями, что ограничено скоростью доставки кислорода в мышцы. Поэтому за счет аэробной пути ресинтеза АТФ могут осуществляться только физические нагрузки умеренной мощности.
Время развертывания составляет 3 – 4 минуты, но у хорошо тренированных спортсменов может составлять 1 мин. Это связано с тем, что на доставку кислорода в митохондрии требуется перестройка практически всех систем организма.
Время работы с максимальной мощностью составляет десятки минут. Это дает возможность использовать данный путь при длительной работе мышц.
По сравнению с другими идущими в мышечных клетках процессами ресинтеза АТФ аэробный путь имеет ряд преимуществ. 1. Экономичность: из одной молекулы гликогена образуется 39 молекул АТФ, при анаэробном гликолизе только 3 молекулы. 2. Универсальность в качестве начальных субстратов здесь выступают разнообразные вещества: углеводы, жирные кислоты, кетоновые тела, аминокислоты. 3. Очень большая продолжительность работы. В покое скорость аэробного ресинтеза АТФ может быть небольшой, но при физических нагрузках она может стать максимальной.
Однако есть и недостатки.
1. Обязательное потребление кислорода, что ограничено скоростью доставки кислорода в мышцы и скоростью проникновения кислорода через мембрану митохондрий. 2. Большое время развертывания. 3. Небольшую по максимальной величине мощность.
Поэтому мышечная деятельность, свойственная большинству видов спорта, не может быть полностью получена этим путем ресинтеза АТФ. В спортивной практике для оценки аэробного ресинтеза используются следующие показатели: максимальное потребление кислорода (МПК), порог аэробного обмена (ПАО), порог анаэробного обмена (ПАНО) и кислородный приход.
МПК – это максимально возможная скорость потребления кислорода организмом при выполнение физической работы. Чем выше МПК, тем выше скорость тканевого дыхания. Чем тренированнее человек, тем выше МПК. МПК рассчитывают обычно на 1кг массы тела. У людей, не занимающихся спортом МПК 50 мл/мин-кг, а у тренированных людей он достигает 90 мл/мин-кг.
В спортивной практике МПК также используется для характеристики относительной мощности аэробной работы, которая выражается в процентах от МПК. Например, относительная мощность работы, выполняемая с потреблением кислорода 3 л/мин спортсменом, имеющим МПК 6 л/мин, будет составлять 50% от уровня МПК.
ПАО – это наибольшая относительная мощность работы, измеряемая по потреблению кислорода в процентах по отношению к МПК. Большие величины ПАО говорят о лучшем развитии аэробного ресинтеза.
ПАНО – это минимальная относительная мощность работы, также измеренная по потреблению кислорода в процентах по отношению к МПК. Высокое ПАНО говорит о том, что аэробный ресинтез выше в единицу времени, поэтому гликолиз включается при гораздо больших нагрузках.
Кислородный приход – это количество кислорода (сверх дорабочего уровня ), использованное во время выполнения данной нагрузки для обеспечения аэробного ресинтеза АТФ. Кислородный приход характеризует вклад тканевого дыхания в энергообеспечение всей проделанной работы. Кислородный приход часто используют для оценки всей проделанной аэробной работы. Под влиянием систематических тренировок в мышечных клетках возрастает количество митохондрий , совершенствуется кислородно-транспортная функция организма, возрастет количество миоглобина в мышцах и гемоглобина в крови.
Ресинтез липидов в энтероцитах
Активация жирной кислоты
Поступившая в энтероцит (как и в любую другую клетку) жирная кислота обязательно активируется через присоединение коэнзима А. Образовавшийся ацил-SКоА участвует в реакциях синтеза эфиров холестерола, триацилглицеролов и фосфолипидов.
Реакция активации жирной кислоты
Ресинтез эфиров холестерола
Холестерол этерифицируется с использованием ацил-SКоА и фермента ацил-SКоА:холестерол-ацилтрансферазы (АХАТ).
Реэтерификация холестерола напрямую влияет на его всасывание в кровь. В настоящее время ищутся возможности подавления этой реакции для снижения концентрации ХС в крови.
Реакция ресинтеза эфиров холестерола
Ресинтез триацилглицеролов
Для ресинтеза ТАГ есть два пути:
Первый путь, основной – 2-моноацилглицеридный – происходит при участии экзогенных 2-МАГ и ЖК в гладком эндоплазматическом ретикулуме энтероцитов: мультиферментный комплекс триацилглицерол-синтазы формирует ТАГ.
Моноацилглицеридный путь образования ТАГ
Поскольку 1/4 часть ТАГ в кишечнике полностью гидролизуется, а глицерол в энтероцитах не задерживается и быстро переходит в кровь, то возникает относительный избыток жирных кислот для которых не хватает глицерола. Поэтому существует второй, глицеролфосфатный, путь в шероховатом эндоплазматическом ретикулуме. Источником глицерол-3-фосфата служит окисление глюкозы. Здесь можно выделить следующие реакции:
- Образование глицерол-3-фосфата из глюкозы.
- Превращение глицерол-3-фосфата в фосфатидную кислоту.
- Превращение фосфатидной кислоты в 1,2-ДАГ.
- Синтез ТАГ.

Глицеролфосфатный путь образования ТАГ
Ресинтез фосфолипидов
Первый путь – с использованием 1,2-ДАГ и активных форм холина и этаноламина для синтеза фосфатидилхолина или фосфатидилэтаноламина.
Ресинтез фосфолипидов из ДАГ на примере фосфатидилхолина
Второй путь – на основе синтезируемой in situ фосфатидной кислоты.

Синтез и ресинтез видов и селекция
На примере создания аллополиплоидов мы видели, что путем сочетания разных геномов можно синтезировать новые формы типа рафанобрассики, тритикале и других, которые не существуют в природе.
Создание генетическим путем новых форм, не существующих в природе, называют синтезом видов
. После отбора в ряду поколении они становятся вполне константными и могут быть самостоятельными таксономическими единицами.
Наряду с созданием новых форм в генетике исследовалась проблема ресинтеза видов
, т. е. искусственного восстановления уже существующих видов на основе комбинаций геномов при отдаленной гибридизации.
Блестящим доказательством происхождения некоторых, а может быть, и многих видов на основе аллополиплоидии служит то, что путем скрещивания существующих видов удается воспроизвести константные аллополиплоиды, которые возникли в природе в процессе длительной эволюции или путем длительной селекции. Одним из первых такую работу провел шведский генетик А. Мюнтцинг в начале 30-х годов.
В роде Galeopsis (пикульник) имеется несколько видов: G. pubescens (2n = 16), G. speciosa (2n = 16), G. tetrahit (2n = 32) и др. Каждый из этих видов встречается в природе. Мюнтцинг сначала скрестил первые два вида и получил гибрид F1 который был почти стерильным. В F2 было получено лишь одно растение, которое оказалось триплоидом.
Схема ресинтеза вида Galeopsis tetrahit
Этот триплоид имел 2n = 24. Он не был похож ни на одну из родительских форм, но был более сходен с третьим видом G. tetrahit, а потому назван G. pseudotetrahit. Данный аллотриплоид вновь скрестили с одним из родительских видов (G. pubescens). В F3 был получен аллотетраплоид (2n = 32), который по наборам хромосом и внешнему виду оказался сходным с природным видом G. tetrahit и свободно с ним скрещивался. Таким путем в короткий срок на основе сочетания разных геномов удалось ресинтезировать вид G. tetrahit.
Область синтеза новых форм еще мало использована в практических целях для селекции и для изучения происхождения видов, так как дисциплина, которую можно было бы назвать филогенетической генетикой, только начинает формироваться.
Еще в 30-х годах было ресинтезировано несколько естественных видов растений. Так, например, в опытах В. А. Рыбина ресинтезирована культурная слива (Prunus domestica). Рыбин скрестил терн P. spinosa (2n = 32) с алычой P. divaricata (2n = 16); Среди гибридов оказалось одно растение, которое имело, как и P. domestica, 2n = 48 хромосом. Это была константная и плодовитая форма, полностью сходная с домашней сливой.
Болгарский ученый Д. Костов ресинтезировал вид табака Nicotiana tabacum. Путем скрещивания пшеницы Triticum licoccoides (2n = 28) с эгилопсом Aegilops speltoides (2n = 14) удвоения числа хромосом у гибрида первого поколения (21×2) удалось синтезировать формы пшеницы с 2n = 42, сходные имеющимися формами Tr. spelta и хорошо с ним скрещивающиеся.
Перечень подобных примеров ресинтеза видов на основе объединения геномов путем аллополиплоидии можно было бы расширить, конечно, ресинтезированные виды не будут копией естественных дедов, так как последние прошли длительный отбор в течение нескольких десятков, а может быть, и сотен тысяч поколений. Но, идя этим путем, генетик может смело смотреть в будущее, ибо он открывает возможность овладения синтезом новых видов, полезных человеку. И на этом пути мы действительно можем выполнить завет И. В. Мичурина не ждать милости у природы, а реконструировать ее по своему усмотрению.
Итак, отдаленная гибридизация может быть источником изменчивости в процессе видообразования и в селекции.
1. Значение отдаленной гибридизации в видообразовании и создании новых форм зависит от способа размножения скрещивающихся видов.
2. При половом размножении в случае возникновения плодовитых гибридов отдаленная гибридизация создает мощный резерв комбинативной изменчивости для естественного и искусственного набора, поскольку в генотипе гибрида сочетается более разнообразий набор генов, чем при внутривидовом скрещивании.
3. При вегетативном размножении и апомиксисе отдаленные гибриды могут быть константными, завоевывая себе признание в природе и опыте.
4. Отдаленная гибридизация открывает для селекции возможность сочетать в гибриде ряд ценных свойств диких сородичей культурных видов, а в его потомстве при расщеплении получить наиболее продуктивные формы, особенно в отношении устойчивости к неблагоприятным факторам среды и заболеваниям.
5. Отдаленная гибридизация позволяет проводить геномный анализ видов, что открывает возможность генетического анализа филогении видов, чем занимаются филогенетическая генетика и цитогенетика.
7. Отдаленная гибридизация у растений с применением аллополиплоидии служит одним из источников наследственной изменчивости для селекции и видообразования при участии естественного отбора.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Дано определение ресинтеза АТФ. Описаны основные пути ресинтеза АТФ в мышечных волокнах: креатинфосфатный, гликолитический, миокиназный и тканевое дыхание. Описаны количественные критерии путей ресинтеза АТФ, соотношение между различными путями ресинтеза АТФ при мышечной работе, а также между путями ресинтеза АТФ и зонами относительной мощности.

Ресинтез АТФ в мышечных волокнах
Определение
Ресинтез АТФ – синтез АТФ из различных энергетических субстратов во время физической работы в мышечных волокнах.
Формула ресинтеза АТФ выглядит следующим образом:
Пути ресинтеза АТФ
Ресинтез АТФ может осуществляться двумя путями:
- с участием кислорода (аэробный путь).
- без участия кислорода (анаэробный путь);
Аэробный путь (тканевое дыхание, аэробное или окислительное фосфорилирование) – основной способ образования АТФ в мышечных волокнах. Он протекает в митохондриях мышечных волокон. В результате тканевого дыхания выделяется 39 молекул АТФ. Окисляемое вещество распадается до углекислого газа и воды.
Анаэробный ресинтез АТФ
Анаэробный ресинтез АТФ в мышечных волокнах возможен посредством нескольких механизмов:
-
– ресинтез АТФ из креатинфосфата; – ресинтез АТФ из гликогена мышц;
- Миокиназный (аденилаткиназный) ресинтез АТФ – ресинтез АТФ из АДФ при значительном накоплении в мышечных волокнах АДФ. Рассматривается как аварийный механизм, обеспечивающий ресинтез АТФ, когда другие пути ресинтеза АТФ невозможны.
Количественные критерии путей ресинтеза АТФ
Существуют количественные критерии путей ресинтеза АТФ. К ним можно отнести: максимальную мощность, время развертывания, время сохранения или поддержания максимальной мощности, метаболическую ёмкость (табл. 1).
- Максимальная мощность – максимальное количество АТФ, которое может образоваться в единицу времени при функционировании данного пути ресинтеза АТФ.
- Время развертывания – минимальная длительность, необходимая для выхода ресинтеза АТФ на свою максимальную мощность.
- Время сохранения или поддержания максимальной скорости – длительность функционирования данного пути ресинтеза АТФ с максимальной мощностью.
- Метаболическая ёмкость – количество АТФ, которое может образоваться во время мышечной работы за счёт данного пути ресинтеза АТФ.
Таблица 1. Количественные критерии основных путей ресинтеза АТФ (С.С. Михайлов, 2009)
Соотношение между различными путями ресинтеза АТФ
При любой мышечной работе функционируют все три основных механизма ресинтеза АТФ, но включаются они последовательно. В первые секунды ресинтез АТФ осуществляется за счет креатинфосфатной реакции, затем включается гликолиз. По мере продолжения работы на смену гликолизу приходит тканевое дыхание (рис.1). Эта смена механизмов ресинтеза АТФ приводит к уменьшению суммарной выработки АТФ.
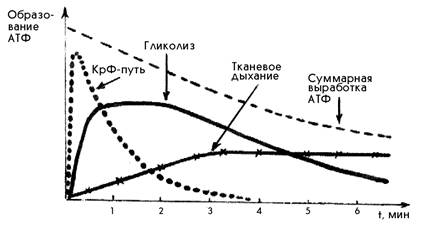
Рис.1. Включение путей ресинтеза АТФ при выполнении физической работы (С.С. Михайлов, 2009)
Пути ресинтеза АТФ и зоны относительной мощности
В.С. Фарфель приводит следующее соотношение мощности работы и основной системы энергообеспечения (табл.2)
Таблица 2. Зоны мощности работы и основная система энергообеспечения (В.С. Фарфель)
| Мощность работы | Основная система энергообеспечения | Типичное время работы |
| Максимальная | Креатинфосфатная реакция | до 20 с |
| Субмаксимальная | Гликолиз | до 5 мин. |
| Большая | Гликолиз+ тканевое дыхание | до 30 мин. |
| Умеренная | Тканевое дыхание | Более 30 мин. |
J.T. Cramer (2008) приводит несколько иное соотношение зон мощности и основных систем энергообеспечения (табл.3)
Таблица 3. — Зоны относительной мощности и основная ситема энергообеспечения (J.T. Cramer, 2008)
| % от максимальной мощности работы | Основная система энергообеспечения | Время работы |
| 90-100 | Креатинфосфатная реакция | 5-10 с |
| 75-90 | Гликолиз | 15-30 с |
| 30-75 | Гликолиз+ тканевое дыхание | 1-3 мин. |
| 20-30 | Тканевое дыхание | Более 3 мин. |
Литература
- Михайлов С.С. Спортивная биохимия. – М.: Советский спорт, 2009.– 348 с.
- Волков Н.И., Несен Э.Н., Осипенко А.А., Корсун С.Н. Биохимия мышечной деятельности.- Киев: Олимпийская литература, 2000.- 504 с.
Похожие записи:
Тест времени реакции на сигнал
Представлена программа расчета времени реакции на сигнал, предназначенная для использования в учебных целях, например на занятиях по…
Саркоплазматическая гипертрофия мышц
Дано определение и описаны механизмы саркоплазматической гипертрофии скелетных мышц. Показано, что этот вид гипертрофии мышц широко…
Классификация типов конституции человека М.В. Черноруцкого
Рассмотрена классификация типов конституции человека, разработанная выдающимся терапевтом М.В. Черноруцким в 1925 году. Классификация типов конституции человека М.В.
Типы гипертрофии скелетных мышц человека
В статье дается классификация различных видов гипертрофии скелетных мышц человека на основе ряда классификационных признаков: времени проявления…
Миомейкер: Мембранный активатор слияния миобластов и образования мышц
Ученые установили, что для образования мышечных волокон необходимо слияние клеток-предшественников, которые называются миобластами. Эти клетки имеют только…
Время развёртывания составляет 3-4 минуты (у хорошо тренированных спортсменов может быть около 1 минуты). Такое большое время развёртывания объясняется тем, что для обеспечения максимальной скорости тканевого дыхания необходима перестройка всех систем организма, участвующих в доставке кислорода в митохондрии мышц [11].
Время работы с максимальной мощностью (на уровне МПК) составляет десятки минут, но в связи с тем, что максимальная мощность данного механизма ограничена на уровне 350-450 кал/мин*кг, только за счёт аэробного пути ресинтеза АТФ невозможно выполнение физических нагрузок типа подтягиваний на перекладине.
Аэробный тренинг
Аэробный тренинг Я рекомендую всем тем, кто считает занятия аэробными тренировками пустой тратой времени, а уж тем более не считает нужным еще и тратить время на их изучение, все-таки прочитать и вникнуть в изложенный ниже материал с таким же фанатизмом и огромным
2.3 БИОЭНЕРГЕТИКА ПОДТЯГИВАНИЙ. 2.3.1 Пути ресинтеза АТФ
2.3 БИОЭНЕРГЕТИКА ПОДТЯГИВАНИЙ. 2.3.1 Пути ресинтеза АТФ Для выполнения как динамической так и статической работы требуется энергия. Непосредственным источником энергии для мышечных сокращений является расщепление высокоэнергетического вещества аденозинтрифосфата
2.3.1.1 Креатинфосфатный механизм ресинтеза АТФ.
2.3.1.1 Креатинфосфатный механизм ресинтеза АТФ. В мышечных клетках всегда имеется креатинфосфат – соединение, обладающее большим запасом энергии и легко отдающее её при взаимодействии с молекулами АДФ, появляющимися в мышечных клетках при физической работе в результате
2.3.1.2 Гликолитическии механизм ресинтеза АТФ.
2.3.1.2 Гликолитическии механизм ресинтеза АТФ. Обеспечивает ресинтез АТФ за счет анаэробного (бескислородного) расщепления глюкозы и гликогена (гликолиз и гликогенолиз) с образованием молочной кислоты (лактата). Данный механизм работает в тех случаях, когда
6.1.3 Источники энергии для аэробного ресинтеза АТФ.
6.1.3 Источники энергии для аэробного ресинтеза АТФ. Длительность поддержания аэробной работы заданного уровня мощности зависит от запасов в организме доступных источников энергии – энергетических субстратов, т.е. тех веществ, которые могут подвергаться окислению
6.1.6 Уменьшение времени развёртывания механизма аэробного ресинтеза АТФ.
6.1.6 Уменьшение времени развёртывания механизма аэробного ресинтеза АТФ. Время развёртывания – это минимальное время, необходимое для выхода ресинтеза АТФ на свою наибольшую скорость, т.е. для достижения максимальной мощности. Время развёртывания аэробного ресинтеза
7.2 Строение мышечных волокон и механизм мышечных сокращений
7.2 Строение мышечных волокон и механизм мышечных сокращений Статическая сила, динамическая сила, статическая силовая выносливость, динамическая силовая выносливость… - физические качества, уровень развития которых определяет спортивный результат в
7.2.2 Механизм мышечного сокращения.
7.2.2 Механизм мышечного сокращения. В соответствии с теорией скользящих нитей мышца сокращается в результате укорочения множества последовательно соединенных саркомеров в миофибриллах, при этом тонкие актиновые нити скользят вдоль толстых миозиновых, двигаясь между
Глава 8. Аутолиз, или Механизм самопереваривания
Глава 8. Аутолиз, или Механизм самопереваривания По мысли А. М. Уголева, аутолиз, или механизм самопереваривания, является одним из наиболее древних. Если попытаться кратко его описать, то можно сказать, что пищеварительные ферменты хозяина (то есть самого организма)
Механизм очищения организма при голодании
Механизм очищения организма при голодании Сперва из организма уходят застоявшиеся воды, поваренная соль, соли кальция. Это происходит на первом этапе голодания. Затем начинают потребляться патологически деформированная ткань, внутриполостной жир, мышечная ткань. Это
Механизм дыхания
Механизм дыхания Механизм дыхания подразделяется на две фазы: вдох, то есть ввод воздуха внутрь легочного пространства, и выдох, то есть выход воздуха из легких.Вдох является результатом активного мышечного действия. Самой главной мышцей для производства каждого вдоха
Спусковой механизм, предохранитель и магазин
Спусковой механизм, предохранитель и магазин Спусковой крючок очень хорошо подходит к указательному пальцу не в последнюю очередь, благодаря превосходной пистолетной рукоятке. Спусковой механизм с предупреждением при минимальном предварительном ходе срабатывает
АТФ в процессе сокращения поставляет необходимую энергию для образования актомиозинового комплекса, а в процессе расслабления мышцы — обеспечивает энергией активный транспорт ионов кальция в ретикулум. Для поддержания сократительной функции мышцы концентрация АТФ в ней должна находиться на постоянном уровне от 2 до 5 ммоль/кг.
Поэтому при мышечной деятельности аденозинтрифосфорная кислота должна восстанавливаться с той же скоростью, с какой расщепляется в процессе сокращения, что осуществляется отдельными биохимическими механизмами ее ресинтеза.
Энергетические источники ресинтеза АТФ в скелетных мышцах и других тканях — богатые энергией фосфатсодержащие вещества. Они присутствуют в тканях (креатинфосфат, аденозиндифосфат) или образуются в процессе катаболизма гликогена, жирных кислот и других энергетических субстратов. Кроме того, в результате аэробного окисления различных веществ возникают энергии протонного градиента на мембране митохондрий.
Ресинтез аденозинтрифосфата может осуществляться в реакциях без участия кислорода (анаэробные механизмы) или с его участием (аэробный механизм). В обычных условиях ресинтез АТФ в мышцах происходит преимущественно аэробным путем. При напряженной физической работе, когда доставка кислорода к мышцам затруднена, включаются и анаэробные механизмы ресинтеза АТФ. В скелетных мышцах человека выявлены три вида анаэробных и один путь аэробного восстановления аденозинтрифосфата.
К анаэробным механизмам относятся креатинфосфокиназный (фосфогенный или алактатный), гликолитический (лактатный) и миокиназный механизмы.
Аэробный механизм ресинтеза АТФ заключается в окислительном фосфорилировании, протекающем в митохондриях, количество которых в скелетных мышцах при аэробных тренировках существенно увеличивается. Энергетическими субстратами аэробного окисления служат: глюкоза, жирные кислоты, частично аминокислоты, а также промежуточные метаболиты гликолиза (молочная кислота) и окисления жирных кислот (кетоновые тела).
Каждый механизм имеет разные энергетические возможности, которые оцениваются по следующим критериям: максимальная мощность, скорость развертывания, метаболическая емкость и эффективность.
Максимальная мощность — это наибольшая скорость образования АТФ в данном метаболическом процессе. Она лимитирует предельную интенсивность работы, выполняемой за счет используемого механизма.
Скорость развертывания — время достижения максимальной мощности данного пути ресинтеза адено-зинтрифосфата от начала работы.
Метаболическая емкость — общее количество АТФ, которое может быть получено в используемом механизме ресинтеза АТФ за счет величины запасов энергетических субстратов. Емкость лимитирует объем выполняемой работы. Метаболическая эффективность — это та часть энергии, которая накапливается в макроэргических связях аденозинт-рифосфата. Она определяет экономичность выполняемой работы и оценивается общим значением коэффициента полезного действия, представляющего отношение всей полезно затраченной энергии к ее общему количеству, выделенному при текущем метаболическом процессе.
Общий коэффициент полезного действия при преобразовании энергии метаболических процессов в механическую работу зависит от двух показателей:
- эффективности фосфорилирования;
- эффективности хемомеханического сопряжения (эффективности преобразования АТФ в механическую работу).
Эффективность хемомеханического сопряжения в процессах аэробного и анаэробного метаболизма примерно одинакова и составляет 50%.
Эффективность фосфорилирования наивысшая в алактатном анаэробном процессе — около 80%, и наименьшая в анаэробном гликолизе — в среднем 44%. В аэробном же процессе она составляет примерно 60%.
Таким образом, анаэробные механизмы имеют большую максимальную мощность и эффективность образования АТФ, но короткое время удержания и небольшую емкость, из-за малых запасов энергетических субстратов. Например, максимальная мощность креатинфосфокиназной реакции развивается уже на 0,5—0,7 с интенсивной работы и поддерживается 10-15 с у нетренированных людей идо 25-30 су высокотренированных спортсменов и составляет 3,8 кДж/кг в минуту.
Гликолитический механизм ресинтеза АТФ отличается невысокой эффективностью. Большая часть энергии остается в молекулах образующейся молочной кислоты. Концентрация последней находится в прямой зависимости от мощности и продолжительности работы, и может быть выделена только путем аэробного окисления.
Гликолиз — это основной путь энергообразования в упражнениях субмаксимальной мощности, предельная продолжительность которых составляет от 30 с до 2,5 мин (бег на средние дистанции, плавание на 100 и 200 м и др.).
Гликолитический механизм энергообразования служит биохимической основой специальной скоростной выносливости организма.
Миокиназная реакция происходит в мышцах при значительном увеличении концентрации АДФ в саркоплазме. Такая ситуация возникает при выраженном мышечном утомлении, когда другие пути ресинтеза уже не возможны.
Таким образом, анаэробные механизмы являются основными в энергообеспечении кратковременных упражнений высокой интенсивности.
При адаптации к интенсивным нагрузкам повышается активность ферментов анаэробных механизмов и запасов энергетических механизмов: содержание креатинфосфата в скелетных мышцах может увеличиваться в 1,5—2 раза, а содержание гликогена — почти в 3 раза.
Читайте также: