
Роль атипических митозов в патологии человека кратко
Обновлено: 04.07.2024
Важное и органическое свойство всех живых организмов — размножение или воспроизведение себе подобных.
Живая материя предстает в виде элементарных единиц на любом уровне организации. Проще говоря, она является дискретной. Соответственно, дискретность — одно из многочисленных свойств живого организма.
Структурные единицы клетки — органоиды.
Целостность клетки зависит от непрерывного воспроизведения новых органоидов взамен тем, что износились. Любой организм состоит из клеток, размножение которых обеспечивает его развитие и существование.
Почему ядро и клетки делятся
Деление клеток — основа размножения. При этом деление ядра всегда происходит до деления клетки. Предположительно, ядро и другие органеллы клетки возникли в результате специализации и дифференциации отдельных участков цитоплазмы в ходе исторического развития. Но в процессе индивидуального развития клеток, ядро происходит только из ядра — в результате деления.
Растительный организм растет, то есть, увеличивается в размерах, в результате увеличения количества клеток путем деления. Если организм одноклеточный, то деление клетки в нем является еще и способом размножения: увеличивается вес и количество новых особей.
Клетка растет определенный промежуток времени. В ходе ее роста изменяется соотношение между растущим объемом клеток и ее растущей поверхностью.
Рост поверхности всегда меньше в абсолютном выражении, чем рост объема. Все потому, что поверхность увеличивается квадратически, а объем — кубически.
Способы деления клетки
Питание клетки осуществляется через поверхность. В определенный момент времени поверхность не может сохранить объем клетки, и она начинает активно делиться.
Клетка делится 4 различными способами:
- Амитозом.
- Митозом.
- Эндомитозом.
- Мейозом.
Какова биологическая роль митоза, мейоза и эндомитоза, мы поговорим позже. А сейчас разберемся с амитозом.
Особенности амитоза
Амитоз — прямое деление ядра, происходящее, когда ядерное вещество перестраивается без образования хромосом.
Виды амитозного распределения
Существенное отличие амитоза от митоза в том, что в первом случае конденсации хромосом не происходит: хромосомы удваиваются. Также для амитоза характерно сохранение физико-химических свойств цитоплазмы.
Согласно физиологическому значению, выделяют три вида амитозного распределения:
- Генеративный амитоз. Клетки делятся полноценно. Их дочерние клетки способны к митозному распределению и стандартному функционированию.
- Реактивный амитоз. Происходит в результате неадекватных действий, оказывающих влияние на организм.
- Дегенеративный амитоз. Такое распределение характеризуется процессом разрушения и смертью клетки.
Процесс амитоза
Амитозное деление клетки в результате расщепления ядра происходит цитоплазматическое сужение. Вначале ядро удлиняется, а после приобретает вид гантелей. При дальнейшем сужении ядро делится на два ядра. После деления ядра происходит деление цитоплазмы, которая делит клетку на две одинаковые (или примерного одинаковые) половины.
Никакого ядерного события не происходит — но образуются две дочерние клетки. Клетка увеличивается за счет ауксентического роста. Происходит расширение ядра, которое приобретает структуру в виде гантели с медианным сужением.
Срединная часть клеточной мембраны приобретает два сужения. Сужение ядра становится все глубже и в конечном счете делит ядро на два дочерних ядра — шпиндельное волокно при этом не образуется. Инвагинации клетки смешаются внутрь. Родительская ячейка делится на пополам: на две равных по размеру дочерние клетки.
Амитоз можно наблюдать у молодых нормально развитых клеток. Хотя чаще всего так делятся высокодифференцированные и более старые клетки. Путем амитоза делятся низкоуровневые организмы, такие как дрожжи, бактерии и пр.
Минус амитоза — невозможность генетической рекомбинации и вероятность экспрессии нежелательных рецессивных генов.
Значение амитоза
Плюс амитоза в том, что ядро и содержимое клетки делятся на две части. При этом дочерние клетки образуются без предварительных изменений структуры органелл, а также ядра.
Деление ядра на две части происходит даже без предварительного растворения оболочки ядра. Веретено деления не формируется — в случае других способов деления это происходит.
За делением ядра происходит деление протопласта и всей клетки на две части. Если ядро дробится на несколько частей, то получаются многоядерные клетки. Для амитоза характерно неравномерное распределение вещества ядра между дочерними ядрами, поэтому биологическая равномерность не обеспечивается. Но при этом образованные клетки отличаются структурной организацией и жизнедеятельностью.
На протяжении некоторого времени амитоз считался патологическим явлением — присущим только клеткам, патологически измененным. Но согласно последним исследованиям, эта мысль не нашла подтверждения. Многочисленные исследования доказывают, что способом амитоза делятся и вполне молодые нормально развитые клетки.
К примеру, амитоз можно обнаружить в клетках харовых водорослей, лука, традесканции.
Встречается амитоз и в специализированных тканях, в которых активно происходят метаболические процессы: в клетках тапетума микроспорангиев, в эндосперме семян отдельных растений и др.
Не характерен амитоз для клеток, в которых важно сохранение полноценной генетической информации: в яйцеклетках и клетках зародыша. В связи с этим некоторые ученые считают амитоз неполноценным способом размножения клеток.
Патология митоза развивается при нарушении нормального течения митотического деления и зачастую приводит к возникновению клеток с несбалансированными кариотипами, следовательно, ведёт к развитию мутаций и анеуплоидии. Также в результате развития отдельных форм патологии наблюдаются хромосомные аберрации. Незавершённые митозы, прекращающиеся по причине дезорганизации или разрушения митотического аппарата приводят к образованию полиплоидных клеток. Полиплоидия и формирование дву- и многоядерных клеток возникают в случае нарушений механизмов цитокинеза. При значительных последствиях патологии митоза возможна гибель клетки.
В нормальных тканях патология встречается в незначительных количествах. Например, в эпидермисе мышей встречается около 0,3 % патологических митозов; в эпителии гортани и матки человека — около 2 %. Патологические митозы часто наблюдаются при канцерогенезе, при различных экстремальных воздействиях, при лучевой болезни или вирусной инфекции,[~ 8] при раке и предраковых гиперплазиях.[~ 9] Частота патологических митозов также увеличивается с возрастом.[73]
Условно различают патологию митоза функционального и органического типа. К функциональным нарушениям относят, например, гипореактивность вступающих в митоз клеток — снижение реакции на физиологические регуляторы, определяющие интенсивность пролиферации нормальных клеток. Органические нарушения возникают при повреждении структур, участвующих в митотическом делении (хромосомы, митотический аппарат, клеточная поверхность), а также при нарушении процессов, связанных с данными структурами (репликация ДНК, образование веретена деления, движение хромосом, цитокинез).[73]
Классификация и общая характеристика различных форм патологии митоза
На основании морфологических признаков и цитохимических нарушений митотического процесса выделяют три основных группы патологии митоза: патология, связанная с повреждением хромосом; патология, связанная с повреждением митотического аппарата; нарушение цитокинеза.[74]
I. Патология митоза, связанная с повреждением хромосом
1) Задержка митоза в профазе наблюдается при нарушениях репликации ДНК.
2) Нарушение спирализации и деспирализации хромосом прослеживается в результате действия на делящуюся клетку различными митотическими ядами. Например, воздействие колхицина приводит к гиперспирализации хромосом, которые приобретают укороченную и утолщенную форму.[74]
3) Раннее (преждевременное) разделение хроматид в профазе (в норме разделение хроматид происходит на рубеже перехода метафазы в анафазу). Обозначенная патология наблюдается, к примеру, при изменении осмотического давления в фибробластах кролика в культуре ткани или же при воздействии канцерогенов (бензопирена, метилхолантрена) на мышиные фибробласты.[74]
В центральной делящейся клетке, находящейся в стадии телофазы, наблюдается парный фрагмент. Микрофотография сделана во время проведения эксперимента Allium test
В центре клеточного поля видна делящаяся клетка в стадии анафазы. Отчётливо заметен хроматидный мост и одиночный фрагмент хромосомы. Микрофотография сделана во время проведения эксперимента Allium test по изучению влияния активного излучения сотового телефона на клетки in vivo
4) Фрагментация и пульверизация хромосом возникает в опухолевых клетках, при вирусной инфекции, в результате воздействия на нормальные клетки ионизирующего излучения или мутагенов. Фрагменты могут быть одиночными, парными и множественными. Те из них, которые лишены центромерного участка, не участвуют в метакинезе, и, соответственно, не расходятся к полюсам деления в анафазе. При массовой фрагментации хромосом (пульверизация) большинство фрагментов также беспорядочно рассеиваются в цитоплазме и не участвуют в метакинезе.[75]
В итоге часть фрагментов хромосом может попасть в одно из дочерних ядер, либо резорбироватья, либо образовать обособленное микроядро. Также отдельные фрагменты обладают способностью воссоединяться своими концами, причём подобные воссоединения носят случайный характер и приводят к хромосомным аберрациям.[76]
5) Хромосомные и хроматидные мосты являются следствием фрагментации хромосом. При воссоединении фрагментов содержащих центромер образуется дицентрическая хромосома, которая в ходе анафазы растягивается между противоположными полюсами деления, образуя мост. Хромосомный (обычно двойной) мост возникает в результате воссоединения фрагментов хромосом, каждый из которых образован двумя хроматидами с центромерой. Хроматидный (обычно одиночный) мост возникает в результате воссоединения двух фрагментов отдельных хроматид с центромерой.[77]
К концу анафазы — в начале телофазы мосты обычно быстро рвутся в результате чрезмерного растягивания дицентрических фрагментов хромосом. Образование мостов приводит к генотипической разнородности дочерних клеток, а также нарушает течение завершающих стадий деления и задерживает цитокинез.[77]
7) Образование микроядер происходит вследствие фрагментации или отставания отдельных хромосом, вокруг которых в телофазе формируется ядерная оболочка, параллельно образованию оболочки вокруг основных дочерних ядер. Новообразованные микроядра либо сохраняются в клетке в течение всего дальнейшего клеточного цикла вплоть до очередного деления, либо подвергаются пикнозу, разрушаются и выводятся из клетки.[78]
8) При нерасхождении хромосом сестринские хроматиды не разъединяются с началом анафазы и вместе отходят к одному из полюсов, что приводит к анеуплоидии.[79]
9) Набухание и слипание хромосом наблюдается в опухолевых клетках и при воздействии токсических доз различных митотических ядов. Вследствие набухания хромосомы теряют свои нормальные очертания и слипаются, превращаясь в комковатые массы. Расхождения хромосом не происходит и клетки в таком состоянии зачастую погибают.[79]
II. Патология митоза, связанная с повреждением митотического аппарата
1) Задержка митоза в метафазе характерна для всей группы патологий митоза, связанных с повреждением митотического аппарата.
2) Колхициновый митоз или к-митоз — одна из форм патологии митоза, связанная с повреждением митотического аппарата вследствие воздействия статмокинетических ядов (колхицина, колцемида, винбластина, винкристина, аценафтена, метанола и др.).[80] В результате воздействия статмокинетических ядов митоз задерживается на стадии метафазы в связи с дезорганизацией различных компонентов митотического веретена деления — центриолей, микротрубочек, кинетохоров. Повреждения также затрагивают клеточное ядро, плазмалемму, различные внутриклеточные органоиды (митохондрии, хлоропласты, аппарат Гольджи). Действие статмокинетических ядов усиливает спирализацию хромосом, что приводит к их укорочению и утолщению, а порой доводит до набухания и слипания хромосом. Как следствие, происходят хромосомные аберрации, образуются микроядра в результате фрагментации или отставания хромосом, развивается анеуплоидия.[81]
Исход к-митоза зависит от дозы и времени воздействия статмокинетического яда на делящуюся клетку. При токсических дозах наблюдается пикноз ядра и гибель клетки. Значительные отравления приводят к полиплоидизации. Воздействие небольших доз обратимо. В течение нескольких часов может восстановиться митотический аппарат и продолжиться митотическое деление.[81]
3) Рассеивание хромосом в метафазе происходит в результате повреждения или полной дезорганизации митотического аппарата.
Трёхполюсный митоз в клетке рака молочной железы
4) Многополюсный митоз связан с аномалией репродукции центриолей, что ведет к формированию дополнительных полюсов и веретен деления. В итоге хромосомы распределяются неравномерно между дочерними ядрами, что в свою очередь ведет к образованию анеуплоидных клеток с несбалансированным набором хромосом.[82]
5) Моноцентрический митоз связан с нарушением разделения центриолей. При этом формируется лишь один полюс, от которого расходятся нити единственного полуверетена. В итоге моноцентрический митоз приводит к полиплоидизации.[83]
6) Асимметричный митоз характеризуется непропорциональным развитием противоположных полюсов деления, что приводит к неравномерному распределению хромосом между дочерними ядрами, то есть к анеуплоидии.[83] В результате асимметричный митоз приводит к образованию микроклеток и гигантских клеток с гипо- и гиперплоидными ядрами.
8) Полая метафаза представляет собой кольцевое скопление хромосом в экваториальной пластинке вдоль периферии клетки.[85]
III. Патология митоза, связанная с нарушением цитотомии
Различают две группы патологии митоза, связанные с нарушением цитотомии: раннюю цитотомию, берущую начало еще в анафазе; либо наоборот, запаздывание или полное отсутствие цитотомии, в результате чего формируются двуядерные клетки, либо образуется одно полиплоидное ядро
На протяжении клеточного цикла центриоли и центросома претерпевают структурные изменения, обусловленные выполняемыми ими в зависимости от стадии функциями. В связи с этим возникло понятие о центриолярном и центросомном циклах. Имеющиеся в литературе данные позволяют говорить о параллельном и независимом от редупликации ДНК их течении. Остановимся поподробнее на цикле редупликации центриолей.
В настоящее время можно считать твердо установленным тот факт, что начало репликации ДНК не совпадает с началом репродукции центриолей. Этот процесс в различных клетках сдвинут в большей или меньшей степени к середине S-периода. Первая стадия образования центриолей – инициация. На этом этапе происходит закладка новой центриоли, иначе называемой процентриолью. На первом этапе образования процентриоль состоит из 9 одиночных микротрубочек, и лишь затем формируются дуплеты и триплеты.
Далее, в течение S-периода, осуществляется удлинение процентриоли – элонгация. Практически во всех гепатоцитах на этой стадии у половины материнских центриолей были обнаружены сателлиты (от 1 до 3), имеющие головку, от которой могут отходить микротрубочки. Длина дочерней центриоли в гепатоцитах регенерирующей печени мыши составляет 1/2 длины материнской центриоли, в клетках культуры 3Т3 – 1/3, в клетках культуры СПЭВ – 1/2.
Рост центриоли продолжается и в G2-периоде; теперь её длина составляет 3/4 от размера материнской центриоли гепатоцита (или, например, 2/3 – для клеток щитовидной железы). Все центриоли на этой стадии формируют диплосомы, сателлиты обнаруживаются лишь изредка и при этом лишены головки.
Терминация роста дочерней центриоли обычно наблюдается к митозу: в клетках культуры СПЭВ и 3Т3 в прометафазе, в эпителии щитовидной железы в телофазе или даже в раннем G1-периоде.
Таким образом, дупликация центриолей идет по полуконсервативному механизму, заключающемуся в разъединении диплосомы и формировании новой центриоли в паре со старой. Вопрос о том, почему для образования новой, дочерней, центриоли необходима старая, материнская, центриоль, пока еще остается открытым. Вероятно, материнская центриоль обладает специфическими структурами, принимающими участие в этом процессе. С другой стороны, в литературе существуют данные, подтверждающие возможность образования центриолей de novo, в отсутствие материнской центриоли.
Следует также отметить, что дупликация центриолей, как и репликация ДНК, зависит от активности циклин Е-Cdk2, а также от циклин А-Cdk2. Ингибирование комплекса циклин Е-Cdk2 при помощи р27 блокирует образование центриолей.
Процессы репликации центриолей и расхождения к полюсам во время митоза обеспечивают поддержание постоянного числа центриолей в клетке. При анализе ультраструктуры клеток печени мыши было показано, что число центриолей соответствует плоидности клетки. Вне зависимости от их количества центриоли формируют в клетке единый комплекс, названный центриолярным.
Преобразования, наблюдаемые в центриолярном цикле, связаны с морфологическими изменениями перицентриолярного материала. Морфофункциональные преобразования центросомы в клеточном цикле составляют центросомный цикл. Перицентриолярные сателлиты исчезают в G2-периоде, и вместо них около обеих материнских центриолей начинает образовываться фибриллярное гало – центр организации микротрубочек веретена деления. Ядерный и центросомный циклы синхронизируются в сверочных точках (checkpoints), что обеспечивает своевременную редупликацию ДНК и центриолей.
Аппарат движения.
Терминами базальное тело, кинетосома, блефаропласт обозначают органеллу, из которой развивается ресничка или жгутик. Как оказалось, все они во многих случаях имеют строение. Сходное со строением центриоли. Это дало полное основание Вольфу (1972) назвать базальное тело ресничной центриолью.
Цитофизиология митоза.
Кинетохор.
1.1.1. Виды кинетохора:
Ø Диффузный кинетохор – располагается по всему телу хромосомы. Выявлен у круглых червей, прямокрылых насекомых, однодольных растений.
Ø Локализованный кинетохор – привязан к центромерному району. Это основной вариант кинетохора в растительных и животных клетках.
Следует обратить внимание, что трехслойная структура кинетохора обнаруживается только в метафазе.
1.1.3. Белки кинетохора
CENP – centromere protein. Это семейство белков кинетохора. Все белки кинетохора делят на 2 группы: конститутивные – постоянно входящие в состав кинетохора (CENP A, CENP B, CENP C, CENP D) и те, которые появляются только во время митоза (CENP E, CENP F).
Так например, CENP F – белок ядерного матрикса: в интерфазе он располагается в ядре, а при вступлении клетки в митоз – на кинетохорах.
Цитокинез.
Силу, необходимую для цитокинеза, создают актин и миозин. Разделение цитоплазмы происходит под действием кольца, состоящего, главным образом, из актиновых филаментов и затягивающегося, подобно петле, вокруг средней части клетки. Этот пучок филаментов, называемый сократимым кольцом, должен быть как-то прикреплен к внутренней стороне плазматической мембране.
Не известно, каким способом в начале анафазы образуется сократимое кольцо. Сформировавшееся кольцо можно преждевременно активировать путем электростимуляции, значит, оно готово к действию раньше, чем будет действительно использовано. Сила, создаваемая сократимым кольцом при цитокинезе, достаточно для того, чтобы согнуть тонкую стеклянную иглу, введенную в клетку. Таким способом, можно измерить величину этой силы. Нет сомнения, что источником силы сокращения здесь, так же как и в мышцах, служит взаимное скольжение актиновых и миозиновых филаментов. Если, например, к митотическим клеткам, обработанным детергентами, добавить инактивированные субфрагменты миозина, блокирующие миозин-связывающие участки актина, то разделение цитоплазмы прекратится. Точно так же введение антител к миозину в яйца морского ежа вызывает сглаживание борозды деления, но на деление ядра не влияет.
Во время сокращения сократимое кольцо сохраняет постоянную толщину. Это позволяет предполагать, что оно постепенно уменьшается в объеме за счет потери части филаментов. После завершения цитокинеза сократимое кольцо полностью распадается, а плазматическая мембрана в области борозды деления стягивается, окружая остаточное тельце.
В большинстве делящихся клеток борозда деления проходит более или менее симметрично, так как дочерние клетки получаются одинаковыми по размеру. Правда, в некоторых случаях сократимое кольцо, под действием которого происходит разделение цитоплазмы, образуется в таком месте, что получаются дочерние клетки разной величины. Такое асимметричное деление играет важную роль в оогенезе и часто наблюдается на ранних этапах развития некоторых эмбрионов, когда неоднородная цитоплазма яйца должна быть точно распределена между разными группами клеток, из которых впоследствии образуются различные части эмбриона. Механизмы, контролирующие в таких случаях ориентацию и расположение сократимого кольца остаются неизвестными.
Патологии митоза.
Роль патологий митоза.
Нарушение нормального течения митоза и неправильное распределение хромосом между дочерними клетками приводит к возникновению клеток с несбалансированными кариотипами. Патологический митоз – один из способов возникновения мутаций и развития анеуплоидии. С патологией связано развитие ряда заболеваний. Патологические митозы часто возникают при канцерогенезе и вирусной инфекции.
Патология митоза может носить функциональный и органический характер. Функциональные нарушения деления клеток могут выражаться, например, в гиперактивности вступающих в митоз клеток. Органические нарушения митоза возникают при повреждении структур (хромосомы, митотический аппарат, клеточная поверхность), участвующих в делении, и связанных с этими структурами процессов (редупликация ДНК, поляризация делящейся клетки, движение хромосом, цитотомия).
Классификация патологий митоза (И.А. Алов, 1972).
v Патологии митоза, связанные с повреждением хромосом.
1. Задержка митоза в профазе. Часто наблюдается при нарушении процессов редупликации хромосом.
2. Нарушения спирализации и деспирализации хромосом.
3. Раннее разделение хроматид.
4. Фрагментация хромосом и образование ацентрических фрагментов часто возникает в опухолях, а также при воздействии на нормальные клетки ионизирующего излучения и различных мутагенов. Большинство образовавшихся фрагментов лишено кинетохора, участвующего в движении хромосом. Поэтому ацентрические фрагменты остаются неподвижными и отстают при движении хромосом. Массивная фрагментация хромосом называется пульверизацией: при этом фрагменты беспорядочно рассеяны по цитоплазме и не участвуют в общем движении хромосом.
Судьба фрагментов различна: они могут попасть в одно из дочерних ядер, резорбироваться или образовать дополнительно микроядро.
6. Отставание хромосом в метакинезе и при расхождении к полюсам возникает при повреждении хромосомы в области кинетохора.
7. Образование микроядер. Судьба отставших хромосом различна. Они либо разрушаются, либо элиминируются из клетки, либо случайно попадают в одно из ядер, либо формируют дополнительное микроядро. Если микроядро формируется ядрышкообразующей хромосомой, то в нем возникает ядрышко. Дальнейшая судьба микроядер неодинакова. Одни из них подвергаются пикнозу, разрушаются и выводятся из клетки, другие, как и вся клетка, проделывают полный клеточный цикл.
8. Нерасхождение хромосом. При этом явлении сестринские хроматиды не разъединяются и вместе отходят к одному из полюсов. Это приводит к увеличению хромосомного набора одного из дочерних ядер за счет уменьшения другого на одну хромосому.
9. Набухание и слипание хромосом. Набухая, хромосомы утрачивают свои правильные очертания и, склеиваясь поверхностями, образуют неправильные комковатые массы. Клетки часто в таком состоянии довольно часто подвергаются гибели.
v Патологии митоза, связанные с повреждением митотического аппарата.
1. Задержка митоза в анафазе.
3. Рассеивание хромосом в метафазе. Связано с повреждением или с полной дезорганизацией митотического аппарата. Типичная экваториальная пластинка не образуется, а хромосомы беспорядочно рассеяны в цитоплазме. Рассеивание хромосом в метафазе отличается от к-митоза отсутствием других особенностей последнего и, прежде всего, типичных изменений хромосом (гиперспирализация, неразделение кинетохоров).
4. Многополюсный митоз. Связан с аномалией репродукции центриолей. Для многополюсного митоза характерно образование нескольких полюсов и веретен деления. Эта патология митоза приводит к неравномерному распределению хромосом между несколькими дочерними клетками, к образованию анеуплоидных клеток с несбалансированными наборами хромосом.
5. Моноцентрический митоз. Эта форма патологии также связана с нарушением разделения центриолей. При этой форме образуется только один полюс с веерообразным веретеном, соответствующем половине обычного двухполюсного веретена. При таком митозе происходит формирование хромосом и их разделение на сестринские хроматиды. Но последние не отходят друг от друга и образуют единую группу, из которой формируется одно полиплоидное ядро.
6. Ассиметричный митоз характеризуется неравномерным развитием противоположных митотических центров и связанных с ними конусов митотического веретена: одна половина митотического аппарата развита сильнее, чем другая.
7. Трехгрупповая метафаза и метафаза с полярными хромосомами. Трехгруппповая метафаза отличается тем, что метафазная клетка содержит, кроме обычной экваториальной пластинки, две дополнительные группы или одиночные хромосомы, расположенные у полюсов.
8. Полая метафаза. Имеет вид широкого кольца хромосом, которые, собираясь в метафазную пластинку, располагаются по периферии клеток.
v Патологии митоза, связанные с нарушением цитотомии.
1. Запаздывание или отсутствие цитотомии.
2. Преждевременная цитотомия.
Препараты и фотографии.
Центросома и центриоль.
Ø Полюс веретена деления. Продольный срез центриоли, поперечный срез центриоли. ЭМФ.
В митотических клетках центриоли в виде диплосом находятся в полюсах веретена деления. Такое расположение центриолей характерно для высших многоклеточных животных. На фотографии видна активность функционарования центриолей в качестве ЦОМТ.
Ø Редупликация центриолей. ЭМФ,
В клетках центриоли часто располагаются вблизи ядра или на некотором расстоянии от него, в полярных клетках они занимают апикальное расположение. На фотографии видны 2 центриоли и 2 процентриоли.
Аппарат движения.
Ø Ресничный эпителий. ЭМФ.
Ø Продольный срез ресничек. Рост аксонемы реснички от базального тела. ЭМФ.
Ø Поперечный срез ресничек. ЭМФ.
Ø Схема строения реснички.
Митоз.
Ø Дробление яйца аскариды. Препарат.
Каждая яйцеклетка окружена толстой гомогенной оболочкой, имеющей более темноокрашенный контур. На препарате необходимо найти деление на стадии яйцеклетки или двух бластомеров. В цитоплазме лежат 2 ли четыре аскариды (в зависимости от вида аскариды). У аскариды очень четко видно веретено деления и центриоли.
Препарат окрашен железным гематоксилином.
Ø Анафаза растительной клетки. Световая фотография.
Окрашивание коллоидным золотом.
Ø Цитокинез растительной клетки. Световая фотография.
На фотографии представлена растительная клетка на стадии поздней телофазы. Можно увидеть реконструкцию ядер. В центре клетки сформирован фрагмопласт.
Окрашивание коллоидным золотом.
Ø Метафаза животной клетки. Световая фатография.
Ø Стадии митоза в клетках пневмоцитов тритона: профаза, прометафаза, метафаза, анафаза, телофаза. Иммунофлуорисцентное окрашивание.
Иммунофлуорисцентное окрашивание: микротрубочки – зеленые, промежуточные филаменты – красные, хромосомы – синие.
Ø Кинетохор. ЭМФ.
Ø Кинетохор. Световая фотография.
Ø Микротрубочки, отходящие от кинетохора.ЭМФ.
На данной ЭМФ представлены кинетохорные микротрубочки и участок хромосомы, но не видно кинетохора.
Ø Цитотомия животной клетки. Образование перетяжки. ЭМФ.
В области перетяжки видно остаточное тельце, образованное микротрубочками.
© 2014-2022 — Студопедия.Нет — Информационный студенческий ресурс. Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав (0.014)

Злокачественная трансформация клеток опосредована генетическими нарушениями. Опухолевые клетки приобретают особый, отличный от нормальных клеток фенотип, на выявлении которого основана морфологическая верификация онкологического диагноза.
1. Клетка:
- Увеличение размеров атипичных клеток, по сравнению с нормальными клетками ткани, вплоть до гигантских размеров; но это необязательный признак;
- Изменение формы клеток, вплоть до самых причудливых. В опухолевой массе может отмечаться как выраженный клеточный полиморфизм (различие размеров и формы), так и, напротив, упрощение строения — мономорфность (наблюдается в низкодифференцированных мезенхимальных опухолях);
- Нарушение ядерно-цитоплазматического соотношения (за счет увеличения ядра);
- Наличие многоядерных клеток с отчетливым ядерным полиморфизмом;
- Атипичная локализация ядра;
- Диссоциация в созревании ядра и цитоплазмы (молодое ядро в зрелой клетке, зрелое ядро в незрелой цитоплазме) [1].
2. Ядро:
- Увеличение размера;
- Изменение формы, ядерный полиморфизм;
- Контур ядра неправильный, неравномерно извилистый, иногда с глубокими вырезками, грубо очерчен;
- Нарушение строения ядерной мембраны: неравномерно утолщена, отмечаются разрывы [1, 2].
- Патология митотического режима. Митотический режим — это комплекс параметров, характеризующих различные аспекты митоза. Например, митотический индекс — процент делящихся клеток от всей популяции — показатель, который отражает митотическую активность; структура митотически активных клеток — процентное соотношение делящихся клеток, находящихся на разных стадиях митоза; относительное количество всех патологических митозов; процентное содержание отдельных видов патологических митозов.
Спектр патологии митоза крайне широк. Но отмечаются некоторые характерные особенности. В частности, под ударом при злокачественной трансформации оказывается метафаза. Отмечается проявление колхицинового митоза, или К-митоза (форма патологического митоза с торможением процесса в результате инактивации веретена деления под действием статокинетических ядов, к которым относится в т. ч. и колхицин), а именно — К-метафаза. Для нее характерны слипание или рассеивание гиперспирализованных хромосом, отставание хромосом и их фрагментов в метафазе. Встречаются трехгрупповые метафазы, при которых, помимо основной экваториальной пластинки, присутствуют еще две группы в области полюсов деления клетки. Возможны метафазы с рассеиванием и массовой фрагментацией хромосом, ассиметричные, многополюсные и моноцентрические митозы [3].
3. Ядрышки:
- Визуализируются при исследовании;
- Увеличены в размерах;
- Форма неправильная, нередко полиморфизм;
- Увеличение количества ядрышек.
4. Хроматин:
- Различные паттерны: нарушение распределения, соотношения эу- и гетерохроматина;
- Крупнозернистый, грубый (часто отмечается в различных опухолях);
- Тонкий, мелкодисперсный, нежный, бластный (например, при остром миелобластном лейкозе);
Выраженность цито- и гистологических нарушений увеличивается с ростом степени злокачественности.
Патологическая анатомия немыслима без визуализации информации, поэтому представим процесс озлокачествления клеток на конкретном примере. В доставленном материале — фрагмент ткани мочевого пузыря. В нормальных условиях уротелиальные клетки относительно мономорфны, содержат достаточный объем цитоплазмы и овальные ядра, имеют округлые гладкие контуры, нежный хроматин, мелкие или вовсе невидимые ядрышки (рис. 1) [5].
.
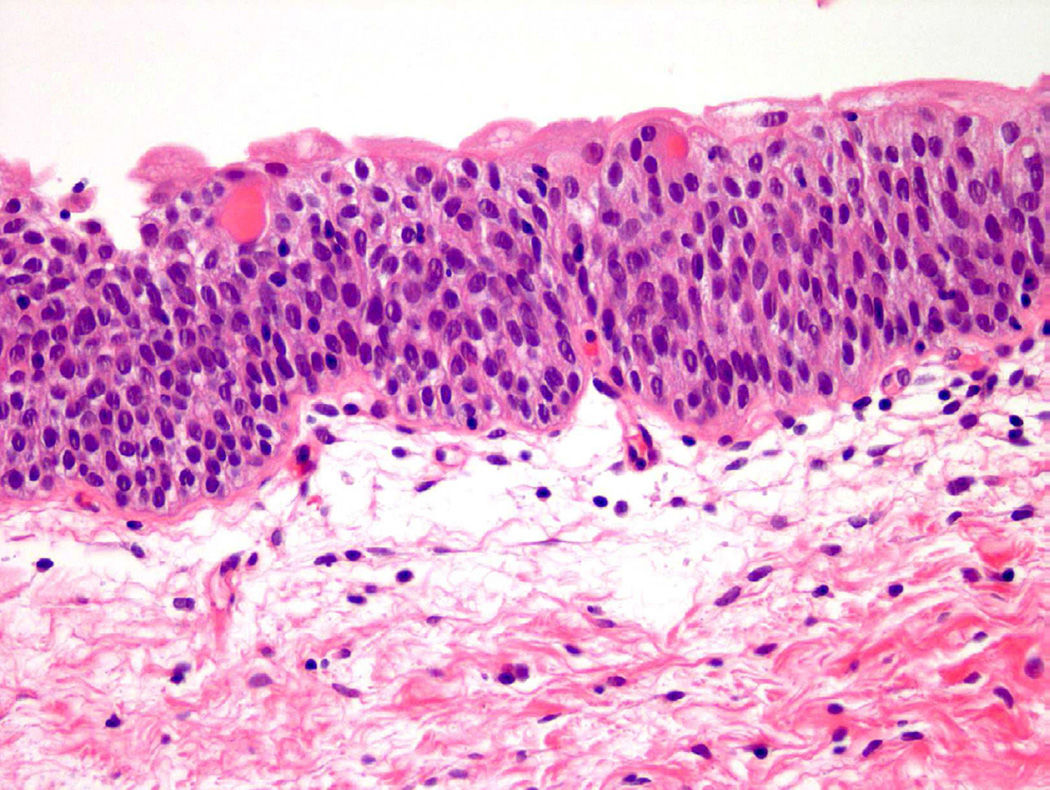
Рисунок 1 | Микроскопическая картина нормального уротелия [5]
Несмотря на большое разнообразие морфологии нормальных уротелиальных клеток, определены специфические особенности, указывающие на атипию. Информативны не поверхностные и не дегенеративные уротелиальные клетки. Основной маркер — увеличение ядерно-цитоплазматического соотношения (более 0,5) [5]. Дополнительные критерии: гиперхромия, неравномерный, грубый хроматин, неравномерный контур ядра. Если ядерно-цитоплазматическое соотношение более 0,7 и присутствуют два дополнительных признака, необходимо заподозрить уротелиальную карциному высокой степени злокачественности (рис. 2) [6].
.

Рисунок 2 | Папиллярная уротелиальная карцинома High Grade
Выраженная тканевая и клеточная атипия. Внутри папиллярных структур опухолевые клетки расположены хаотично, с утратой полярности. Значительный полиморфизм ядер, гиперхромность, отчетливые ядрышки. Митотическая активность увеличивается, фигуры митоза отмечаются во всех слоях уротелия. Единичные очаги некроза [8].
Цитологические критерии уротелиальной карциномы высокой степени злокачественности: высокое ядерно-цитоплазматическое соотношение, неравномерный контур мембран ядер, эксцентрическая локализация ядра, крупные, полиморфные ядра, выраженные ядрышки (рис. 3.1, 3.2) [7], могут обнаруживаться признаки плоскоклеточной и железистой дифференцировки (рис. 4) [8].
.
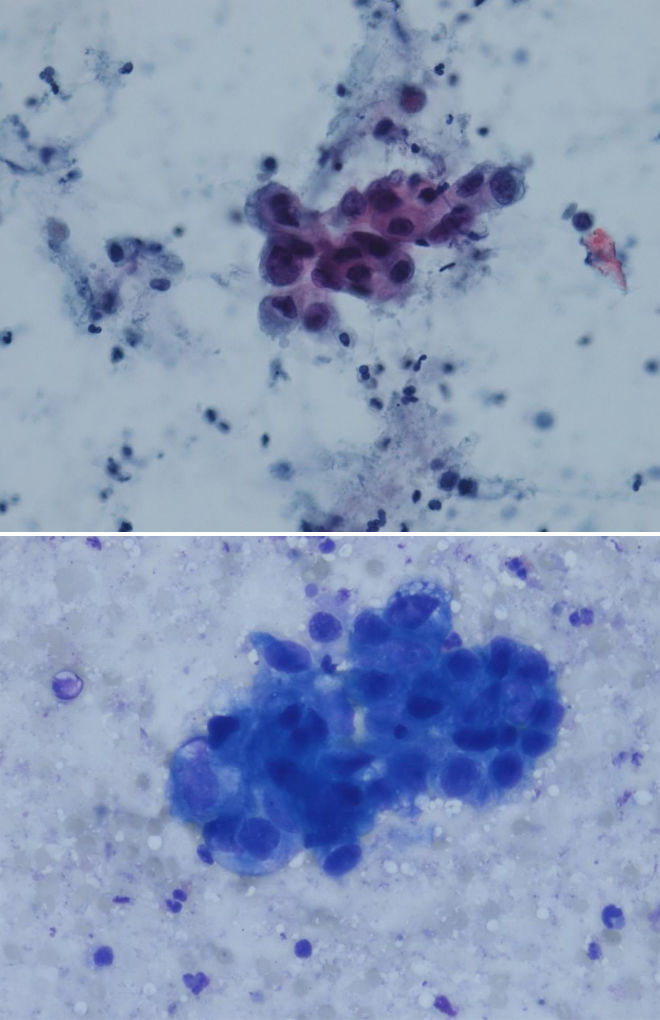
Рисунки 3.1 и 3.2 | Папиллярная уротелиальная карцинома High Grade
Крупные ядра с высоким ядерно-цитоплазматическим соотношением, грубый хроматин и нечеткие, размытые контуры ядра (окраска Papanicolaou и DiffQuik) [9].
.
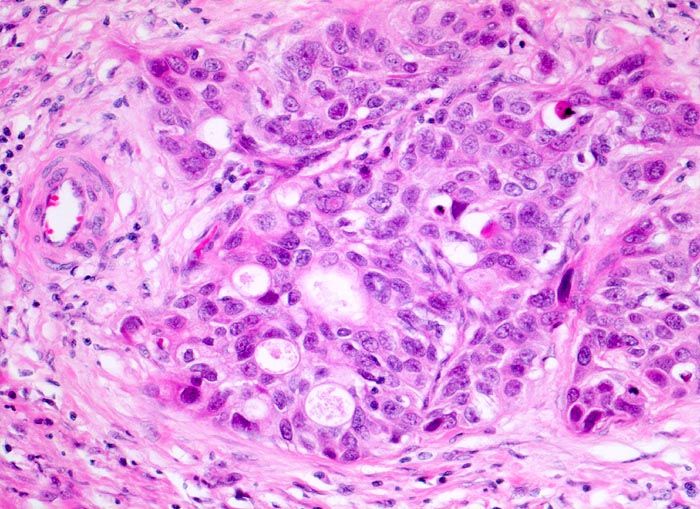
Рисунок 4 | Папиллярная уротелиальная карцинома High Grade с формированием железистых структур [10]
Читайте также: