
Регуляция работы генов у эукариот кратко
Обновлено: 28.06.2024
Система регуляции экспрессии генов у эукариот связана с особенностями функционирования эукариотического генома. Хотя и у прокариот, и у эукариот функционируют системы регуляторных белков, наличие ядра и нуклеосомная организация хроматина эукариот дают намного больше возможностей для регуляции.
Ядро явилось важнейшим эволюционным приобретением эукариот. Благодаря ядерной мембране, разделяются зоны транскрипции и трансляции, что позволяет осуществлять сложную и многообразную регуляцию экспрессии генов. Такая регуляция происходит на всех этапах.
Регуляция на уровне транскрипции. Основным уровнем регуляции экспрессии у эукариот является регуляция на уровне транскрипции. Варианты такой регуляции весьма разнообразны.
Наиболее универсальным методом регуляции транскрипции служит конденсация-деконденсация хроматина. Хорошо известно, что при активации генетического материала он деконденсируется. С другой стороны, далеко не весь эухроматин транскрибируется. Поэтому имеются и другие пути контроля транскрипции.
Для эукариот характерна дифференциальная экспрессия генов в разных клетках организма, поэтому многоклеточные организмы имеют разнообразные дифференцированные клетки. Хотя во всех клетках содержатся одинаковые гены, экспрессируются они не одинаково.
Основное направление регуляции клеточной дифференцировки осуществляется при помощи тканеспецифических транскрипционных факторов. Они представляют собой белки регуляторных генов, контролирующие дифференциальную активность других генов.
Иным направлением является использование альтернативных промоторов. В разных типах клеток с одного гена могут быть образованы разные белки. Так, ген белка дистрофина содержит 7 промоторов, которые считывают информацию, начиная с разных экзонов.
Большое значение в регуляции экспрессии генов у эукариот придается фактору метилирования ДНК внутри регуляторных областей. Метилированию подвергается цитозин в составе ЦГ-динуклеотида, что обычно приводит к инактивации гена. Деметилирование ДНК восстанавливает активность. Этот важный процесс регулируют ферменты метилтрансферазы.
С данным явлением схожа инактивация Х-хромосомы в соматических клетках женского организма млекопитающих. Правда, одна из двух Х-хромосом в клетках плода инактивируется случайным образом, поэтому в разных клетках женского организма экспрессируются как отцовские, так и материнские Х-хромосомы.
Для эукариот не характерна регуляция по типу оперона. Большинство м-РНК эукариот – моноцистронные. Более того, гены, контролирующие один метаболический путь, у эукариот часто разбросаны по всему геному. Но в этом правиле есть исключение.
Гены р-РНК и митохондрий транскрибируются на единую транскрипционную единицу, которая разрезается после процессинга. Считается, что такой способ позволяет унифицировать регуляцию схожих генов.
Регуляция на уровне процессинга. У эукариот транскрипция гена еще не означает его проявления в фенотипе. Молекулы РНК, синтезированные в ходе транскрипции, у эукариот подвергаются существенным модификациям. Совокупность таких преобразований и составляет сущность процессинга.
На уровне процессинга в первую очередь необходимо отметить механизм альтернативного сплайсинга, позволяющий изменять порядок сшивки экзонов. Таким образом, на основе одной и той же нуклеотидной последовательности одного гена формируются разные белки, состоящие из разных сочетаний одних и тех же аминокислотных блоков.
Феномен альтернативного сплайсинга изменил наши представления о природе гена. Его распространенность оказалась значительно выше, а роль в эволюции – важнее, чем предполагалось первоначально. Альтернативный сплайсинг характерен для более чем трети генов человека.
Так, на гене белка тропонина, содержащего 18 экзонов, за счет альтернативного сплайсинга может образовываться 64 различных белковых продукта. В среднем один ген человека кодирует около трех белков.
Другим интересным способом регуляции на уровне процессинга является тканеспецифическое редактирование РНК. Оно обнаружено у микроорганизмов, грибов, млекопитающих и проявляется заменой отдельных нуклеотидов в молекуле РНК при помощи специального ферментного комплекса.
Если в случае замены вместо смыслового кодона образуется стоп-кодон, то в новой полипептидной цепи будут отсутствовать все аминокислоты, идущие после него. Получается белок с совершенно новыми свойствами.
В ооцитах некоторых животных происходит накопление и-РНК, у которых не закончен процессинг. Такие РНК не транслируются. Окончание процессинга и последующая экспрессия наступают только после оплодотворения.
Примерно половина гя-РНК полностью распадается в ядре, не выходя за его пределы. Возможно распаду подвержены такие транскрипты, которые не способны превратиться в зрелую и-РНК.
Регуляция на уровне трансляции. Механизмы регуляции экспрессии на уровне трансляции изучены недостаточно полно. Избирательная трансляция м-РНК осуществляется отбором определенных м-РНК путем блокировки доступа к рибосомам.
В случае избирательной стабилизации определенных типов м-РНК в цитоплазме, они не подвергаются распаду после трансляции. Наглядным примером дифференциальной стабильности м-РНК может служить синтез белков глобинов на стабильных м-РНК безъядерных ретикулоцитов млекопитающих.
Мутации, блокирующие полиаденилирование, наоборот, приводят к образованию очень нестабильных транскриптов. Обнаружены и другие механизмы, регулирующие стабильность м-РНК.
Регуляция на уровне посттрансляционной модификации белка. Посттрансляционная модификация полипептида и превращение его в функционально активную молекулу белка завершает процесс реализации генетической информации. Она представляет собой различные модификации определенных аминокислот (фосфорилирование, ацетилирование), удаление некоторых из них, и на этой основе формирование вторичной, третичной, четвертичной структуры белка.
На посттрансляционном уровне также возможна регуляция экспрессии. Широко распространен механизм регуляции активности ферментов, основанный на присоединении молекул-эффекторов, в роли которых часто выступают конечные продукты биосинтеза.
В последние годы в геномах разных животных были обнаружены многочисленные РНК, выполняющие регуляторные функции. Они получили название риборегуляторов. Особое значение имеют короткие двунитиевые микро-РНК. Их способность ослаблять работу отдельных генов получила название РНК-интерференции.
Проблема регуляции экспрессии генов приводит к вопросу о координации этой регуляции: каким образом в каждой клетке происходит активация именно необходимой комбинации генов, определяющих ее фенотип? Для млекопитающих (в том числе и для человека) установлено наличие большого числа факторов регуляции надклеточного уровня, включая факторы нервной и эндокринной систем.
Работа генов в любом организме – прокариотическом, эукариотическом, одноклеточном или многоклеточном контролируется и координируется.
11.1. Регуляция работы генов у прокариот.
Классическим примером регуляции экспрессии (или реализации) служит: функционирование 1) лактозного и 2) триптафанового оперона кишечной палочки.
Лактозный оперон. Многими учеными замечено, что некоторые ферменты у дрожжей и бактерий образуются в клетках только при выращивании их на определенных питательных средах. Например, при выращивании кишечной палочки на питательной среде, не содержащей лактозы, ее клетки содержат незначительное число (меньше пяти) молекул фермента лактазы, разлагающего лактозу на глюкозу и галактозу. При добавлении в питательную среду лактозы бактериальные клетки в течение 2-3 мин синтезируют большое количество лактазы (свыше 5 тыс. молекул). При удалении из среды лактозы синтез лактазы быстро прекращается. Вещества, индуцирующие синтез ферментов, которые их разлагают, называются индукторами (в данном примере индуктором является лактоза).
Триптафановый оперон. Подобные механизмы используются клеткой для выключения синтеза нужных ей соединений при их наличии в питательной среде. Например, аминокислота триптофан синтезируется при участии фермента триптофан-синтетазы. Однако, если в среде, на которой выращиваются бактерии, присутствует триптофан, синтез фермента немедленно прекращается. Это явление получило название репрессии, а вызывающий его фактор (в нашем примере — триптофан) — корепрессором.
а)
б)
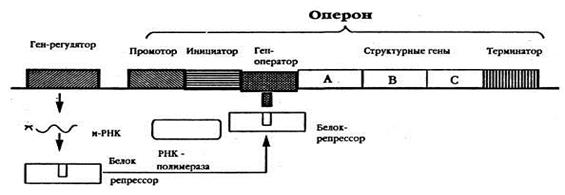
Оперон – это группа генов (т.е. участок ДНК), работа которых контролируется одним геном-регулятором. Единица считывания информации у прокариот.
Ген-регулятор – это ген, находящийся обычно на некотором расстоянии от оперона (т.е. он не входит в организацию оперона), постоянно активен и на основе его информации синтезируется особый белок-репрессор. Репрессор обладает очень сильным сродством к гену- оператору и может легко присоединяться к нему.
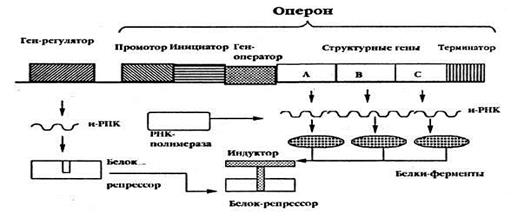
Индукция. Присоединение молекулы индуктора к активному участку молекулы репрессора изменяет его третичную структуру так, что он не может связаться с геном-оператором и блокировать его. Оператор оказывается в активном состоянии и включает структурные гены, а именно, отсутствие блокады гена оператора позволяет РНК-полимеразе связаться с геном-промотором. Закрепившись, РНК-полимераза может начать транскрипцию структурных генов оперона. В качестве индукторов могут выступать продукты распада белка, гормоны и др. метаболиты.
Инактивация. Если индуктор в клетке отсутствует, то репрессор обладает способностью связываться с геном-оператором. При этом происходит инактивация оператора и тем самым предотвращается включение структурных генов, т.е. РНК-полимераза не связывается с геном-промотором и транскрипция отсутствует
Оперон включает в себя следующие гены:
1. Ген-промотор с инициатором - это место первичного прикрепления РНК-полимеразы (фермента, синтезирующего РНК на ДНК)— фермента, катализирующего реакции ДНК-зависимого синтеза и-РНК. Является местом начала транскрипции. Кроме того, промотор определяет, какая из двух цепей ДНК будет служит матрицей для синтеза иРНК. Промотор находится со стороны 5'-конца кодирующей цепи гена.
2. Ген-оператор - точка начала считывания генетической информации, он управляет функционированием структурных генов оперона, т.е. включает и выключает их (т.е. структурные гены), следовательно, они активны непостоянно. Если этот ген свободен, то это значит, что работа структурных генов разрешена, если он связывается с репрессором, то работа этих генов прекращается.

3. Структурные гены - это гены, несущие генетическую информацию о последовательности аминокислот в молекулах белков (ферментов).
4. Терминатор– это участок (локус) в конце оперона, сигнализирующий о прекращении транскрипции, т.е. место окончания синтеза иРНК.
11.2. Регуляция работы генов у эукариот. Схема регуляции транскрипции у эукариот разработана Г. П. Георгиевым (1972).
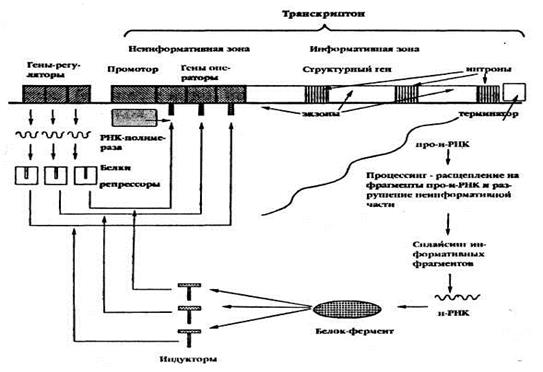
Рис. ___. Схема регуляции транскрипции у эукариот
Единица транскрипции (т.е. единица считывания информации) у эукариот называется транскриптоном. Он состоит из неинформативной (акцепторной) и информативной (структурной) зон.
Неинформативная зона начинается промотором с инициатором. Далее следуют группа генов-операторов, за которыми расположена информативная зона.
Информативная зона образована структурным геном, разделенным на экзоны (информативные участки) и интроны. (неинформативные участки).
Заканчивается транскриптон терминатором.
Работу транскриптона регулирует несколько генов-регуляторов, дающих информацию для синтеза нескольких белков-репрессоров. Индукторами в клетках эукариот являются сложные молекулы (например, гормоны), для расщепления которых требуется несколько ферментов (многоступенчатые реакции). Когда индукторы освобождают гены-операторы от белков-репрессоров, РНК-полимераза разрывает водородные связи между двумя цепочками ДНК транскриптона и по правилу комплементарности на нем сначала синтезируется большая молекула проинформационной РНК (про-и-РНК), списывающая информацию как с информативной, так и с неинформативной зон. В дальнейшем в ядре клетки происходит процессинг(англ. processing –обработка, переработка) — ферментативное разрушение неинформативной части РНК и расщепление ферментами рестриктазами информативной части на фрагменты, соответствующие экзонам. Молекула и-РНК (моноцистронпая), соответствующая экзонам структурного гена, формируется посредством сплайсинга(сплавления или соединения) отдельных информативных фрагментов ферментами лигазами. Далее и-РНК выходит из ядра, идет в рибосомы, где и происходитсинтез белка-фермента, необходимого для расщепления индукторов. Включение и выключение транскриптона происходит принципиально так же, как и оперона.
Работа генов в любом организме – прокариотическом, эукариотическом, одноклеточном или многоклеточном контролируется и координируется.
11.1. Регуляция работы генов у прокариот.
Классическим примером регуляции экспрессии (или реализации) служит: функционирование 1) лактозного и 2) триптафанового оперона кишечной палочки.
Лактозный оперон. Многими учеными замечено, что некоторые ферменты у дрожжей и бактерий образуются в клетках только при выращивании их на определенных питательных средах. Например, при выращивании кишечной палочки на питательной среде, не содержащей лактозы, ее клетки содержат незначительное число (меньше пяти) молекул фермента лактазы, разлагающего лактозу на глюкозу и галактозу. При добавлении в питательную среду лактозы бактериальные клетки в течение 2-3 мин синтезируют большое количество лактазы (свыше 5 тыс. молекул). При удалении из среды лактозы синтез лактазы быстро прекращается. Вещества, индуцирующие синтез ферментов, которые их разлагают, называются индукторами (в данном примере индуктором является лактоза).
Триптафановый оперон. Подобные механизмы используются клеткой для выключения синтеза нужных ей соединений при их наличии в питательной среде. Например, аминокислота триптофан синтезируется при участии фермента триптофан-синтетазы. Однако, если в среде, на которой выращиваются бактерии, присутствует триптофан, синтез фермента немедленно прекращается. Это явление получило название репрессии, а вызывающий его фактор (в нашем примере — триптофан) — корепрессором.
а)
б)
Оперон – это группа генов (т.е. участок ДНК), работа которых контролируется одним геном-регулятором. Единица считывания информации у прокариот.
Ген-регулятор – это ген, находящийся обычно на некотором расстоянии от оперона (т.е. он не входит в организацию оперона), постоянно активен и на основе его информации синтезируется особый белок-репрессор. Репрессор обладает очень сильным сродством к гену- оператору и может легко присоединяться к нему.
Индукция. Присоединение молекулы индуктора к активному участку молекулы репрессора изменяет его третичную структуру так, что он не может связаться с геном-оператором и блокировать его. Оператор оказывается в активном состоянии и включает структурные гены, а именно, отсутствие блокады гена оператора позволяет РНК-полимеразе связаться с геном-промотором. Закрепившись, РНК-полимераза может начать транскрипцию структурных генов оперона. В качестве индукторов могут выступать продукты распада белка, гормоны и др. метаболиты.
Инактивация. Если индуктор в клетке отсутствует, то репрессор обладает способностью связываться с геном-оператором. При этом происходит инактивация оператора и тем самым предотвращается включение структурных генов, т.е. РНК-полимераза не связывается с геном-промотором и транскрипция отсутствует
Оперон включает в себя следующие гены:
1. Ген-промотор с инициатором - это место первичного прикрепления РНК-полимеразы (фермента, синтезирующего РНК на ДНК)— фермента, катализирующего реакции ДНК-зависимого синтеза и-РНК. Является местом начала транскрипции. Кроме того, промотор определяет, какая из двух цепей ДНК будет служит матрицей для синтеза иРНК. Промотор находится со стороны 5'-конца кодирующей цепи гена.
2. Ген-оператор - точка начала считывания генетической информации, он управляет функционированием структурных генов оперона, т.е. включает и выключает их (т.е. структурные гены), следовательно, они активны непостоянно. Если этот ген свободен, то это значит, что работа структурных генов разрешена, если он связывается с репрессором, то работа этих генов прекращается.
3. Структурные гены - это гены, несущие генетическую информацию о последовательности аминокислот в молекулах белков (ферментов).
4. Терминатор– это участок (локус) в конце оперона, сигнализирующий о прекращении транскрипции, т.е. место окончания синтеза иРНК.
11.2. Регуляция работы генов у эукариот. Схема регуляции транскрипции у эукариот разработана Г. П. Георгиевым (1972).
Рис. ___. Схема регуляции транскрипции у эукариот
Единица транскрипции (т.е. единица считывания информации) у эукариот называется транскриптоном. Он состоит из неинформативной (акцепторной) и информативной (структурной) зон.
Неинформативная зона начинается промотором с инициатором. Далее следуют группа генов-операторов, за которыми расположена информативная зона.
Информативная зона образована структурным геном, разделенным на экзоны (информативные участки) и интроны. (неинформативные участки).
Заканчивается транскриптон терминатором.
Работу транскриптона регулирует несколько генов-регуляторов, дающих информацию для синтеза нескольких белков-репрессоров. Индукторами в клетках эукариот являются сложные молекулы (например, гормоны), для расщепления которых требуется несколько ферментов (многоступенчатые реакции). Когда индукторы освобождают гены-операторы от белков-репрессоров, РНК-полимераза разрывает водородные связи между двумя цепочками ДНК транскриптона и по правилу комплементарности на нем сначала синтезируется большая молекула проинформационной РНК (про-и-РНК), списывающая информацию как с информативной, так и с неинформативной зон. В дальнейшем в ядре клетки происходит процессинг(англ. processing –обработка, переработка) — ферментативное разрушение неинформативной части РНК и расщепление ферментами рестриктазами информативной части на фрагменты, соответствующие экзонам. Молекула и-РНК (моноцистронпая), соответствующая экзонам структурного гена, формируется посредством сплайсинга(сплавления или соединения) отдельных информативных фрагментов ферментами лигазами. Далее и-РНК выходит из ядра, идет в рибосомы, где и происходитсинтез белка-фермента, необходимого для расщепления индукторов. Включение и выключение транскриптона происходит принципиально так же, как и оперона.
Урок посвящен знакомству с механизмами транскрипции и трансляции в клетках эукариот и прокариот. Вы узнаете о современных направлениях молекулярной биологии – генной и клеточной инженерии. А так же познакомитесь с вирусами – неклеточной форме жизни.
4. Глоссарий по теме (перечень терминов и понятий, введенных на данном уроке);
оператор - участок ДНК между промотором и структурными генами в опероне располагается;
репрессор - связан особый белок связанный с оператором при этом РНК-полимераза не может начать синтез иРНК.
Оперон – это группа генов (т.е. участок ДНК), работа которых контролируется одним геном-регулятором. Единица считывания информации у прокариот.
Ген-регулятор – это ген, находящийся обычно на некотором расстоянии от оперона (т.е. он не входит в организацию оперона), постоянно активен и на основе его информации синтезируется особый белок-репрессор
Бактериофаг - вирус, инфицирующий бактерии.
Вирион - вирусная частица.
Капсид - белковая оболочка вирусной частицы.
Клонирование - совокупность процедур, использующихся для получения клонов.
Ретровирусы - группа РНК-содержащих вирусов, содержащих обратную транскриптазу; синтезированная на РНК-матрице двухцепочечная ДНК может встраиваться в хромосому инфицированной этим вирусом клетки.
Генная инженерия – (Gene engineering) – совокупность приемов, методов и технологий, в том числе технологий получения рекомбинантных рибонуклеиновых и дезоксирибонуклеиновых кислот, по выделению единичных или нескольких генов из организма, осуществлению манипуляций с генами и введению их в другие организмы.
Клеточная инженерия – это один из основных разделов современной биотехнологии, основанный на выделении и культивировании тканей и клеток высших многоклеточных организмов
Биотехноло́гия — интеграция естественных и инженерных наук, позволяющая наиболее полно реализовать возможности живых организмов или их производные для создания и модификации продуктов или процессов различного назначения
5. Основная и дополнительная литература по теме урока (точные библиографические данные с указанием страниц);
1. Общая биология 10-11, дидактические материалы/ авт.-сост С.С. Красновидова, С. А. Павлов, А. Б. Павлов, - М. Просвещение, 2000г., стр.6-42
2. Общая биология 10-11 классы: подготовка к ЕГЭ. Контрольные и самостоятельные работы/ Г. И. Лернер. – М.: Эксмо, 2007.стр 35-45
3. Биология: общая биология. 10-11 классы: учебник/ А. А. Каменский, Е. А. Криксунов, В. В. Пасечник.- М.: Дрофа, 2018. Стр.55-68
6. Открытые электронные ресурсы по теме урока (при наличии);
7. теоретический материал для самостоятельного изучения;
Ген – это фрагмент молекулы ДНК, содержащий регуляторные элементы и структурную область, и соответствующий одной единице транскрипции, которая определяет возможность синтеза полипептидной цепи или молекулы РНК.
Ген прокариот называется опероном, в его состав входят два основных участка:
- регуляторный (неинформативный),
- структурный (информативный).
У прокариот на долю регуляторных элементов приходится около 10 %, структурных – 90 %.
Структурная область генов прокариот (единица транскрипции) может быть представлена одним кодирующим участком, который называется цистроном, либо несколькими кодирующими участками (полицистронная единица транскрипции). В структурной зоне закодирована информация о последовательности аминокислот в виде генетического кода. Со структурной области считывается мРНК. При наличии у прокариот полицистронной единицы транскрипции на одном структурном участке одновременно может синтезироваться несколько разновидностей мРНК.
К регуляторным элементам генов прокариот относятся участки, управляющие работой гена:
- промотор,
- оператор,
- терминатор.
Промотор определяет начало транскрипции (участок инициации). С промотором соединяется фермент РНК-полимераза, осуществляющий синтез мРНК. Другой элемент, управляющий процессом транскрипции, – оператор, который располагается поблизости от промотора или внутри него. Этот участок может быть свободным, тогда РНК-полимераза соединяется с промотором и начинается транскрипция. Если оператор связан с белком-репрессором, РНК-полимераза не может нормально соединиться с промотором, и транскрипция невозможна. Следующий регуляторный элемент – терминатор – находится за структурной областью и содержит сигнальный участок остановки транскрипции.

Механизм функционирования системы регуляции синтеза белка был открыт в 1962 году Жакобом и Моно при исследовании культивирования кишечной палочки в лактозной среде и назван lac-опероном.
Упрощенно этот механизм может быть описан следующим образом. На основе информации гена-регулятора синтезируется белок-репрессор; если он активный, он связывается с геном-оператором, перекрывая путь для РНК-полимеразы – процесс трансляции и последующего синтеза белка выключается (запрещается). Если появляется индуктор (например, лактоза в lac-опероне), он присоединяется с белку-репрессору, приводя его в неактивное состояние. Оператор становится активным и включает процесс считывания информации со структурных генов – разрешает трансляцию. Происходит считывание информации с ДНК, начинается синтез необходимого белка – фермента (например, β-галактозидазы в lac-опероне).
Это только один из возможных механизмов, который называется запрещающей индукцией. Существуют и другие механизмы регуляции синтеза белка: разрешающая индукция, разрешающая и запрещающая репрессия, в которых принимают участие апоиндукторы и корепрессоры.
Строение генов у эукариот намного сложнее. Генетическая система эукариот называется транскриптоном. Транскриптон также состоит из двух частей:
- регуляторной (неинформативной),
- структурной (информативной),
относительная пропорция которых противоположна генам прокариот: на долю регуляторного участка приходится 90 %, структурного – 10 %.
Регуляторный участок представляет собой ряд последовательно расположенных промоторов и операторов и несколько терминаторов. Структурный участок состоит из одной единицы транскрипции и имеет “прерывистое” строение: кодирующие участки (экзоны) чередуются с некодирующими (интронами). Одномоментно на структурном участке у эукариот может синтезироваться только одна молекула мРНК, однако благодаря наличию альтернативного сплайсинга в разнос время (в зависимости от потребности клетки) на одной и той же структурной части могут синтезироваться разные виды мРНК (от одной до нескольких десятков).
Вирус (от лат. virus — яд) — простейшая форма жизни, микроскопическая частица, представляющая собой молекулы нуклеиновых кислот (ДНК или РНК, некоторые, например, мимивирусы, имеют оба типа молекул), заключённые в белковую оболочку и способные инфицировать живые организмы. От других инфекционных агентов вирусы отличает капсид. Вирусы, за редким исключением, содержат только один тип нуклеиновой кислоты: либо ДНК, либо РНК.
Вирусы являются паразитами, так как вирусы не способны размножаться вне клетки. Вне клетки вирусные частицы ведут себя как химические вещества. В настоящее время известны вирусы, размножающиеся в клетках растений, животных, грибов и бактерий (последних обычно называют бактериофагами). Обнаружены также вирусы, поражающие другие вирусы (вирусы-сателлиты).
В наше время существуют три гипотезы происхождения вирусов. виды вирусов в биологии Гипотеза клеточного происхождения сообщает о том, что внеклеточные агенты появились из фрагментов РНК и ДКН, которые смогли высвободиться от организма большего размера. Регрессивная гипотеза показывает, что вирусы были мелкими клетками, ведущими паразитический образ жизни в более крупных видах, но со временем утратили гены, которые нужны для паразитического существования. Гипотеза коэволюции предполагает, что вирусы возникли в то же время, в которое появились живые клетки, то есть уже миллиарды лет назад. И появились в результате построения сложных комплексов нуклеиновых кислот и белков. Кратко о вирусах (по биологии этих организмов база знаний наша, к сожалению, далека от совершенства) вы можете прочитать в данной статье. Каждая из перечисленных выше теорий имеет свои минусы и недоказанные гипотезы. Вирусы как форма жизни. Существует два определения формы жизни вирусов. Согласно первому, внеклеточные агенты - это комплекс органических молекул. Второе определение сообщает о том, что вирусы являются особой формой жизни. Вирусы (биология подразумевает появление многих новых видов вирусов) характеризуются как организмы на границе живого. Они похожи на живые клетки тем, что имеют свой неповторимый набор генов и эволюционируют исходя из метода естественного отбора. Также они могут размножаться, создавая при этом собственные копии. Так как вирусы не имеют клеточного строения, ученые не рассматривают их как живую материю. молекулярная биология вирусов Для того чтобы синтезировать собственные молекулы, внеклеточным агентам нужна клетка-хозяин. Отсутствие собственного обмена веществ не позволяет им размножаться без посторонней помощи.
Цикл вируса состоит из нескольких этапов, которые являются взаимоперекрывающимися. На первом этапе вирус прикрепляется, то есть образовывает специфическую связь между своими белками и рецепторами клетки-хозяина. Далее нужно проникнуть в саму клетку и передать ей свой генетический материал. Некоторые виды переносят еще и белки. После этого происходит потеря капсида, и геномная нуклеиновая кислота высвобождается. После того как паразит попадает внутрь клетки, начинается сборка вирусных частиц и модификация белка. И в итоге вирус выходит из клетки. Даже если он продолжает активно развиваться, то может и не убивать клетку, а продолжать в ней жить. Заболевания человека Вирусы биология интерпретирует как низшее проявление жизни на планете Земля. Одним из самых простых вирусных заболеваний человека является простуда. Однако данные паразиты могут вызывать и очень серьезные заболевания, такие как СПИД или птичий грипп. вирусы и бактерии биология Каждый вирус имеет определенный механизм действия на своего хозяина. Этот процесс включает лизис клеток, который приводит к их смерти. У многоклеточных организмов при отмирании большого количества клеток начинает плохо функционировать весь организм. Во многих случаях вирусы могут и не наносить вреда человеческому здоровью. В медицине это называется латентностью. Примером такого вируса является герпес. Некоторые латентные виды способны приносить пользу. Порой их присутствие вызывает иммунный ответ против бактериальных патогенов.
8. примеры и разбор решения заданий тренировочного модуля (не менее 2 заданий).

Используя конспект урока, найдите и выделите цветом по вертикали и горизонтали в филворде основные понятия втранскрипции и трансляции у прокариот. (надписи на рисунке убрать)

Правильный вариант: репрессор, оперон, субстрат, оператор, промотор.
Вставьте пропущенные слова.
Регуляция генной активности у высших организмов сложнее, чем у _____. У эукариот эти процессы регулируют ________. Они образуются в специальных клетках ______ внутренней секреции. Они регулируют синтез _______ в специальных клетках – мишенях.
Правильный вариант:
Регуляция генной активности у высших организмов сложнее, чем у бактерий. У эукариот эти процессы регулируют гормоны. Они образуются в специальных клетках желёз внутренней секреции. Они регулируют синтез РНК в специальных клетках – мишенях.
Подсказка: Еще раз посмотрите как происходит регуляция транскрипции у эукариот..
Система регуляции экспрессии генов у эукариот связана с особенностями функционирования эукариотического генома. Хотя и у прокариот, и у эукариот функционируют системы регуляторных белков, наличие ядра и нуклеосомная организация хроматина эукариот дают намного больше возможностей для регуляции. Ядро явилось важнейшим эволюционным приобретением эукариот. Благодаря ядерной мембране, разделяются зоны транскрипции и трансляции, что позволяет осуществлять сложную и многообразную регуляцию экспрессии генов. Такая регуляция происходит на всех этапах.
Регуляция на уровне транскрипции. Основным уровнем регуляции экспрессии у эукариот является регуляция на уровне транскрипции. Варианты такой регуляции весьма разнообразны.
Наиболее универсальным методом регуляции транскрипции служит конденсация-деконденсация хроматина. Хорошо известно, что при активации генетического материала он деконденсируется. С другой стороны, далеко не весь эухроматин транскрибируется. Поэтому имеются и другие пути контроля транскрипции.
Для эукариот характерна дифференциальная экспрессия генов в разных клетках организма, поэтому многоклеточные организмы имеют разнообразные дифференцированные клетки. Хотя во всех клетках содержатся одинаковые гены, экспрессируются они не одинаково. Основное направление регуляции клеточной дифференцировки осуществляется при помощи тканеспецифических транскрипционных факторов. Они представляют собой белки регуляторных генов, контролирующие дифференциальную активность других генов.
Иным направлением является использование альтернативных промоторов. В разных типах клеток с одного гена могут быть образованы разные белки. Так, ген белка дистрофина содержит 7 промоторов, которые считывают информацию, начиная с разных экзонов.
Большое значение в регуляции экспрессии генов у эукариот придается фактору метилирования ДНК внутри регуляторных областей. Метилированию подвергается цитозин в составе ЦГ-динуклеотида, что обычно приводит к инактивации гена. Деметилирование ДНК восстанавливает активность. Этот важный процесс регулируют ферменты метилтрансферазы.
С данным явлением схожа инактивация Х-хромосомы в соматических клетках женского организма млекопитающих. Правда, одна из двух Х-хромосом в клетках плода инактивируется случайным образом, поэтому в разных клетках женского организма экспрессируются как отцовские, так и материнские Х-хромосомы.
Для эукариот не характерна регуляция по типу оперона. Большинство м-РНК эукариот – моноцистронные. Более того, гены, контролирующие один метаболический путь, у эукариот часто разбросаны по всему геному. Но в этом правиле есть исключение. Гены р-РНК и митохондрий транскрибируются на единую транскрипционную единицу, которая разрезается после процессинга. Считается, что такой способ позволяет унифицировать регуляцию схожих генов.
Регуляция на уровне процессинга. У эукариот транскрипция гена еще не означает его проявления в фенотипе. Молекулы РНК, синтезированные в ходе транскрипции, у эукариот подвергаются существенным модификациям. Совокупность таких преобразований и составляет сущность процессинга.
На уровне процессинга в первую очередь необходимо отметить механизм альтернативного сплайсинга, позволяющий изменять порядок сшивки экзонов. Таким образом, на основе одной и той же нуклеотидной последовательности одного гена формируются разные белки, состоящие из разных сочетаний одних и тех же аминокислотных блоков.
Феномен альтернативного сплайсинга изменил наши представления о природе гена. Его распространенность оказалась значительно выше, а роль в эволюции – важнее, чем предполагалось первоначально. Альтернативный сплайсинг характерен для более чем трети генов человека. Так, на гене белка тропонина, содержащего 18 экзонов, за счет альтернативного сплайсинга может образовываться 64 различных белковых продукта. В среднем один ген человека кодирует около трех белков (Тарантул В. З., 2003).
Другим интересным способом регуляции на уровне процессинга является тканеспецифическое редактирование РНК. Оно обнаружено у микроорганизмов, грибов, млекопитающих и проявляется заменой отдельных нуклеотидов в молекуле РНК при помощи специального ферментного комплекса. Если в случае замены вместо смыслового кодона образуется стоп-кодон, то в новой полипептидной цепи будут отсутствовать все аминокислоты, идущие после него. Получается белок с совершенно новыми свойствами.
В ооцитах некоторых животных происходит накопление и-РНК, у которых не закончен процессинг. Такие РНК не транслируются. Окончание процессинга и последующая экспрессия наступают только после оплодотворения.
Примерно половина гя-РНК полностью распадается в ядре, не выходя за его пределы. Возможно распаду подвержены такие транскрипты, которые не способны превратиться в зрелую и-РНК.
Регуляция на уровне трансляции. Механизмы регуляции экспрессии на уровне трансляции изучены недостаточно полно. Избирательная трансляция м-РНК осуществляется отбором определенных м-РНК путем блокировки доступа к рибосомам.
В случае избирательной стабилизации определенных типов м-РНК в цитоплазме, они не подвергаются распаду после трансляции. Наглядным примером дифференциальной стабильности м-РНК может служить синтез белков глобинов на стабильных м-РНК безъядерных ретикулоцитов млекопитающих. Мутации, блокирующие полиаденилирование, наоборот, приводят к образованию очень нестабильных транскриптов. Обнаружены и другие механизмы, регулирующие стабильность м-РНК.
Регуляция на уровне посттрансляционной модификации белка. Посттрансляционная модификация полипептида и превращение его в функционально активную молекулу белка завершает процесс реализации генетической информации. Она представляет собой различные модификации определенных аминокислот (фосфорилирование, ацетилирование), удаление некоторых из них, и на этой основе формирование вторичной, третичной, четвертичной структуры белка. На посттрансляционном уровне также возможна регуляция экспрессии. Широко распространен механизм регуляции активности ферментов, основанный на присоединении молекул-эффекторов, в роли которых часто выступают конечные продукты биосинтеза.
В последние годы в геномах разных животных были обнаружены многочисленные РНК, выполняющие регуляторные функции. Они получили название риборегуляторов. Особое значение имеют короткие двунитиевые микро-РНК. Их способность ослаблять работу отдельных генов получила название РНК-интерференции.
Проблема регуляции экспрессии генов приводит к вопросу о координации этой регуляции: каким образом в каждой клетке происходит активация именно необходимой комбинации генов, определяющих ее фенотип? Для млекопитающих (в том числе и для человека) установлено наличие большого числа факторов регуляции надклеточного уровня, включая факторы нервной и эндокринной систем.
Читайте также: