
Работа и сила мышц физиология кратко
Обновлено: 05.07.2024
Мышечное сокращение. Характеристика и механизм мышечного сокращения
Выделяют несколько последовательных этапов запуска и осуществления мышечного сокращения.
1. Потенциал действия распространяется вдоль двигательного нервного волокна до его окончаний на мышечных волокнах.
2. Каждое нервное окончание секретирует небольшое количество нейромедиатора ацетилхолина.
3. Ацетилхолин действует на ограниченную область мембраны мышечного волокна, открывая многочисленные управляемые ацетилхолином каналы, проходящие сквозь белковые молекулы, встроенные в мембрану.
4. Открытие управляемых ацетилхолином каналов позволяет большому количеству ионов натрия диффундировать внутрь мышечного волокна, что ведет к возникновению на мембране потенциала действия.
5. Потенциал действия проводится вдоль мембраны мышечного волокна так же, как и по мембране нервного волокна.
6. Потенциал действия деполяризует мышечную мембрану, и большая часть возникающего при этом электричества течет через центр мышечного волокна. Это ведет к выделению из саркоплазматического ретикулума большого количества ионов кальция, которые в нем хранятся.
7. Ионы кальция инициируют силы сцепления между актиновыми и миозиновыми нитями, вызывающие скольжение их относительно друг друга, что и составляет основу процесса сокращения мыщц.
8. Спустя долю секунды с помощью кальциевого насоса в мембране саркоплазматического ретикулума ионы кальция закачиваются обратно и сохраняются в ретикулуме до прихода нового потенциала действия. Удаление ионов кальция от миофибрилл ведет к прекращению мышечного сокращения.
Далее мы обсудим молекулярные механизмы этого процесса.
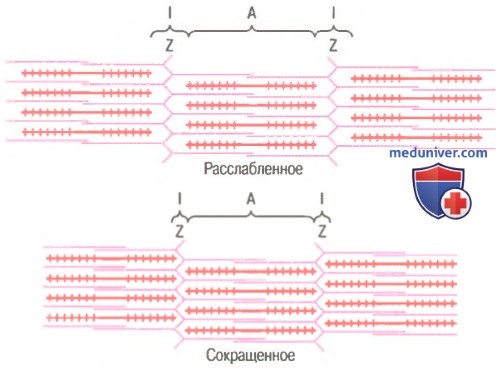
Миофибрилла в расслабленном и сокращенном состоянии. Показано (вверху), что актиновые нити (розовые) вдвинуты в пространства между миозиновыми нитями (красные). Сближение Z-дисков друг с другом (внизу).
Молекулярные механизмы мышечного сокращения
Механизм скольжения нитей для мышечного сокращения. На рисунке показан основной механизм мышечного сокращения. Показано расслабленное состояние саркомера (вверху) и сокращенное состояние (внизу). В расслабленном состоянии концы актиновых нитей, отходящие от двух последовательных Z-дисков, лишь незначительно перекрываются. Наоборот, в сокращенном состоянии актиновые нити втягиваются внутрь между миозиновыми так сильно, что их концы максимально перекрывают друг друга. При этом Z-диски притягиваются актиновыми нитями к концам миозиновых. Таким образом, мышечное сокращение осуществляется путем механизма скольжения нитей.
Что заставляет нити актина скользить внутрь среди нитей миозина? Это связано с действием сил, генерируемых при взаимодействии поперечных мостиков, исходящих от нитей миозина, с нитями актина. В условиях покоя эти силы не проявляются, однако распространение потенциала действия вдоль мышечного волокна приводит к выделению из саркоплазматическо-го ретикулума большого количества ионов кальция, которые быстро окружают миофи-бриллы. В свою очередь, ионы кальция активируют силы взаимодействия между нитями актина и миозина, в результате начинается сокращение. Для осуществления процесса сокращения необходима энергия. Ее источником являются высокоэнергетические связи молекулы АТФ, которая разрушается до АДФ с высвобождением энергии. В следующих разделах мы приведем известные детали молекулярных процессов сокращения.
Молекулярные особенности сократительных нитей
Миозиновая нить. Она состоит из множества молекул миозина, молекулярная масса каждой составляет около 480000. На рисунке показана отдельная молекула; и также — объединение многих молекул миозина в миозиновую нить, а также взаимодействие одной стороны этой нити с концами двух актиновых нитей.
В состав молекулы миозина входят 6 полипептидных цепей: 2 тяжелые цепи с молекулярной массой около 200000 каждая и 4 легкие цепи с молекулярной массой около 20000 каждая. Две тяжелые цепи спирально закручиваются вокруг друг друга, формируя двойную спираль, которую называют миозиновым хвостом. С одного конца обе цепи изгибаются в противоположных направлениях, формируя глобулярную полипептидную структуру, называемую миозиновой головкой. Таким образом, на одном конце двойной спирали молекулы миозина образуются 2 свободные головки; 4 легкие цепи также включены в состав миозиновой головки (по 2 в каждой). Они помогают регулировать функцию головки во время мышечного сокращения.
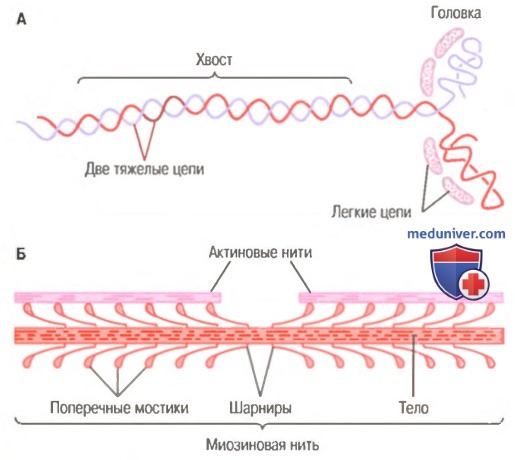
А. Молекула миозина.
Б. Объединение многих молекул миозина в одну миозиновую нить.
Показаны также тысячи миозиновых поперечных мостиков и взаимодействие их головок с прилежащими актиновыми нитями.
Миозиновая нить состоит из 200 или более отдельных молекул миозина. Видно, что хвосты молекул миозина объединяются, формируя тело нити, а многочисленные головки молекул выдаются наружу по сторонам тела. Кроме того, наряду с головкой в сторону выступает часть хвоста каждой миозиновой молекулы, образуя плечОу которое выдвигает головку наружу от тела, как показано на рисунке. Выступающие плечи и головки вместе называют поперечными мостиками. Каждый поперечный мостик может сгибаться в двух точках, называемых шарнирами. Один из них расположен в месте, где плечо отходит от тела миозиновой нити, а другой — где головка крепится к плечу. Движение плеча позволяет головке или выдвигаться далеко наружу от тела миозиновой нити, или приближаться к телу. В свою очередь, повороты головки участвуют в процессе сокращения, что обсуждается в следующих разделах.
Общая длина каждой миозиновой нити остается постоянной и равна почти 1,6 мкм. В самом центре миозиновой нити на протяжении 0,2 мкм поперечных мостиков нет, поскольку снабженные шарнирами плечи отходят в стороны от центра.
Сама миозиновая нить сплетена таким образом, что каждая последующая пара поперечных мостиков смещена в продольном направлении относительно предыдущей на 120°, что обеспечивает распределение поперечных мостиков во всех направлениях вокруг нити.
АТФ-азная активность миозиновой головки. Есть и другая особенность миозиновой головки, необходимая для мышечного сокращения: миозиновая головка функционирует как фермент АТФ-аза. Как объясняется далее, это свойство позволяет головке расщеплять АТФ и использовать энергию расщепления высокоэнергетической связи для процесса сокращения.
Актиновая нить. Актиновая нить состоит из трех белковых компонентов: актина, тропомиозина и тропонина.
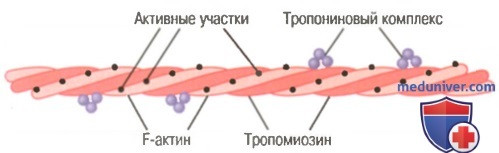
Актиновая нить, состоящая из двух спиралевидных цепочек молекул F-актина и двух цепочек молекул тропомиозина, расположенных в желобках между цепочками актина.
К одному концу каждой молекулы тропомиозина прикреплен тропониновый комплекс, который запускает сокращение.
Основой актиновой нити являются две цепи белковой молекулы F-актина. Обе цепи закручиваются в спираль так же, как и молекула миозина.
Каждая цепь двойной спирали F-актина состоит из полимеризованных молекул G-актина с молекулярной массой около 42000. К каждой молекуле G-актина прикреплена 1 молекула АДФ. Полагают, что эти молекулы АДФ являются активными участками на актиновых нитях, с которыми взаимодействуют поперечные мостики миозиновых нитей, обеспечивая мышечное сокращение. Активные участки на обеих цепях F-актина двойной спирали расположены со смещением таким образом, что вдоль всей поверхности актиновой нити встречается один активный участок примерно через каждые 2,7 нм.
Длина каждой актиновой нити — около 1 мкм. Основания актиновых нитей прочно встроены в Z-диски; концы этих нитей выступают в обоих направлениях, располагаясь в пространствах между миозиновыми молекулами.
Молекулы тропомиозина. Актиновая нить также содержит другой белок — тропомиозин. Каждая молекула тропомиозина имеет молекулярную массу 70000 и длину 40 нм. Эти молекулы спирально оплетают спираль из F-актина. В состоянии покоя молекулы тропомиозина располагаются поверх активных участков актиновых нитей, препятствуя их взаимодействию с миозиновыми нитями, лежащему в основе сокращения.
Тропонин и его роль в мышечном сокращении. По ходу молекул тропомиозина к ним периодически прикреплены другие белковые молекулы, называемые тропонином. Они представляют собой комплексы трех слабосвязанных белковых субъединиц, каждая из которых играет специфическую роль в регуляции мышечного сокращения. Одна из субъединиц (тропонин I) имеет высокое сродство к актину, другая (тропонин Т) — к тропомиозину, третья (тропонин С) — к ионам кальция. Считают, что этот комплекс прикрепляет тропомиозин к актину. Высокое сродство тропонина к ионам кальция, как полагают, инициирует процесс сокращения, о чем говорится в следующей статье.
Видео физиология мышц и мышечного сокращения - профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021

Весь контент iLive проверяется медицинскими экспертами, чтобы обеспечить максимально возможную точность и соответствие фактам.
У нас есть строгие правила по выбору источников информации и мы ссылаемся только на авторитетные сайты, академические исследовательские институты и, по возможности, доказанные медицинские исследования. Обратите внимание, что цифры в скобках ([1], [2] и т. д.) являются интерактивными ссылками на такие исследования.
Если вы считаете, что какой-либо из наших материалов является неточным, устаревшим или иным образом сомнительным, выберите его и нажмите Ctrl + Enter.
Основное свойство мышечной ткани, образующей скелетные мышцы, - сократимость приводит к изменению длины мышцы под влиянием нервных импульсов. Мышцы действуют на кости рычагов, соединяющихся при помощи суставов. При этом каждая мышца действует на сустав только в одном направлении. У одноосного сустава (цилиндрического, блоковидного) движение костных рычагов совершается только вокруг одной оси, поэтому мышцы располагаются по отношению к такому суставу с двух сторон и действуют на него в двух направлениях (сгибание - разгибание; приведение - отведение, вращение). Например, у локтевого сустава одни мышцы - сгибатели, другие - разгибатели. Друг по отношению к другу эти мышцы, действующие на сустав в противоположных направлениях, являются антагонистами. Как правило, на каждый сустав в одном направлении действуют две или более мышц. Такие содружественные по направлению действия мышцы называют синергистами. У двуосного сустава (эллипсоидный, мыщелковый, седловидный) мышцы группируются соответственно двум его осям, вокруг которых совершаются движения. К шаровидному суставу, имеющему три оси движения (многоосный сустав), мышцы прилежат с нескольких сторон и действуют на него в разных направлениях. Так, например, у плечевого сустава имеются мышцы - сгибатели и разгибатели, осуществляющие движение вокруг фронтальной оси, отводящие и приводящие - вокруг сагиттальной оси и вращатели - вокруг продольной оси (вовнутрь - пронаторы и кнаружи - супинаторы).
В группе мышц, выполняющих то или иное движение, можно выделить мышцы главные, обеспечивающие данное движение, и вспомогательные, о подсобной роли которых говорит само название. Вспомогательные мышцы моделируют движение, придают ему индивидуальные особенности.
Для функциональной характеристики мышц используются такие показатели, как их анатомический и физиологический поперечник. Анатомический поперечник - это величина (площадь) поперечного сечения, перпендикулярного длиннику мышцы и проходящего через брюшко в наиболее широкой его части. Этот показатель характеризует величину мышцы, ее толщину. Физиологический поперечник мышцы представляет собой суммарную площадь поперечного сечения всех мышечных волокон, входящих в состав исследуемой мышцы. Поскольку сила сокращающейся мышцы зависит от количества мышечных волокон, величины поперечного сечения, то физиологический поперечник мышцы характеризует ее силу. У мышц веретенообразной, лентовидной формы с параллельным расположением волокон анатомический и физиологический поперечники совпадают. Иная картина у перистных мышц, имеющих большое количество коротких мышечных пучков. Из двух равновеликих мышц, имеющих одинаковый анатомический поперечник, у перистой мышцы физиологический поперечник больше, чем у веретенообразной. Суммарное поперечное сечение мышечных волокон у перистой мышцы больше, а сами волокна короче, чем у веретенообразной. В связи с этим перистая мышца по сравнению с последней обладает большей силой, однако размах сокращения ее коротких мышечных волокон меньше. Перистые мышцы имеются там, где необходима значительная сила мышечных сокращений при сравнительно небольшом размахе движений (мышцы голени, стопы, некоторые мышцы предплечья). Мышцы веретенообразной, лентовидной формы, построенные из длинных мышечных волокон, при сокращении укорачиваются на большую величину. В то же время они развивают меньшую силу, чем перистые мышцы, имеющие одинаковый с ними анатомический поперечник.
Работа мышц. Поскольку концы мышцы прикреплены на костях, то точки ее начала и прикрепления при сокращении приближаются друг к другу, а сами мышцы при этом выполняют определенную работу. Таким образом, тело человека или его части при сокращении соответствующих мышц изменяют свое положение, приходят в движение, преодолевают сопротивление силы тяжести или, наоборот, уступают этой силе. В других случаях при сокращении мышц тело удерживается в определенном положении без выполнения движения. Исходя из этого, различают преодолевающую, уступающую и удерживающую работу мышц.
Преодолевающая работа мышц выполняется в том случае, если сила сокращения мышцы изменяет положение части тела, конечности или ее звена, с грузом или без него, преодолевая силу сопротивления.
Уступающей называют работу, при которой сила мышцы уступает действию силы тяжести части тела (конечности) и удерживаемого ею груза. Мышца работает, однако она не укорачивается при этом, а, наоборот, удлиняется; например, когда невозможно поднять или удержать на весу предмет, имеющий большую массу. При большом усилии мышц приходится опустить это тело на пол или на другую поверхность.
Удерживающая работа выполняется, если силой мышечных сокращений тело или груз удерживается в определенном положении без перемещения в пространстве. Например, человек стоит или сидит, не двигаясь, или держит груз в одном и том же положении. Сила мышечных сокращений уравновешивает массу тела или груза. При этом мышцы сокращаются без изменения их длины (изометрическое сокращение).
Преодолевающую и уступающую работу, когда сила мышечных сокращений перемещает тело или его части в пространстве, можно рассматривать как динамическую работу. Удерживающая работа, при которой движения всего тела или части тела не происходит, является работой статической.
Кости, соединенные суставами, при сокращении мышц действуют как рычаги. В биомеханике выделяют рычаг первого рода, когда точки сопротивления и приложения мышечной силы находятся по разные стороны от точки опоры, и рычаг второго рода, в котором обе силы прилагаются по одну сторону от точки опоры, на разном расстоянии от нее.
Рычаг второго рода одноплечий. В биомеханике (в отличие от механики) он бывает двух видов. Вид такого рычага зависит от места расположения точки приложения силы и точки действия силы тяжести, которые и в том, и в другом случае находятся по одну сторону от точки опоры. Первый вид рычага второго рода (рычаг силы) имеет место в том случае, если плечо приложения мышечной силы длиннее плеча сопротивления (силы тяжести). Рассматривая в качестве примера стопу, можно видеть, что точкой опоры (ось вращения) служат головки костей плюсны, а точкой приложения мышечной силы (трехглавой мышцы голени) является пяточная кость. Точка сопротивления (тяжесть тела) приходится на место сочленения костей голени со стопой (голеностопный сустав). В этом рычаге отмечаются выигрыш в силе (плечо приложения силы длиннее) и проигрыш в скорости перемещения точки сопротивления (ее плечо короче). У второго вида одноплечевого рычага (рычага скорости) плечо приложения мышечной силы короче, чем плечо сопротивления, где приложена противодействующая сила, сила тяжести. Для преодоления силы тяжести, точка приложения которой отстоит на значительном расстоянии от точки вращения в локтевом суставе (точка опоры), необходима значительно большая сила мышц-сгибателей, прикрепляющихся вблизи от локтевого сустава (в точке приложения силы). При этом наблюдаются выигрыш в скорости и размахе движения более длинного рычага (точка сопротивления) и проигрыш в силе, действующей в точке приложения этой силы.
Механизм мышечного сокращения и расслабления заключается в перемещении нитей актина относительно нитей миозина. Нити актина двигаются как по туннелю, между миозиновыми фибриллами, за счет чего волокно укорачивается. Такое сокращение, сопровождающееся изменением длины, называют изотоническим. Тип сокращения, который осуществляется при неизменной длине, называют изометрическим. Энергию для перемещения нитей дает АТФ.
Сила сокращения мышц зависит от количества нейромоторных единиц, участвующих в этом сокращении, от частоты раздражения, до известного предела. Максимальное напряжение, которое может развить мышца, определяется числом образующих ее волокон: чем оно больше, тем больше сила мышц. Поэтому перистые мышцы, в которых велико число волокон, отличаются большой силой. Проявление силы зависит от особенностей прикрепления мышцы к костям. Мышцы с большей площадью опоры имеют большие возможности для проявления силы.
Мышца, сокращаясь, производит работу. Величина ее равна произведению массы груза на поднятую высоту. Отсюда следует, что максимальная работа, выполняемая при одиночном сокращении мышцы, зависит от ее силы (чем больше сила, тем больший груз может быть поднят) и степени укорочения мышцы.
В процессе естественной деятельности человека величина работы, выполняемой той или иной мышцей, в значительной степени зависит от способности ее длительно находиться в сокращенном состоянии, т. е. от степени выносливости мышц. Различают выносливость к статическим и динамическим усилиям. Выносливость к статическим усилиям определяется временем, в течение которого удерживается величина заданного усилия. Она различна для различных мышц. Наименьшей выносливостью характеризуется трехглавая мышца плеча (1 мин при усилии, равном 50 % от максимального), наибольшей — икроножная мышца (7 мин). Выносливость к динамической работе зависит как от величины поднимаемого груза, так и от темпа работы. Работа бывает наибольшей при какой-то средней величине груза и частоте движений. Выносливость к динамической и статической работе можно увеличивать путем тренировки. При длительной как динамической, так и статической работе наступает утомление мышц.
Работа мышц — необходимое условие их существования. Длительная бездеятельность мышц ведет к их атрофии и потере ими работоспособности. Тренировка, т. е. систематическая, нечрезмерная работа мышц, способствует увеличению их объема, возрастанию силы и работоспособности, что важно для физического развития всего организма.
Глава 7. Возрастная анатомия и физиология системы крови
7.1. Общая характеристика крови
Кровь вместе с кроветворными и кроверазрушающими органами составляет целостную систему крови, которая включает костный мозг, селезенку, тимус, лимфатические узлы, миндалины и отдельные лимфоидные фолликулы слизистых оболочек дыхательных, пищеварительных и мочеполовых путей. У взрослого человека общее количество крови составляет 5—8 % веса тела, что соответствует 5—6 л. Кровь имеет красный цвет и рН 7,35. Она является жидкой тканью и разновидностью соединительной ткани, находится в непрервывном движении, располагается в замкнутой системе кровеносных сосудов. Кровь вместе с лимфой и межтканевой жидкостью составляет внутреннюю среду организма, в которой протекает жизнедеятельность всех клеток и тканей. Особенностью крови является ее высокая способность к регенерации, которая происходит в специальных кроветворных органах и носит название кроветворения. Кровь выполняет следующие функции: транспортную, защитную, терморегуляторную. Транспортная функция заключается в переносе питательных веществ (глюкоза, аминокислоты, жиры и др.) к клеткам, а конечные продукты обмена веществ (аммиак, мочевину, мочевую кислоту и др.) — от них к органам выделения. Осуществляя перенос гормонов и других физиологически активных веществ, воздействующих на различные органы и ткани, она участвует в выполнении регуляторной функции. С функциями крови тесно связана регуляция постоянства температуры тела. Кровь переносит тепло от органов с интенсивным его образованием к органам с менее интенсивной теплопродукцией и к местам, где она охлаждается (поверхность кожи). Защитную функцию кровь выполняет благодаря способности лейкоцитов к фагоцитозу и наличию в ней иммунных тел, обезвреживающих микроорганизмы и их яды, разрушающих чужеродные белки. Доставляя кислород от легких к тканям, принося к легким углекислый газ, кровь осуществляет дыхательную функцию.
В кровеносных сосудах в состоянии покоя циркулирует не вся кровь. Около 40—50 % ее находится в кровяных депо (селезенке, печени, сосудах кожи и легких). Депонированная кровь в единице объема содержит больше форменных элементов по сравнению с кровью, циркулирующей в сосудах. Выход крови из депо в общий кровоток обусловливается многими факторами. Наиболее важным из них является дефицит кислорода в организме, возникающий под влиянием различных причин (мышечная деятельность, кровопотери, пониженное атмосферное давление и др.). Уменьшение количества крови более чем на 1/3 опасно для жизни. Потеря крови в небольших количествах (200—400 мл) для здоровых людей не только не представляет вреда, но даже стимулирует кроветворение.
Кровь состоит из форменных элементов (55—58 %) — эритроцитов, лейкоцитов и тромбоцитов (рис. 15), и жидкой части — плазмы (42—45 %).
Самсонова, А.В. Биомеханика мышц [Текст]: учебно-методическое пособие / А.В.Самсонова, Е.Н. Комисарова; Под ред. А.В.Самсоновой; СПбГУФК им. П.Ф.Лесгафта. – СПб.: [б.и.], 2008. – 127 с.
Самсонова А.В., Комиссарова Е.Н.
БИОМЕХАНИКА МЫШЦ
В учебно-методическом пособии рассмотрены теоретические и практические аспекты биомеханики мышечной деятельности: строение мышц с точки зрения биомеханики; механика мышечного сокращения; зависимость силы и скорости сокращения мышц от анатомических, физиологических и биомеханических факторов; результирующее действие мышц в организме. Пособие содержит большой фактический материал из практики спорта.
Более подробно функционирование опорно-двигательного аппарата человека и биомеханика мышц описаны в книге:
Самсонова А.В. – главы: 2, 3, 4, 5, 6
Комиссарова Е.Н. – глава 1, глоссарий
СОДЕРЖАНИЕ
Глава 1. Архитектура скелетных мышц
1.2. Макроструктура мышцы
1.3. Микроструктура мышцы
1.5. Теория скользящих нитей
1.6. Состояние мышцы
1.7. Типы скелетных мышечных волокон и их морфофункциональная характеристика
1.8. Влияние различных факторов на состав мышечных волокон
1.9. Контрольные вопросы
Глава 2. Функционирование рецепторного аппарата мышц и суставов
2.1. Рецепторы опорно-двигательного аппарата человека
2.1.1. Мышечные веретена
2.1.2. Рецепторы Гольджи
2.1.3. Рецепторы суставов
2.2. Зависимость частоты импульсации рецепторов от длины, скорости и напряжения мышцы
2.2.3. Зависимость частоты импульсации рецепторов Гольджи от степени напряжения мышцы
2.3. Способы оценки афферентного притока, поступающего от рецепторов мышц
2.4. Уровни построения движений и рецепторы опорно-двигательного аппарата
2.5. Контрольные вопросы
Глава 3. Механика мышечного сокращения
3.1. Биомеханические свойства мышц
3.2. Трехкомпонентная модель мышцы
3.3. Функционирование биомеханической модели мышцы в простейших двигательных задачах
3.4. Контрольные вопросы
Глава 4. Факторы, определяющие силу и скорость сокращения мышцы
4.1. Основные понятия
4.2. Анатомические факторы, определяющие силу и скорость сокращения мышц
4.2.1. Сила и скорость сократительного компонента мышцы
4.2.2. Сила и скорость сокращения мышцы в целом
4.3. Физиологические факторы, определяющие силу и скорость сокращения мышц
4.3.1. Физиологические механизмы регуляции силы и скорости сокращения мышцы
4.3.2. Время сокращения мышцы
4.4. Биомеханические факторы, определяющие силу и скорость сокращения мышц
4.4.1. Длина мышцы
4.4.2. Характер работы мышц
4.4.3. Значение внешней силы
4.5. Контрольные вопросы
Глава 5. Результирующее действие мышц в организме
5.1. Звенья тела как рычаги
5.2. Методы определения морфометрических характеристик мышц нижних конечностей человека
5.2.1. Моделирование ОДА человека и мышц нижних конечностей
5.2.2. Рентгенографический метод определения морфометрических характеристик мышц
5.2.3. Анатомический метод определения морфометрических характеристик мышц
5.2.4. Биомеханический метод определения морфометрических характеристик мышц
5.3. Фазовые траектории мышц &? способ представления результатов, характеризующих моторную функцию мышц
5.4. Программа расчета морфометрических характеристик мышц MORFOMETR
5.5. Контрольные вопросы
Глава 6. Функционирование мышц в спортивных движениях
6.1. Биомеханический анализ физических упражнений
6.2. Обучение двигательным действиям
6.3. Классификация физических упражнений
6.4. Сравнение основного и специальных упражнений
6.5. Оценка функциональной подготовленности спортсменов на основе анализа фазовых портретов мышц
6.6. Контрольные вопросы
ВЫДЕРЖКИ ИЗ КНИГИ
ВВЕДЕНИЕ
Авторы стремились изложить материал предельно просто и доступно. В связи с этим, пособие содержит большое количество иллюстраций, а в конце пособия помещен глоссарий. Отзывы об учебно-методическом пособии просим отправлять по адресу:
ГЛАВА 2
ФУНКЦИОНИРОВАНИЕ РЕЦЕПТОРНОГО АППАРАТА МЫШЦ И СУСТАВОВ
2.1. Рецепторы опорно-двигательного аппарата человека
При изучении анатомии и физиологии (А.С. Солодков, Е.Б. Сологуб, 2001) вы изучали двигательную сенсорную систему. Одним из отделов этой системы являются проприорецепторы, расположенные в мышцах, сухожилиях и суставных сумках. В мышцах расположены мышечные веретена, в сухожилиях – сухожильные органы Гольджи. В суставных сумках расположены рецепторы суставов.
2.1.1. Мышечные веретена
Еще в XIX веке В. Кюне обнаружил в скелетных мышцах структуры, напоминающие веретено. Затем, в начале XX века Нобелевский лауреат Чарльз Скотт Шеррингтон показал, что эти структуры служат чувствительными рецепторами. Мышечные веретена рассеяны по всем скелетным мышцам. Концы их обычно прикрепляются к мышечным волокнам параллельно. Каждое веретено покрыто капсулой, которая расширятся в центре и образует ядерную сумку. Внутри веретена содержатся интрафузальные мышечные волокна. Эти волокна в 2-3 раза тоньше обычных (экстрафузальных) волокон скелетных мышц.
Интрафузальные волокна подразделяются на два типа:
- Длинные и толстые (диаметр 20-25 мкм), которые информируют ЦНС о динамическом компоненте движения – скорости изменения длины мышцы. Таких волокон в мышечном веретене не более двух.
- Короткие и тонкие (диаметр 10–12 мкм), которые информируют ЦНС о статическом компоненте движения – текущей длине мышцы. Таких волокон в мышечном веретене от 2 до 12.
2.1.2. Рецепторы Гольджи
Нервно-сухожильные веретена (рецепторы Гольджи) открыл в 1903 году Камилло Гольджи. Впоследствии за эти исследования ему была присуждена Нобелевская премия. Рецепторы Гольджи располагаются в месте перехода мышечных волокон в сухожилия. Их длина составляет 0,5-1,0 мм, а диаметр – 0,1- 0,2 мм. Отдельный нервный аксон несет афферентные импульсы в спинной мозг и называется аксоном Ib. Он начинается в виде веточек, проходящих между коллагеновыми волокнами сухожилия (рис. 2.1а). Когда мышечные волокна сокращаются, коллагеновые волокна натягиваются и сжимают нервные веточки, которые начинают импульсировать (рис. 2.1б). Таким образом, в результате последовательного крепления сухожильных органов к мышечным волокнам они возбуждаются при укорочении возбужденной мышцы. Сухожильные рецепторы возбуждаются в 1,5 – 8 раз более эффективно при мышечном сокращении, нежели при пассивном растяжении.
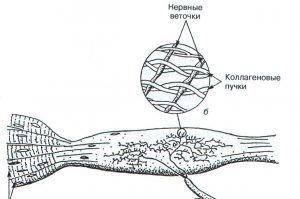
Рис. 2.1. Строение сухожильного органа Гольджи (А.Дж. Мак-Комас, 2001)
2.1.3. Рецепторы суставов
Суставные рецепторы подразделяются на несколько типов в зависимости от их реакции на амплитуду, скорость и направление движения в суставе.
Тельца Руффини находятся в капсуле сустава и воспринимают направление и скорость изменения межзвенного угла. Частота их импульсации возрастает с увеличением скорости изменения суставного угла.
Тельца Паччини посылают в ЦНС информацию о положении отдельных частей тела в пространстве и относительно друг друга. Эти рецепторы посылают в ЦНС информацию о значениях межзвенных углов, то есть о положении сустава. Их импульсация продолжается в течение всего периода сохранения межзвенного угла, и она тем больше, чем больше изменения угла.
2.2. Зависимость частоты импульсации рецепторов от длины, скорости и напряжения мышцы
Реакция мышечных веретен на активное или пассивное укорочение мышцы была предсказана в 1928 году Дж. Фултоном и Дж. Писуньери на основе анатомического анализа. Поскольку веретена располагаются параллельно мышечным волокнам, частота разрядов веретенных афферентов при любом укорочении мышцы должна снижаться. В последующем это предположение полностью подтвердилось. Исследования свойств изолированных мышечных веретен, проведенные лауреатом Нобелевской премии Бернардом Катцем (B. Katz, 1950) продемонстрировали, что их растяжение приводит к деполяризации окончаний афферентных волокон. Величина деполяризации при растяжении увеличивается. При этом зависимость частоты импульсации веретенных афферентов от растяжения мышцы близка к линейной. Эту зависимость принято называть статическим ответом веретенного афферента на пассивное растяжение мышцы.
Исследования свойств мышечных веретен свидетельствуют о том, что активность первичных окончаний чувствительного нерва зависит не только от длины, но и от скорости растяжения мышцы. Способность менять частоту своей импульсации в зависимости от скорости удлинения мышцы была названа динамической чувствительностью веретенных афферентов. Зависимость между скоростью растяжения мышц и частотой импульсации первичного афферента также близка к линейной.
2.2.3. Зависимость частоты импульсации рецепторов Гольджи от степени напряжения мышцы
Исследования, проведенные на свободно перемещающихся животных в условиях стационарного режима локомоции, показали, что кривая, отражающая изменение частоты импульсации рецепторов Гольджи во времени полностью соответствует огибающей электромиограммы (рис. 2.2). При этом частота импульсации не превышает 200 имп/с.
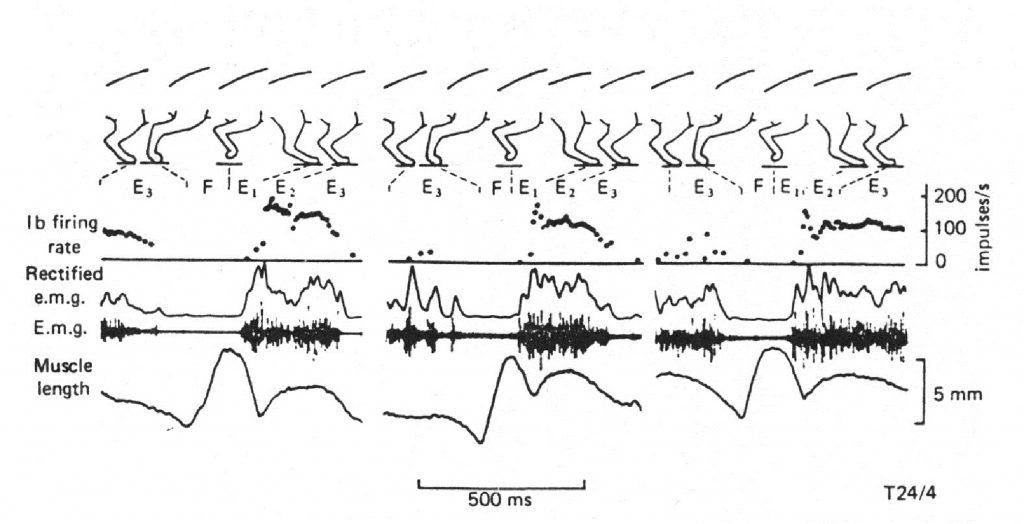
Рис. 2.2. Зависимость частоты импульсации рецепторов Гольджи от степени напряжения мышцы при локомоции свободно перемещающейся кошки
Таким образом, рецепторы мышц адекватно реагируют на изменение длины и скорости растяжения мышцы. Связь между этими характеристиками и частотой импульсации афферентов мышечных веретен близка к линейной. Рецепторы Гольджи адекватно отражают развитие напряжения мышцы. Рецепторы суставов реагируют на положение и угловую скорость звеньев опорно-двигательного аппарата.
2.3. Способы оценки афферентного притока, поступающего от рецепторов мышц
С начала XX века и до настоящего времени накоплен богатый материал о свойствах мышечных рецепторов. В основном эти данные были получены на наркотизированных или другим способом обездвиженных животных. Затем исследования были продолжены на децеребрированных животных. Последующие эксперименты, проведенные с помощью вживленных электродов и телеметрической передачи сигналов, убедительно доказали, что у свободно перемещающегося животного разряды первичных афферентов проявляют высокую активность в фазе пассивного растяжения и очень низкую – в фазе активного укорочения.
Одновременно с проведением экспериментов на животных импульсация рецепторов мышц стала изучаться на человеке. С этой целью была разработана методика микронейрографии, суть которой заключается в регистрации афферентной активности мышц посредством тонкого игольчатого электрода, введенного в нерв. Это позволило регистрировать потенциалы действия в нерве у человека при выполнении изометрических напряжений и даже произвольных движений. Было отмечено, что непрерывная афферентная активность возникала при пассивном растяжении мышцы. При быстром движении афферентная активность уменьшалась на время укорочения мышцы. Следует, однако, отметить, что использование этой методики невозможно при исследовании быстрых, мощных высокоамплитудных движений, какими являются движения спортсменов. В настоящее время разработана методика оценки афферентной активности мышц посредством регистрации ВПСМ (вызванных потенциалов спинного мозга). Однако ее особенности также не позволяют использовать этот способ для регистрации афферентного притока при спортивных движениях. Это связано с тем, что помехи, возникающие со стороны других органов тела (сердца, мышц спины), на несколько порядков выше, чем проявляемый сигнал.
Наряду с разработкой методик, позволяющих напрямую регистрировать разряды рецепторов мышц, существуют исследования, моделирующие работу рецепторного аппарата мышц. W.Z. Rymer, J.С. Houk, P.E. Crago (1977) предложили формулу для описания зависимости частоты разрядов мышечных афферентов от степени удлинения и скорости сокращения мышц. В модели, предложенной S.S. Schafer и S. Schafer, (1969) частота разрядов мышечных афферентов зависит не только от удлинения и скорости сокращения мышцы, но и от ускорения.
Методика, позволяющая косвенно судить о функционировании рецепторного аппарата мышц и суставов при выполнении спортивных движений, разработана А.В. Самсоновой (1997). При выполнении двигательных действий можно зарегистрировать изменение межзвенных углов и электрическую активность мышц. Предлагаемая методика дает возможность в каждый момент времени иметь информацию об изменении длины мышцы и скорости ее сокращения. Кроме того, методика позволяет рассчитать значения межзвенных углов и угловое ускорение.
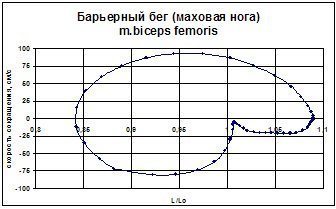
Рис. 2.3. Фазовая траектория двуглавой м. бедра при преодолении барьера спортсменкой высокой квалификации
Рис. 2.3. Фазовая траектория двуглавой м. бедра при преодолении барьера спортсменкой высокой квалификации
2.4. Уровни построения движений и рецепторы опорно-двигательного аппарата
Афферентация уровня А основана на импульсации мышечных веретен (длина и скорость сокращения мышцы) и рецепторов Гольджи (уровень возбуждения мышцы при ее укорочении). Эта информация очень слабо осознается ЦНС, то есть, по гипотезе Н.А.Бернштейна, этот уровень почти никогда не бывает ведущим.
Афферентация уровня В опирается на информацию, поступающую от суставных рецепторов. Это уровень выступает как ведущий в ряде физических упражнений, таких как наклоны тела вперед и назад, а также циклические движения. Сигналы от суставных рецепторов хорошо осознаются.
2.5. Контрольные вопросы
- Какие рецепторы расположены в мышцах?
- Какие рецепторы расположены в суставах?
- Как называются мышечные волокна, расположенные в мышечных веретенах?
- Какую информацию несут в ЦНС мышечные веретена?
- Охарактеризуйте функционирование рецепторов Гольджи.
- Дайте характеристику рецепторам суставов. О каких изменениях они несут информацию в ЦНС?
- Информация каких рецепторов хорошо осознается ЦНС, а каких — плохо?
РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА
ОСНОВНАЯ ЛИТЕРАТУРА
ДОПОЛНИТЕЛЬНАЯ ЛИТЕРАТУРА
- Бочаров, А.Ф. Биомеханика: Учебное пособие [Текст] / А.Ф. Бочаров, Г.П. Иванова, В.П. Муравьев. – СПб. [б.и.]: СПбГАФК им. П.Ф. Лесгафта, 2000. – 74 с.
- Донской, Д.Д. Биомеханика: Учеб. для ин-тов физ. культуры [Текст]/ Д.Д. Донской, В.М. Зациорский. – М.: Физкультура и спорт, 1979. – 264 с.
- Иваницкий, М.Ф. Анатомия человека (с основами динамической морфологии): Учеб. для ин-тов физ. культуры [Текст] / Под ред. Б.А. Никитюка, А.А. Гладышевой, Ф.В. Судзиловского. – М.: Физкультура и спорт, 1985. – 544 с.
- Козлов И.М. Биомеханические факторы организации спортивных движений: монография [Текст] /И.М.Козлов Санкт-Петербургская гос. академия физ. культуры им. П.Ф.Лесгафта – СПб, [б.и.], 1998.– 141 с.
- Коренберг, В.Б. Спортивная биомеханика. Словарь-справочник: Учебное пособие [Текст] / В.Б. Коренберг. – Малаховка [б.и.]: МГАФК, 1999. – 192 с.
- Мак-Комас Алан. Дж. Скелетные мышцы. Строение и функции [Текст] /Алан Дж. Мак-Комас.&? Киев: Олимпийская литература, 2001.– 407 с.
- Михайлов С.С. Спортивная биохимия: учебник для вузов и колледжей физической культуры [Текст] / С.С.Михайлов; СПбГУФК им. П.Ф.Лесгафта, СПб, [б.и.], 2006. – 230 с.
- Петров, В.А. Механика спортивных движений [Текст]./ Петров В.А., Гагин Ю.А. М.: Физкультура и спорт, 1974.– 232 с.
- Солодков А.С., Физиология человека. Общая. Спортивная. Возрастная: Учебник [Текст]/ Солодков А.С., Сологуб Е.Б.– М.: Терра-Спорт, Олимпия пресс, 2001.– 520 с. ил.
- Теория и методика физической культуры [Текст] / Под ред. проф. Ю.Ф.Курамшина.– М.: Советский спорт, 2004.–463 с.
- Энока Р.М. Основы кинезиологии [Текст]. – Киев: Олимпийская литература, 1998.– 399 с.
Как приобрести
Похожие записи:
Effect of KAATSU-training on the maximum voluntary isometric contraction of lower extremity muscles of qualified football players
Изучалось влияние KAATSU-тренинга на изометрическую силу мышц квалифицированных футболистов. Установлено, что интенсивный рост максимальной силы мышц…
Удаление фасции снижает силу мышц
В опытах на диких индейках показано, что удаление фасции снижает силу мышц на 30%. Эти результаты ставят под…
Искусственный интеллект в спортивной тренировке
Описана система комплексного контроля в спорте, построенная на основе искусственного интеллекта. Разработанная система внедрена в подготовку пауэрлифтеров высокой…
Обучение двигательным действиям без ошибок
Учебное пособие доктора педагогических наук, профессора, заведующего кафедрой теории и методики адаптивной физической культуры НГУ им. П.Ф. Лесгафта,…
Гормоны и гипертрофия скелетных мышц
В пособии представлены современные фактические данные о теории и практике увеличения массы скелетных мышц под…
Тейпирование в спорте — книга
Учебное пособие канд. мед. наук, доцента, профессора кафедры спортивной медицины и технологий здоровья НГУ им. П.Ф. Лесгафта Валерия…
Читайте также: