
Механизм работы кофермента трансаминаз кратко
Обновлено: 02.07.2024
Эта реакция заключается в том, что аминокислота и кетокислота обмениваются друг с другом своими функциональными группами при альфа-углеродном атоме. В результате вступившая в реакцию аминокислота превращается в соответствующую альфа-кетокислоту, а кетокислота становится аминокислотой.
Эту реакцию катализируют ферменты под названием ТРАНСАМИНАЗЫ (АМИНОТРАНСФЕРАЗЫ). Коферментом всех трансаминаз является активная форма витамина В6 - пиридоксальфосфат (фосфопиридоксаль).
ГЛАВНЫЕ ОСОБЕННОСТИ РЕАКЦИИ ТРАНСАМИНИРОВАНИЯ:
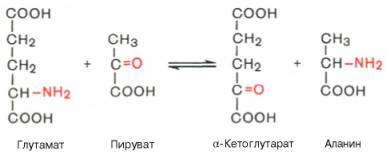
1. Это циклический процесс, все стадии которого катализируются одним и тем же ферментом - трансаминазой данной пары кислот. В этот цикл вступает одна аминокислота и кетокислота (в приведенном примере - это глутамат и ПВК). Образуются другая альфа-кетокислота и аминокислота (в приведенном примере - альфа-кетоглутарат и аланин).
3. Каждая трансаминаза обычно специфична для одной пары субстратов и соответствующей ей пары продуктов и все стадии реакции катализируются только одним ферментом. По участникам процесса для данного фермента дают ему название:
Фермент, который катализирует приведенную реакцию, можно назвать: Аланинаминотрансфераза (АлТ) (глутаминово-пировиноградная трансаминаза (ГПТ)).
4. Трансаминазы обычно обладают высокой субстратной специфичностью и высокой активностью. Наиболее активными в клетках являются те трансаминазы, для которых хотя бы один из субстратов является дикарбоновой альфа-кетокислотой - это АлТ - аланинаминотрансфераза (ГПТ) и АсТ - аспартатаминотрансфераза (ГЩТ).
Определение активности этих ферментов в сыворотке крови имеет важное значение в ДИФФЕРЕНЦИАЛЬНОЙ ДИАГНОСТИКЕ таких заболеваний, как ВИРУСНЫЙ ГЕПАТИТ и ИНФАРКТ МИОКАРДА.
АсТ и АлТ являются внутриклеточными ферментами. Поэтому в норме их активность в крови очень мала. При вирусном гепатите или при инфаркте миокарда наблюдается разрушение клеток печени или миокарда соответственно. Поэтому в крови будет наблюдаться повышение активности и АлТ, и АсТ, но неодинаковое для каждого из ферментов.
В клетках сердечной мышцы (миокарда) содержится гораздо больше АсТ, чем АлТ, а в клетках печени - наоборот: АлТ намного больше, чем АсТ. Поэтому ПРИ ИНФАРКТЕ МИОКАРДА АКТИВНОСТЬ АСТ в крови БУДЕТ ЗНАЧИТЕЛЬНО ВЫШЕ, ЧЕМ АЛТ, А ПРИ ВИРУСНОМ ГЕПАТИТЕ АКТИВНОСТЬ АЛТ БУДЕТ ВЫШЕ, ЧЕМ АСТ.
Поэтому определение активности этих ферментов в крови помогает врачам в постановке правильного диагноза.
БИОЛОГИЧЕСКАЯ РОЛЬ РЕАКЦИЙ ТРАНСАМИНИРОВАНИЯ.
1. ОБЕСПЕЧИВАЮТ СИНТЕЗ НОВЫХ АМИНОКИСЛОТ ИЗ ЧИСЛА ЗАМЕНИМЫХ. Из заменимых аминокислот также могут образоваться необходимые клетке кетокислоты.
Эта функция позволяет ругулировать содержание различных аминокислот в клетках организма (корректировка аминокислотного состава клеток).
В основе этой роли - полная обратимость реакции трансаминирования. Для характеристики направления каждой конкретной реакции трансаминирования и определения положения равновесия данной реакции вычисляют ее КОНСТАНТУ РАВНОВЕСИЯ (Кравновесия):
Для большинства реакций трансаминирования эта константа равновесия близка к единице.

При избытке какой-либо из кислот соотношение быстро восстанавливается трансаминазой за счет другой пары. Недостающее количество какой-либо из кислот может быть взято клеткой из других метаболических путей (например, альфа-кетоглутарат может быть взят из ЦТК). Избыток какой-либо из кислот может быть ликвидирован другими ферментами (например, избыточное количество глутамата окисляется глутаматдегидрогеназой).
2.ОБЕСПЕЧИВАЮТ ПРОТЕКАНИЕ РЕАКЦИЙ КОСВЕННОГО ДЕЗАМИНИРОВАНИЯ.
3.ОБЕСПЕЧИВАЮТ СИНТЕЗ МОЧЕВИНЫ
КОСВЕННОЕ ДЕЗАМИНИРОВАНИЕ (ТРАНСДЕЗАМИНИРОВАНИЕ).
Оксидазы аминокислот в клетках организма человека имеют очень низкую активность. Поэтому прямое окислительное дезаминирование аминокислот почти не протекает. Исключением является никотинамидный фермент глутаматдегидрогеназа, который легко дезаминирует глутаминовую кислоту, расщепляя ее до NH3 и альфа-кетоглутарата. Альфа-кетоглутарат может легко вступать в реакции трансаминирования с другими аминокислотами, превращаясь обратно в глутаминовую кислоту.

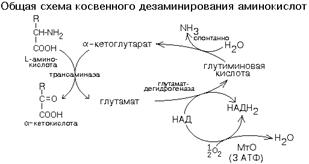
Значит, если аминокислота не может дезаминироваться прямо, то она может дезаминироваться косвенно с участием пары "альфа-кетоглутарат/глутамат".
Некоторые аминокислоты не имеют "своей собственной" трансаминазы, которая могла бы перенести их аминогруппу на альфа-кетоглутарат (лизин, треонин, пролин, оксипролин), но для этих аминокислот есть специальные реакции, в результате которых аминогруппа этих аминокислот оказывается в составе глутаминовой кислоты.
Таким образом, глутаминовая кислота работает в паре с альфа-кетоглутаровой: альфа-кетоглутарат принимает у разных аминокислот азот в форме аминогруппы, превращаясь в глутамат; глутамат легко дезаминируется прямым путем высокоактивной глутаматдегидрогеназой с образованием аммиака.
Общий итог косвенного дезаминирования такой же, как и результат прямого окислительного дезаминирования.
Это вариант дезаминирования, который протекает в две стадии:
а) трансаминирование с участием альфа-кетоглутаровой кислоты;
б) дезаминирование образовавшейся на первой стадии глутаминовой кислоты.
Косвенное дезаминирование является наиболее важным, т.к. именно таким путем дезаминируются большинство аминокислот живого организма. Это обусловлено тем, что фермент глутамат-ДГ более активен, чем оксидазы аминокислот.
ПРЕИМУЩЕСТВА КОСВЕННОГО ДЕЗАМИНИРОВАНИЯ ПЕРЕД ПРЯМЫМ:
1. В этом процессе не образуется Н2О2, в отличие от прямого дезаминирования, а образуется Н2О и 3 молекулы АТФ (энергия).
2. Косвенное дезаминирование обеспечивает гораздо более быстрое дезаминирование (благодаря высокой активности фермента глутамат-ДГ и трансаминазы);
3. Косвенное дезаминирование протекает не только быстрее, но скорость этого процесса может регулироваться. Возможность регуляции косвенного дезаминирования обусловлена обратимостью процесса и автономной саморегуляцией. Аллостерическими регуляторами глутаматдегидрогеназы являются АДФ (сильный активатор) и АТФ (ингибитор). Ингибирующее действие на этот фермент оказывают также ГТФ и НАДН2.
Если АТФ начинает расходоваться больше, чем образуется в процессе биоокисления в митохондриях, то после активации глутамат-ДГ происходит смещение реакций в сторону образования АТФ, благодаря тому, что глутамат начинает быстрее дезаминироваться, а образующиеся кетокислоты, либо сразу, либо после определенных превращений, могут также вступать на путь митохондриального окисления. Это наблюдается при недостатке жирных кислот или углеводов. То есть при недостатке жиров и углеводов в пище, в качестве дополнительного источника энергии организм начинает использовать аминокислоты. Нередко приходится при голодании разрушать собственные белки. Т.е. при похудании в организме разрушаются не только жиры и углеводы, но и собственные белки. Присутствие достаточного количества жиров и углеводов в пище позволяет сберечь белки от нерационального использования.
СИНТЕЗ ЗАМЕНИМЫХ АМИНОКИСЛОТ В ОРГАНИЗМЕ
Иногда в организме возникает ситуация, когда потребность в тех или иных аминокислотах больше, чем их поступление с пищей. Допустим, организму требуется 7 граммов аланина и 15 граммов глутамата, а с пищей поступило всего лишь 5 граммов аланина, но зато 20 граммов глутамата. Тогда организм, используя механизм трансаминирования, может синтезировать недостающее количество аланина из ПВК, который синтезируется в бих количествах (сотни граммов в сутки). Таким образом, в организме постоянно происходит корректировка аминокислотного состава, но это не уменьшает потребности организма в пищевом белковом азоте. Эта корректировка осуществляется с использование соответствующих альфа-кетокислот, образующихся в ходе метаболизма углеводов и жиров.
Незаменимые аминокислоты не могут образуются в организме, потому что в ходе распада углеводов и жиров не образуется соответствующих им альфа-кетокислот.
Подводя итог, можно сказать, что в результате общих путей катаболизма аминокислот, они разрушаются до СО2, NH3 и безазотистых фрагментов, которые также могут разрушаться до СО2 и Н2О. Аммиак подвергается реакциям обезвреживания.
Эта реакция заключается в том, что аминокислота и кетокислота обмениваются друг с другом своими функциональными группами при альфа-углеродном атоме. В результате вступившая в реакцию аминокислота превращается в соответствующую альфа-кетокислоту, а кетокислота становится аминокислотой.
Эту реакцию катализируют ферменты под названием ТРАНСАМИНАЗЫ (АМИНОТРАНСФЕРАЗЫ). Коферментом всех трансаминаз является активная форма витамина В6 - пиридоксальфосфат (фосфопиридоксаль).
ГЛАВНЫЕ ОСОБЕННОСТИ РЕАКЦИИ ТРАНСАМИНИРОВАНИЯ:
1. Это циклический процесс, все стадии которого катализируются одним и тем же ферментом - трансаминазой данной пары кислот. В этот цикл вступает одна аминокислота и кетокислота (в приведенном примере - это глутамат и ПВК). Образуются другая альфа-кетокислота и аминокислота (в приведенном примере - альфа-кетоглутарат и аланин).
3. Каждая трансаминаза обычно специфична для одной пары субстратов и соответствующей ей пары продуктов и все стадии реакции катализируются только одним ферментом. По участникам процесса для данного фермента дают ему название:
Фермент, который катализирует приведенную реакцию, можно назвать: Аланинаминотрансфераза (АлТ) (глутаминово-пировиноградная трансаминаза (ГПТ)).
4. Трансаминазы обычно обладают высокой субстратной специфичностью и высокой активностью. Наиболее активными в клетках являются те трансаминазы, для которых хотя бы один из субстратов является дикарбоновой альфа-кетокислотой - это АлТ - аланинаминотрансфераза (ГПТ) и АсТ - аспартатаминотрансфераза (ГЩТ).
Определение активности этих ферментов в сыворотке крови имеет важное значение в ДИФФЕРЕНЦИАЛЬНОЙ ДИАГНОСТИКЕ таких заболеваний, как ВИРУСНЫЙ ГЕПАТИТ и ИНФАРКТ МИОКАРДА.
АсТ и АлТ являются внутриклеточными ферментами. Поэтому в норме их активность в крови очень мала. При вирусном гепатите или при инфаркте миокарда наблюдается разрушение клеток печени или миокарда соответственно. Поэтому в крови будет наблюдаться повышение активности и АлТ, и АсТ, но неодинаковое для каждого из ферментов.
В клетках сердечной мышцы (миокарда) содержится гораздо больше АсТ, чем АлТ, а в клетках печени - наоборот: АлТ намного больше, чем АсТ. Поэтому ПРИ ИНФАРКТЕ МИОКАРДА АКТИВНОСТЬ АСТ в крови БУДЕТ ЗНАЧИТЕЛЬНО ВЫШЕ, ЧЕМ АЛТ, А ПРИ ВИРУСНОМ ГЕПАТИТЕ АКТИВНОСТЬ АЛТ БУДЕТ ВЫШЕ, ЧЕМ АСТ.
Поэтому определение активности этих ферментов в крови помогает врачам в постановке правильного диагноза.
БИОЛОГИЧЕСКАЯ РОЛЬ РЕАКЦИЙ ТРАНСАМИНИРОВАНИЯ.
1. ОБЕСПЕЧИВАЮТ СИНТЕЗ НОВЫХ АМИНОКИСЛОТ ИЗ ЧИСЛА ЗАМЕНИМЫХ. Из заменимых аминокислот также могут образоваться необходимые клетке кетокислоты.
Эта функция позволяет ругулировать содержание различных аминокислот в клетках организма (корректировка аминокислотного состава клеток).
В основе этой роли - полная обратимость реакции трансаминирования. Для характеристики направления каждой конкретной реакции трансаминирования и определения положения равновесия данной реакции вычисляют ее КОНСТАНТУ РАВНОВЕСИЯ (Кравновесия):
Для большинства реакций трансаминирования эта константа равновесия близка к единице.
При избытке какой-либо из кислот соотношение быстро восстанавливается трансаминазой за счет другой пары. Недостающее количество какой-либо из кислот может быть взято клеткой из других метаболических путей (например, альфа-кетоглутарат может быть взят из ЦТК). Избыток какой-либо из кислот может быть ликвидирован другими ферментами (например, избыточное количество глутамата окисляется глутаматдегидрогеназой).
2.ОБЕСПЕЧИВАЮТ ПРОТЕКАНИЕ РЕАКЦИЙ КОСВЕННОГО ДЕЗАМИНИРОВАНИЯ.
3.ОБЕСПЕЧИВАЮТ СИНТЕЗ МОЧЕВИНЫ
КОСВЕННОЕ ДЕЗАМИНИРОВАНИЕ (ТРАНСДЕЗАМИНИРОВАНИЕ).
Оксидазы аминокислот в клетках организма человека имеют очень низкую активность. Поэтому прямое окислительное дезаминирование аминокислот почти не протекает. Исключением является никотинамидный фермент глутаматдегидрогеназа, который легко дезаминирует глутаминовую кислоту, расщепляя ее до NH3 и альфа-кетоглутарата. Альфа-кетоглутарат может легко вступать в реакции трансаминирования с другими аминокислотами, превращаясь обратно в глутаминовую кислоту.
Значит, если аминокислота не может дезаминироваться прямо, то она может дезаминироваться косвенно с участием пары "альфа-кетоглутарат/глутамат".
Некоторые аминокислоты не имеют "своей собственной" трансаминазы, которая могла бы перенести их аминогруппу на альфа-кетоглутарат (лизин, треонин, пролин, оксипролин), но для этих аминокислот есть специальные реакции, в результате которых аминогруппа этих аминокислот оказывается в составе глутаминовой кислоты.
Таким образом, глутаминовая кислота работает в паре с альфа-кетоглутаровой: альфа-кетоглутарат принимает у разных аминокислот азот в форме аминогруппы, превращаясь в глутамат; глутамат легко дезаминируется прямым путем высокоактивной глутаматдегидрогеназой с образованием аммиака.
Общий итог косвенного дезаминирования такой же, как и результат прямого окислительного дезаминирования.
Это вариант дезаминирования, который протекает в две стадии:
а) трансаминирование с участием альфа-кетоглутаровой кислоты;
б) дезаминирование образовавшейся на первой стадии глутаминовой кислоты.
Косвенное дезаминирование является наиболее важным, т.к. именно таким путем дезаминируются большинство аминокислот живого организма. Это обусловлено тем, что фермент глутамат-ДГ более активен, чем оксидазы аминокислот.
ПРЕИМУЩЕСТВА КОСВЕННОГО ДЕЗАМИНИРОВАНИЯ ПЕРЕД ПРЯМЫМ:
1. В этом процессе не образуется Н2О2, в отличие от прямого дезаминирования, а образуется Н2О и 3 молекулы АТФ (энергия).
2. Косвенное дезаминирование обеспечивает гораздо более быстрое дезаминирование (благодаря высокой активности фермента глутамат-ДГ и трансаминазы);
3. Косвенное дезаминирование протекает не только быстрее, но скорость этого процесса может регулироваться. Возможность регуляции косвенного дезаминирования обусловлена обратимостью процесса и автономной саморегуляцией. Аллостерическими регуляторами глутаматдегидрогеназы являются АДФ (сильный активатор) и АТФ (ингибитор). Ингибирующее действие на этот фермент оказывают также ГТФ и НАДН2.
Если АТФ начинает расходоваться больше, чем образуется в процессе биоокисления в митохондриях, то после активации глутамат-ДГ происходит смещение реакций в сторону образования АТФ, благодаря тому, что глутамат начинает быстрее дезаминироваться, а образующиеся кетокислоты, либо сразу, либо после определенных превращений, могут также вступать на путь митохондриального окисления. Это наблюдается при недостатке жирных кислот или углеводов. То есть при недостатке жиров и углеводов в пище, в качестве дополнительного источника энергии организм начинает использовать аминокислоты. Нередко приходится при голодании разрушать собственные белки. Т.е. при похудании в организме разрушаются не только жиры и углеводы, но и собственные белки. Присутствие достаточного количества жиров и углеводов в пище позволяет сберечь белки от нерационального использования.
СИНТЕЗ ЗАМЕНИМЫХ АМИНОКИСЛОТ В ОРГАНИЗМЕ
Иногда в организме возникает ситуация, когда потребность в тех или иных аминокислотах больше, чем их поступление с пищей. Допустим, организму требуется 7 граммов аланина и 15 граммов глутамата, а с пищей поступило всего лишь 5 граммов аланина, но зато 20 граммов глутамата. Тогда организм, используя механизм трансаминирования, может синтезировать недостающее количество аланина из ПВК, который синтезируется в бих количествах (сотни граммов в сутки). Таким образом, в организме постоянно происходит корректировка аминокислотного состава, но это не уменьшает потребности организма в пищевом белковом азоте. Эта корректировка осуществляется с использование соответствующих альфа-кетокислот, образующихся в ходе метаболизма углеводов и жиров.
Незаменимые аминокислоты не могут образуются в организме, потому что в ходе распада углеводов и жиров не образуется соответствующих им альфа-кетокислот.
Подводя итог, можно сказать, что в результате общих путей катаболизма аминокислот, они разрушаются до СО2, NH3 и безазотистых фрагментов, которые также могут разрушаться до СО2 и Н2О. Аммиак подвергается реакциям обезвреживания.
Трансаминазы обычно катализируют реакции двойного замещения ( реакции типа пинг-понг; разд. В таких реакциях аминогруппа переносится сначала с первого субстрата-аминокислоты-на кофермент, а затем происходит отделение образовавшейся ос-кетокислоты от фермента; после этого с ферментом связывается второй субстрат-вступающая в реакцию а-кетокислота. При этом аминогруппа переносится с пиридокс-аминфосфата на второй субстрат. [2]
Трансаминазы - классический пример ферментов, катализирующих бимолекулярные реакции, протекающие по механизму типа пинг-понг ( разд. В таких реакциях первый субстрат должен уйти из активного центра фермента до того, как второй субстрат сможет к нему присоединиться. Сначала с активным центром фермента связывается приходящая аминокислота, которая отдает свою аминогруппу пиридоксальфосфату и в форме ос-кетокислоты покидает активный центр. Затем с активным центром связывается приходящая ос-кетокислота; она принимает аминогруппу от пиридокс-аминфосфата и отделяется от активного центра, теперь уже в форме аминокислоты. [4]
Трансаминазы в отличие от прочих ферментов, содержащих витамин В6, характеризуются тем, что роль кофермента может выполнять как пиридоксаминфосфат, ISK и пиридоксальфосфат, тогда как в реакциях декарбоксилирования, рацемизации и в других реакциях, катализируемых В6 - ферментами, витамин В6 проявляет каталитическое действие только в форме пиридоксаль-фосфата. [5]
Трансаминазы содержатся как в митохондриях, так и в растворимой фракции ци-топлазмььэукариотических клеток, причем митохондриальные и цитоплазматические формы ферментов различаются по физико-химическим свойствам. [7]
Трансаминазы ( аминотрансферазы) - ферменты, катализирующие перенос аминогруппы от а-аминокислот к а-кетокислотам. [8]
Все трансаминазы ( как и декарбоксилазы аминокислот) содержат один и тот же кофермент-пиридоксальфосфат. Для реакций трансаминирования характерен общий механизм. Специфичность трансаминаз обеспечивается белковым компонентом. Ферменты трансаминирования катализируют перенос NH2 - rpynnbi не на а-кетокислоту, а сначала на кофермент пиридоксальфосфат. [9]
Аминотрансферазы или трансаминазы катализируют межмолекулярный перенос аминогруппы с аминокислот на кетокислоты. Коферментом трансаминаз является фосфопиродоксаль. Он служит непосредственным переносчиком аминогруппы с аминокислоты на кетокислоту. [10]
Кроме этой трансаминазы , специфически приспособленной к L-глутаминовой кислоте, встречается также, главным образом в растительном мире, специфическая трансаминаза для системы L-аспарагиновая кислота - оксалилуксусная кислота. [11]
АМИНОТРАНСФЕРАЗЫ ( трансаминазы ), ферменты класса трансфераз; катализируют окислительно-восстановит. Определение активности алани-новой и аспарагиновой А. [12]
В реакции участвует фермент трансаминаза с пиридоксинфос-фатом ( В ]) в качестве кофермента. В реакции переаминирования глутаминовая кислота служит донором аминогруппы. [13]
Пиридоксальфосфат входит в качестве простетической группы в ферменты трансаминазы , участвующие в обратимом переносе аминогрупп ( переаминировании) белковых аминокислот. Дисульфидное производное пиридоксина - пиридитол ( 55) - является психотропным агентом антидепрессантного действия с седативным компонентом. Он проявляет ноотропный эффект и уменьшает головные боли. Назначают пиридитол при неглубоких депрессиях, мигрени, церебральном атеросклерозе, а у детей - при задержке психического развития и олигофрении. Уместно отметить, что но-отропным и анксиолитическим ( снимающим беспокойство) эффектом обладает мексидол ( 56), который также имеет гид-роксигруппу при С-3. В основном он используется в виде сук-цината как антиоксидант и антигипоксант. Он также способствует рассасыванию внутриглазных кровоизлияний. [14]
К ним относятся ферменты обмена аминокислот: ами-нотрансферазы ( трансаминазы ), катализирующие реакции переаминирования, а также ампдипотрансфора-зы, осуществляющие перенос ампдшювой группировки. [15]
Реакции трансаминирования являются обратимыми и, как выяснилось позже, универсальными для всех живых организмов. Эти реакции протекают при участии специфических ферментов, названных А.Е. Браун-штейном аминоферазами (по современной классификации, аминотранс-феразы, или трансаминазы). Теоретически реакции трансаминиро-вания возможны между любой амино- и кетокислотой, однако наиболее интенсивно они протекают в том случае, когда один из партнеров представлен дикарбоновой амино- или кетокислотой. В тканях животных и у микроорганизмов доказано существование реакций трансаминирования между монокарбоновыми амино- и кетокислотами. Донорами NН2-группы могут также служить не только α-, но и β-, γ- и ω-аминогруппы ряда аминокислот. В лаборатории А. Майстера доказано, кроме того, трансами-нирование глутамина и аспарагина с кетокислотами в тканях животных.
В переносе аминогруппы активное участие принимает кофермент транс-аминаз пиридоксальфосфат (производное витамина В6; см. главу 5), который в процессе реакции обратимо превращается в пиридоксаминфосфат.
Механизм реакции трансаминирования. Общую теорию механизма ферментативного трансаминирования разработали советские ученые А.Е. Браун-штейн и М.М. Шемякин. Одновременно подобный механизм был предложен американскими биохимиками Э. Снеллом и Д. Метцлером. Все трансаминазы (как и декарбоксилазы аминокислот) содержат один и тот же кофермент – пиридоксальфосфат. Для реакций трансаминирования харак -терен общий механизм. Специфичность трансаминаз обеспечивается белковым компонентом. Ферменты трансаминирования катализируют перенос NH2-группы не на α-кетокислоту, а сначала на кофермент пиридоксаль-фосфат. Образовавшееся промежуточное соединение (шиффово основание) подвергается внутримолекулярным превращениям (лабилизация α-водо-родного атома, перераспределение энергии связи), приводящим к освобождению α-кетокислоты и пиридоксаминфосфата; последний на второй стадии реакции реагирует с любой другой α-кетокислотой, что через те же стадии образования промежуточных соединений (идущих в обратном направлении) приводит к синтезу новой аминокислоты и освобождению пиридоксальфосфата. Опуская промежуточные стадии образования шиффовых оснований, обе стадии реакции трансаминирования можно представить в виде общей схемы:
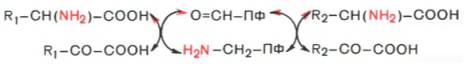
Более подробно механизм действия трансаминаз представлен на рис. 12.3.
В связи с тем что во всех пиридоксалевых ферментах (включая транс-аминазы) карбонильная группа кофермента (—СНО) оказалась связанной с ε-аминогруппой лизина белковой части, в классический механизм реакции трансаминирования А.Е. Браунштейн и Э. Снелл внесли следующее дополнение. Оказалось, что взаимодействие между субстратом, т.е. L-амино-кислотой (на рисунке – аспартат), и пиридоксальфосфатом происходит не путем конденсации с выделением молекулы воды, а путем реакции замещения, при которой NH2-группа субстрата вытесняет ε-NН2-группу лизина в молекуле ферментного белка, что приводит к формированию пиридоксальфосфатного комплекса.
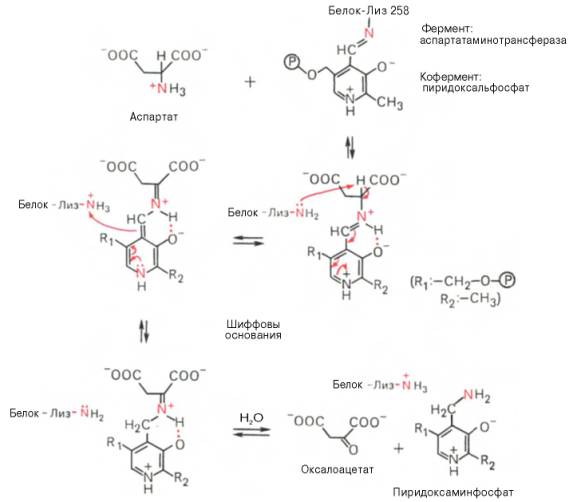
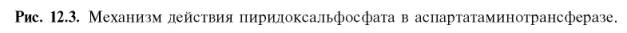
Существование представленного механизма реакции трансаминирова-ния доказано разнообразными методами, включая методы спектрального анализа по идентификации промежуточных альдиминных и кетиминных производных пиридоксальфосфата.
Роль трансаминаз и реакций трансаминирования в обмене аминокислот.
Чрезвычайно широкое распространение трансаминаз в животных тканях, у микроорганизмов и растений, их высокая резистентность к физическим, химическим и биологическим воздействиям, абсолютная стереохимическая специфичность по отношению к L-аминокислотам, а также высокая каталитическая активность в процессах трансаминирования послужили предметом детального исследования роли этих ферментов в обмене аминокислот. Ранее было указано, что при физиологических значениях рН среды активность оксидазы L-аминокислот резко снижена. Учитывая это обстоятельство, а также высокую скорость протекания реакции трансами-нирования, А.Е. Браунштейн выдвинул гипотезу о возможности существования в животных тканях непрямого пути дезаминирования аминокислот через реакции трансаминирования, названного им трансдезаминированием. Основой для выдвижения этой гипотезы послужили также данные Г. Эйлера о том, что в животных тканях из всех природных аминокислот с высокой скоростью дезаминируется только L-глутаминовая кислота в реакции, катализируемой высокоактивной и специфической глутамат-дегидрогеназой.
Согласно гипотезе, получившей экспериментальное подтверждение, все или почти все природные аминокислоты (исключение составляет метионин) сначала реагируют с α-кетоглутаровой кислотой в реакции трансами-нирования с образованием глутаминовой кислоты и соответствующей кетокислоты. Образовавшаяся глутаминовая кислота затем подвергается непосредственному окислительному дезаминированию под действием глу-таматдегидрогеназы. Схематически механизм трансдезаминирования можно представить в следующем виде:
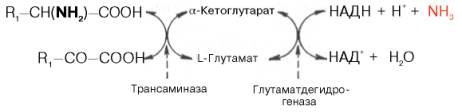
Суммарная реакция при этом следующая:
Поскольку обе реакции (трансаминирование и дезаминирование глу-таминовой кислоты) являются обратимыми, создаются условия для синтеза по существу любой аминокислоты, если в организме имеются соответствующие α-кетокислоты. Известно, что организм животных и человека не наделен способностью синтеза углеродных скелетов (α-кетокислот), так называемых незаменимых аминокислот; этой способностью обладают только растения и многие микроорганизмы.
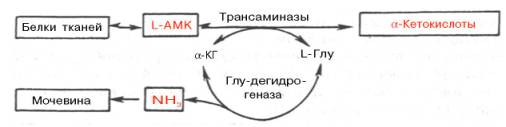
Рис. 12.4. Центральная роль трансаминаз L-аминокислот и глутаматдегидрогеназы в биосинтезе и распаде аминокислот в тканях животных. АМК - аминокислоты; α-КГ - α-кетоглутарат.
Механизм, при помощи которого в живых организмах осуществляется синтез природных аминокислот из α-кетокислот и аммиака, был назван А.Е. Браунштейном трансреаминированием. Сущность его сводится к восстановительному аминированию α-кетоглутаровой кислоты с образованием глутаминовой кислоты (реакцию катализирует НАДФ-зависимая глута-матдегидрогеназа, работающая в режиме синтеза) и к последующему трансаминированию глутамата с любой α-кетокислотой. В результате образуется L-аминокислота, соответствующая исходной кетокислоте, и вновь освобождается α-кетоглутаровая кислота, которая может акцептировать новую молекулу аммиака. Роль реакций трансаминирования как в дезаминировании, так и в биосинтезе аминокислот может быть представлена в виде схемы:
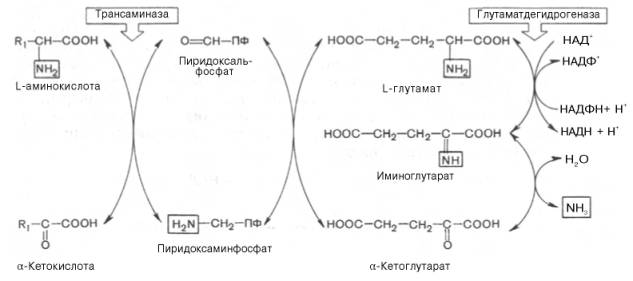
Таким образом, трансаминазы катализируют опосредованное через глутаматдегидрогеназу дезаминирование природных аминокислот (черные стрелки) и биосинтез аминокислот (красные стрелки). В более упрощенной форме роль этих ключевых ферментов азотистого обмена представлена на рис. 12.4.
Получены доказательства существования в организме теплокровных животных еще одного механизма непрямого (опосредованного) дезами-нирования L-аминокислот, при котором Глу, Асп и АМФ выполняют роль системы переноса NН2-группы; гидролитическое дезаминирование АМФ приводит к образованию инозинмонофосфата (ИМФ) и аммиака:
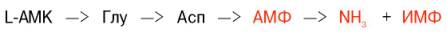
Возможно, что в аналогичной системе в качестве промежуточного переносчика NH2-группы вместо АМФ участвует НАД.
Клиническое значение определения активности трансаминаз. Широкое распространение и высокая активность трансаминаз в органах и тканях человека, а также сравнительно низкие величины активности этих ферментов в крови послужили основанием для определения уровня ряда трансаминаз в сыворотке крови человека при органических и функциональных поражениях разных органов. Для клинических целей наибольшее значение имеют две трансаминазы – аспартат-аминотрансфераза (AcAT) и аланин-аминотрансфераза (АлАТ), катализирующие соответственно следующие обратимые реакции:
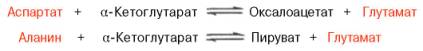
В сыворотке крови здоровых людей активность этих трансаминаз в тысячи раз ниже, чем в паренхиматозных органах. Поэтому органические поражения при острых и хронических заболеваниях, сопровождающиеся деструкцией клеток, приводят к выходу трансаминаз из очага поражения в кровь. Так, уже через 3–5 ч после развития инфаркта миокарда уровень АсАТ в сыворотке крови резко повышается (в 20–30 раз). Максимум активности обеих трансаминаз крови приходится на конец первых суток, а уже через 2–3 дня при благоприятном исходе болезни уровень сывороточных трансаминаз возвращается к норме. Напротив, при затяжном процессе или наступлении повторного инфаркта миокарда наблюдается новый пик повышения активности этих ферментов в крови. Этим объясняется тот факт, что в клинике трансаминазный тест используется не только для постановки диагноза, но и для прогноза и проверки эффективности лечения . При поражениях клеток печени, например при гепатитах, также наблюдается гипертрансаминаземия (за счет преимущественного повышения уровня АлАТ), но она имеет более умеренный и затяжной характер, а повышение активности трансаминазы в сыворотке крови происходит медленно. При различного рода коронарной недостаточности (стенокардия, пороки сердца и др., кроме инфаркта миокарда) гипертрансаминаземия или не наблюдается, или незначительна. Определение активности трансаминаз в сыворотке крови при заболеваниях сердца следует отнести к дифференциально-диагностическим лабораторным тестам. Повышение уровня трансаминаз в сыворотке крови отмечено, кроме того, при некоторых заболеваниях мышц, в частности при обширных травмах, гангрене конечностей и прогрессивной мышечной дистрофии.
Превращения α-кетокислот. Образовавшиеся в процессе дезаминиро-вания и трансдезаминирования α-кетокислоты подвергаются в тканях животных различным превращениям и могут вновь трансаминироваться с образованием соответствующей аминокислоты. Это так называемый синтетический путь превращения. Опыты с перфузией растворов α-кето-кислот и аммиака через изолированную печень показали, что в оттекающей из печени жидкости действительно имеются соответствующие исходным кетокислотам L-аминокислоты. Открыты, кроме того, гликогенные, кето-генные и окислительные пути, ведущие к образованию соответственно глюкозы, жирных кислот, кетоновых тел и компонентов цикла трикарбоновых кислот (ЦТК). Эти процессы можно представить в виде общей сводной схемы:
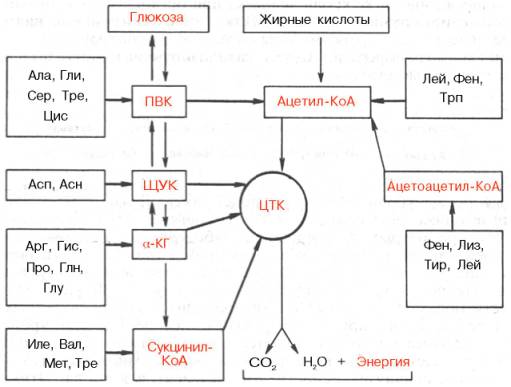
![]()
Аминотрансфера́зы (трансаминазы) — ферменты из группы трансфераз, переносящие аминогруппы без образования свободного аммиака. Аминотрансферазы также называют трансаминазами, а реакцию — трансаминированием. Ферменты относятся к подклассу КФ 2.6.
Для аминотрансфераз донором аминогрупп являются аминокислоты, а акцептором — кетокислоты:
AK1 + KK2 ↔ KK1 + AK2

В составе простетической группы аминотрансферазы содержат производные витамина B6. Во время переноса аминогруппы простетическая группа переходит из пиридоксаль-5-фосфатной формы в пиридосамино-5-фосфатную форму.
Механизм реакции трансаминирования открыт в 1937 году советскими учеными А.Е. Браунштейном и М.Г.Крицман.
Процесс протекает в две стадии.
-
пиридоксальфосфата (-СНО) взаимодействует с аминогруппой аминокислоты с образованием иминной связи в основании Шиффа: сначала α-аминогруппа аминокислоты-донора замещает ε-аминогруппу апофермента, а затем происходит перегруппировка через кетимин и в результате гидролиза образуется пиридосамино-5-фосфат и α-кетокислота.
- Реакции повторяются в обратном порядке
Аминотрансферазы содержатся практически во всех органах, но наиболее активно реакции трансаминирования идут в печени.
К этой группе ферментов относятся такие важные для клинической лабораторной диагностики ферменты, как АСТ и АЛТ.
Литература
- Комов В.П. Биохимия: Учеб. для вузов / В.П.Комов, В.Н.Шведова. - М.:Дрофа, 2004. - 640 с.: ил. - (Высшее образование: Современный учебник)
Wikimedia Foundation . 2010 .
Полезное
Смотреть что такое "Аминотрансферазы" в других словарях:
АМИНОТРАНСФЕРАЗЫ — (трансаминазы) ферменты класса трансфераз, обратимо катализирующие перенос аминогрупп от аминокислот на кетокислоты. Катализируемые аминотрансферазами реакции трансаминирования осуществляют связь между белковым и углеводным обменом в живых… … Большой Энциклопедический словарь
АМИНОТРАНСФЕРАЗЫ — трансаминазы, ферменты класса трансфераз, катализирующие обратимые реакции трансаминирования, т. е. перенос аминогруппы ( NH2) и водорода от одной молекулы субстрата к другой без промежуточного образования аммиака. Кофер менты А.… … Биологический энциклопедический словарь
Аминотрансферазы — I Аминотрансферазы (синоним трансаминазы) ферменты, катализирующие реакцию переноса аминогруппы (NH2 группы) вместе с протоном (ионом водорода) и парой электронов от аминокислот или аминов к кетокислотам или другим соединениям, содержащим в… … Медицинская энциклопедия
аминотрансферазы — (трансаминазы), ферменты класса трансфераз, обратимо катализирующие перенос аминогрупп от аминокислот на кетокислоты. Катализируемые аминотрансферазы реакции трансаминирования осуществляют связь между белковым и углеводным обменом в живых… … Энциклопедический словарь
аминотрансферазы — (син. трансаминазы) ферменты класса трансфераз (КФ 2.6.1), катализирующие обратимый перенос аминогруппы с аминокислот на кетокислоты; повышение активности А. в сыворотке крови наблюдается при патологических состояниях, сопровождающихся… … Большой медицинский словарь
Аминотрансферазы — аминоферазы, трансаминазы, ферменты из группы трансфераз (См. Трансферазы), катализируют перенос аминогрупп ( NH2) от α аминокислот на α кетокислоты. А. обнаружены в большинстве тканей животных и растений, играют важную роль в азотистом… … Большая советская энциклопедия
АМИНОТРАНСФЕРАЗЫ — (трансаминазы), ферменты кл. трансфераз, обратимо катализирующие перенос аминогрупп от аминокислот на кетокислоты. Катализируемые А. реакции трансаминирования осуществляют связь между белковым и углеводным обменом в живых организмах … Естествознание. Энциклопедический словарь
АМИНОТРАНСФЕРАЗЫ — ферменты из класса трансфераз, катализируют перенос аминогрупп (NH2) между аминокислотами и кетокислотами без промежуточного образования аммиака. Коферменты А. производные витамина B6 пиридоксальфосфат и пиридоксаминфосфат. Наибольшее клинич.… … Ветеринарный энциклопедический словарь
Аминотрансферазы трансаминазы — Аминотрансферазы, трансаминазы * амінатрансферазы, трансаміназы * aminotransferases or transaminases ферменты, катализирующие перенос αаминогрупп к. л. аминокислоты на αуглеродный атом какой либо αкетокислоты. Результатом реакции является… … Генетика. Энциклопедический словарь
Трансаминирование аминокислот. Структура кофермента и механизм переаминирования. Специфичность трансаминаз. Значение реакций трансаминирования. Диагностическое значение определения в сыворотке крови органоспецифичных аминотрансфераз АЛТ и АСТ. Коэффициент де Ритиса.
Трансаминирование
Трансаминирование - реакция переноса α-аминогруппы с аминокислоты на α-кетокислоту, в результате чего образуются новая кетокислота и новая аминокислота. Константа равновесия для большинства таких реакций близка к единице (Кр~1,0), поэтому процесс трансаминирования легко обратим (см. схему А).
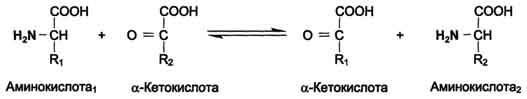
Реакции катализируют ферменты аминотрансферазы, коферментом которых служит пиридоксальфосфат (ПФ) - производное витамина В6 (пиридоксина, см. раздел 3) (см. схему Б).
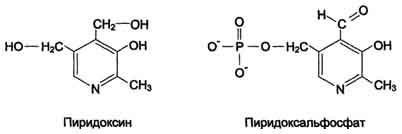
Аминотрансферазы обнаружены как в цитоплазме, так и в митохондриях клеток эукариот. Причём митохондриальные и цитоплазматические формы ферментов различаются по физико-химическим свойствам. В клетках человека найдено более 10 аминотрансфераз, отличающихся по субстратной специфичности. Вступать в реакции трансаминирования могут почти все аминокислоты за исключением лизина, треонина и пролина.
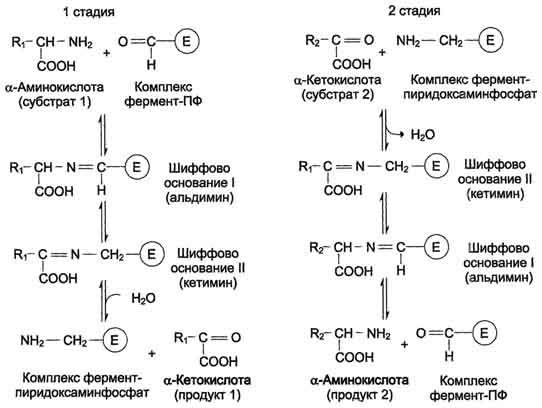
Механизм реакции
Аминотрансферазы - классический пример ферментов, катализирующих реакции, протекающие по механизму типа "пинг-понг" (см. раздел 2). В таких реакциях первый продукт должен уйти из активного центра фермента до того, как второй субстрат сможет к нему присоединиться.
Активная форма аминотрансфераз образуется в результате присоединения пиридоксальфосфата к аминогруппе лизина прочной альдиминной связью (рис. 9-6). Лизин в положении 258 входит в состав активного центра фермента. Кроме того, между ферментом и пиридоксальфосфатом образуются ионные связи с участием заряженных атомов фосфатного остатка и азота в пиридиновом кольце кофермента.
Пиридоксальфосфат в данном случае служит переносчиком аминогрупп. При этом наиболее важную роль играет его альдегидная группа, которая может обратимо присоединять различные амины с образованием шиффовых оснований. Реакции трансаминирования проходят в 2 стадии, во время которых пиридрксальфосфат претерпевает обратимые превращения между свободной альдегидной формой (ПФ) и аминированной формой (пиридоксаминфосфат).
Последовательность реакций трансаминирования представлена ниже.
На первой стадии к пиридоксальфосфату в активном центре фермента с помощью альдиминной связи присоединяется аминогруппа от первого субстрата - аминокислоты. Образуются комплекс фермент-пиридокса-минфосфат и кетокислота - первый продукт реакции. Этот процесс включает промежуточное образование 2 шиффовых оснований.
На второй стадии комплекс фермент-пиридоксаминфосфат соединяется с кетокислотой (вторым субстратом) и снова через промежуточное образование 2 шиффовых оснований передаёт аминогруппу на кетокислоту. В результате фермент возвращается в свою нативную форму, и образуется новая аминокислота - второй продукт реакции. Если альдегидная группа пиридоксальфосфата не занята аминогруппой субстрата, то она образует шиффово основание (альдимин) с ε-аминогруппой радикала лизина в активном центре фермента (см. схему на с. 471).
Органоспецифичные аминотрансферазы АНТ и ACT
Чаще всего в реакциях трансаминирования участвуют аминокислоты, содержание которых в тканях значительно выше остальных - глутамат, аланин, аспартат и соответствующие им кетокислоты - α-кетоглутарат, пируват и оксалоацетат. Основным донором аминогруппы служит глутамат.
Суммарно эти реакции можно представить в виде схемы:

Акцептором аминогруппы любой аминокислоты, подвергающейся трансаминированию (аминокислота 1), служит α-кетоглутарат. Принимая аминогруппу, он превращается в глутамат, который способен передавать эту группу любой α-кетокислоте с образованием другой аминокислоты (аминокислота 2).
Аминотрансферазы обладают субстратной специфичностью к разным аминокислотам. В тканях человека обнаружено более 10 разных аминотрансфераз.
Наиболее распространёнными ферментами в большинстве тканей млекопитающих являютсяаланинаминотрансфераза (АЛТ), по обратной реакции - глутамат-пируватаминотрансфераза(ГПТ) и аспартатаминотрансфераза (ACT), по обратной реакции - глутамат-оксалоацетатаминотрансфераза (ГОТ).
АЛТ (АлАТ) катализирует реакцию транса-минирования между аланином и α-кетоглутаратом (см. схему А на с. 472).
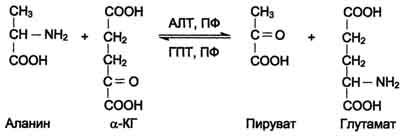
Локализован этот фермент в цитозоле клеток многих органов, но наибольшее его количество обнаружено в клетках печени и сердечной мышцы.
ACT (АсАТ) катализирует реакцию трансами-нирования между аепартатом и α-кетоглутаратом аналогично предыдущей (см. схему Б на с. 472).
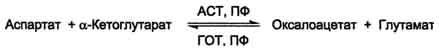
В результате образуются оксалоацетат и глутамат. ACT имеет как цитоплазматическую, так и митохондриальную формы. Наибольшее его количество обнаружено в клетках сердечной мышцы и печени.
Так как наибольшее количество АЛТ и ACT сосредоточено в печени и миокарде, а содержание в крови очень низкое, можно говорить об органоспецифичности этих ферментов.
В результате работы аминотрансфераз аминный азот многих аминокислот переходит в состав глутамата. Есть основания считать, что накопление аминогрупп в форме глутаминовой кислоты происходит в цитозоле. Затем глутамат с помощью транслоказ попадает в митохондрии, где активна специфическая ACT. В результате действия этого фермента глутамат снова превращается в α-кетоглутарат. Последний используется для непрямого дезаминирования аминокислот, содержащихся в митохондриях. Это очень важно, так как только глутамат в тканях млекопитающих наиболее быстро может подвергаться окислительному дезаминированию (см. ниже).
Читайте также: