
Гетерогенность модулирующей системы мозга кратко
Обновлено: 05.07.2024
ЭКГ, ЭМГ и др.), которые обычно используются для оценки ФС, все они в одинаковой мере связаны с активацией неспецифической системы мозга и что по динамике любой из них можно предсказать изменение всех остальных реакций. Такой подход основывался на появившейся в 1951 г. концепции Д. Линдсли о единстве и синергизме восходящих и нисходящих влияний от неспецифической системы мозга, обусловливающих параллелизм всех ЭЭГ-х, вегетативных и моторных реакций активации.
Однако позже были получены данные о низких коэффициентах корреляции между различными показателями активации, а также о диссоциации ЭЭГ и поведенческого пробуждения. Введение собаке атропина вызывает ЭЭГ медленного сна, в то время как животное поведенчески продолжает бодрствовать. С помощью физосигмина можно вызвать активированную ЭЭГ, тогда как животное будет находиться в состоянии дремоты.
381
Сон
Длительное время господствовало представление о том, что во время сна, когда в течение нескольких часов в организме идут процессы восстановления, наблюдается общее снижение активности тела и мозга.
386
Такое понимание соответствовало особенностям сонного поведения, когда человек или животное, приняв удобную позу, многие часы находились без движения, а также регистрируемой при этом ЭЭГ, в которой появлялись медленные волны.
Позже представление о падении активности мозга во время сна было отвергнуто прежде всего благодаря открытию особой стадии сна, так называемого парадоксального или быстрого сна. Его открытие в 1953 г. связано с именами аспиранта Э. Азеринского и исследователя Н. Клейтмена, которые во время ночного сна у здоровых людей зарегистрировали быстрые движения глаз. Периоды таких движений в течение ночи появлялись 4-5 раз и, как в дальнейшем было показано У. Дементом и Н. Клейтменом, они совпадали с десинхронизацией в ЭЭГ. По данным М. Жуве, десинхронизация ЭЭГ во время сна сочетается с фазическими двигательными реакциями в вице подергивания конечностей, вибрис, хвоста.
Таким образом, сон — это не просто углубление и распространение торможения. Это сложный процесс, имеющий свои стадии, каждая из которых сама по себе уникальна. Различие между ними не может быть описано количественно. При этом мозг во время сна характеризуется высоким уровнем активности и в некотором смысле даже более высоким, чем при спокойном бодрствовании (рис. 46).
387
393
Естественный сон характеризуется циклической сменой медленного и парадоксального сна. Полный цикл, состоящий из смены медленного сна на быстрый с последующим возвращением к медленному, у человека занимает 60-90 мин. На ночной сон приходится 4-5 полных цикла. Сон всегда начинается с медленного сна (рис. 48). У нормального человека на быстрый сон в среднем приходится около 20% от общей продолжительности сна. Длительность фазы быстрого сна к утру увеличивается,.а медленного уменьшается, так что до 72% медленного сна расходуется в течение первой половины ночи. Фазические компоненты ПС более выражены в последних циклах сна.
Структура сна у большинства млекопитающих сходна. Различия касаются в основном числа циклов смены MC на ПС. Так, у кошки в течение 5-часового периода развивается 10 циклов, тогда как у крысы количество циклов за это же время достигает 22. Такое
Рис. 48. Типичное чередование фаз сна у взрослого молодого человека.
Рис. 48. Типичное чередование фаз сна у взрослого молодого человека. Д — бодрствование, MC — медленный сон. Черной линией показаны периоды парадоксального сна (ПС) с характерной ЭЭГ-десинхронизацией и быстрыми движениями глаз (БДГ) (по Р. Томпсону, 1974)

394
408
В 1936 г., описав впервые ОАС, или синдром биологического стресса, Г.Селье выделяет его три стадии: стадию тревоги (I), стадию резистентности (II) и стадию истощения (III).
II стадия стресса — стадия сопротивления. Если действие совместимо с возможностями адаптации, то в организме стабилизируется фаза сопротивления. При этом признаки тревоги практически исчезают, а уровень сопротивляемости поднимается значительно выше обычного.
Рис. 50. Основные пути действия стрессора — триада стресса (по Г. Селье, 1982)

409
411
Рис. 51. Схема развития стресс-реакции (по Г. Л. Кассилю, 1975)

418
Ранее предполагалось, что по динамике любой из физиологических реакций (ЭЭГ, ЭМГ, диаметр зрачка, кожное сопротивление, ЧСС), испытывающих влияние неспецифической системы, можно предсказать изменение всех остальных реакций. Такой подход основывался на концепции Д. Линдсли о единстве и синергизме влияний от восходящих и нисходящих неспецифических систем мозга, обусловливающих параллелизм всех электроэнцефалографических, вегетативных и двигательных реакций. Предполагалось, что активация в ЭЭГ (проявление действия восходящей активирующей неспецифической системы) развивается параллельно эффектам нисходящей неспецифической системы в виде соматических и вегетативных реакций (изменения тонуса мышц, движения глаз, изменения частоты дыхания и сердцебиения, КГР, изменения диаметра сосудов). Однако в дальнейшеммежду ними были найдены очень низкие коэффициенты корреляции. На основе обзора литературных данных Р. Jla3apyc приходит к выводу, что максимальные значения коэффициентов корреляции не превышали 0,5.
Кроме того, при определенных условиях была обнаружена диссоциация между поведенческой и электроэнцефалографической активациями. Так, введение атропина собаке вызывало ЭЭГ-картину медленного сна, при этом животное поведенчески продолжало бодрствовать. Аналогичные данные были получены и на кошках. С помощью физостигмина у них можно было вызвать активацию на ЭЭГ, но поведенчески животное могло находиться в состоянии дремоты. Дж. Моруцци показал, что если перерезать ствол мозга выше варолиевого моста, но ниже ретикулярной формации, то в результате прерывания синхронизирующих влияний, поступающих из него в кору, в ЭЭГ развивается десинхронизация. В это время животное может спать.
ЭКГ, ЭМГ и др.), которые обычно используются для оценки ФС, все они в одинаковой мере связаны с активацией неспецифической системы мозга и что по динамике любой из них можно предсказать изменение всех остальных реакций. Такой подход основывался на появившейся в 1951 г. концепции Д. Линдсли о единстве и синергизме восходящих и нисходящих влияний от неспецифической системы мозга, обусловливающих параллелизм всех ЭЭГ-х, вегетативных и моторных реакций активации.
Однако позже были получены данные о низких коэффициентах корреляции между различными показателями активации, а также о диссоциации ЭЭГ и поведенческого пробуждения. Введение собаке атропина вызывает ЭЭГ медленного сна, в то время как животное поведенчески продолжает бодрствовать. С помощью физосигмина можно вызвать активированную ЭЭГ, тогда как животное будет находиться в состоянии дремоты.
активации, а несколько субсистем, выражением которых являются вегетативные, моторные и ЭЭГреакции.
продемонстрировал возможность избирательной блокады каждого из этих состояний.
Известна точка зрения A.A. Роутенберга, выделяющего две системы активации: лимбическую систему и ретикулярную формацию ствола мозга с их относительным антагонизмом. Активирующая ретикулярная формация обеспечивает энергетическую базу исполнения реакций и имеет тенденцию участвовать в оборонительном поведении, тогда как лимбическая система занята преимущественно активацией вегетативных процессов и связана с подготовительными фазами поведения.
С помощью факторного анализа ЭЭГ затылка и лба В.М. Русаловым выделено четыре общемозговых интегральных ЭЭГ-фактора: 1) фактор энергии медленных волн; 2) фактор частоты медленных ритмов; 3) фактор активности бета-2; 4) фактор пространственно-временной сопряженности ЭЭГ. Факторы рассматриваются как отражение независимых аспектов регуляции уровня неспецифической активации, за которыми стоит функционирование раздельных и относительно самостоятельных систем активации мозга.
В свете этих представлений функциональное состояние является результатом взаимодействия или баланса отдельных субсистем активации, т. е. ФС — явление системное, которое требует системного подхода.
Серьезное подтверждение существования субсистем активации, взаимодействие которых определяет функциональное состояние человека, получено в психофизических экспериментах.
Две системы субъективной активации методом семантического дифференциала выделены В.И. Викторовым. Показано их различное взаимоотношение с успешностью обучения в вузе. Фактор А, охватывающий континуум состояний от спокойствия до волевого усилия, связан с успешностью обучения куполообразной зависимостью, тогда как фактор Р, соответствующий шкале переживаний от печали до радости, и успешность обучения связаны отрицательной линейной зависимостью.
Ч. Осгуда интерпретированы как оценка (1), сила (потенция) (2) и активность (3), где фактор активности соответствует шкале эмоциональности, фактор силы — шкале бодрствования, по Р. Тейеру, а фактор оценки — шкале приятных и неприятных переживаний. Существенной особенностью пространства аффективных значений Ч. Осгуда является его универсальность, т. е.
независимость от выборки обследуемых, различающихся культурой, возрастом, уровнем интеллектуального развития, языком. Это указывает на то, что в пространстве аффективных значений, по-видимому, находят отражение некоторые общие физиологические закономерности работы мозга, имеющие отношение к базальным механизмам регуляции состояний человека.
Подтверждение существования двух субсистем активации: эмоциональной и неэмоциональной —
получено нами при построении семантических пространств состояний методом многомерного шкалирования (по алгоритму Торгерсона). Испытуемые ранжировали степень различия пар слов из списка, который включал как названия эмоций, так и состояний с минимальной эмоциональной компонентой (например, спокойствие, растерянность, уверенность и др.). Семантическое пространство состояний у всех оказалось трехмерным. Его оси ортогональны и интерпретированы как: 1) шкала знаков состояний (положительные — отрицательные состояния); 2) шкала бодрствования или готовности к действию (неэмоциональная активация — НЭА); 3) шкала выраженности эмоционального тона (эмоциональная активация — ЭА).
Индивидуальные семантические пространства состояний различались субъективной значимостью трех основных осей пространства. У лиц с низкой реактивностью (по тесту Я. Стреляу) и высокой экстраверсией (по Г. Айзенку) шкала бодрствования (или неэмоциональной активации) имела более высокий
удельный вес по сравнению со шкалой эмоциональной активации.
Представление о системном характере регуляции ФС получило подтверждение и при изучении
специального класса регуляторов ФС — волновых генераторов электрической активности мозга.
Известно, что модулирующая система распространяет два типа влияний: 1) тонические, когда изменяется средний уровень возбудимости нервной системы, и 2) ритмические, когда возбудимость нервных элементов меняется периодически.
Сейчас накоплены многочисленные данные о том, что поток возбуждения, поступающий в мозг по сенсорным путям, квантуется его ритмической активностью (тета-, альфа-активностью, волнами ЭКГ и др.). Ритмической модуляции подвергаются также и исполнительные, двигательные системы. Время простой сенсомоторной реакции, вероятность появления произвольной двигательной реакции модулируются периодом альфа-волны у человека. Ухудшение и улучшение зрительного восприятия эмоционального слова по показателю КГР связаны с различными фазами альфа-волны. Скорость привыкания также зависит от фазы альфа-волны, с которой совпадает подача повторяющегося стимула. Двигательные инструментальные реакции крысы — нажим на педаль и ее отпускание — соответствуют разным противоположным фазам тета-волн. Высказано предположение, что ритмическая активность мозга осуществляет квантование и в системе памяти. На основе изучения нейронных механизмов ритмической активности мозга у животных и ЭЭГ у человека выделено два самостоятельных волновых генератора: генератор низкочастотного (7-8 Гц) и среднечастотного (9-10 Гц) альфа-ритма. Они были идентифицированы как две субсистемы активации:
эмоциональной и неэмоциональной. Оба генератора обладают резонансными свойствами. Их активность представлена в УВП разными компонентами. Конкретное ФС человека в условиях бодрствования определяется балансом двух систем ритмической активности. При этом чем выше активность среднечастотного альфа-генератора, тем ниже уровень активности низкочастотного генератора, и наоборот [11].
Для среднечастотного альфа-генератора найден ЭЭГ-показатель его активности в виде реакции усвоения ритма световых мельканий, совпадающего с его резонансной частотой. Чем больше доля высоких гармоник в составе реакции усвоения, тем выше уровень активности среднечастотного генератора. Показателем активности низкочастотного альфа-генератора может служить вертекспотенциал. Чем больше его амплитуда, тем выше активность низкочастотного альфа-генератора.
Оптимальность ФС для выполняемой работы связана с преобладанием активности среднечастотного альфа-генератора над низкочастотным. Взаимоотношение этих двух волновых генераторов или двух систем активации и обусловливает колоколообразную форму кривой, которой обычно описывают отношения эффективности деятельности и ФС. Падение эффективности выполняемой работы при высоких уровнях активации нервной системы детерминировано сдвигом баланса двух систем активации в направлении преобладания системы ЭА над НЭА.
Длительное время господствовало представление о том, что во время сна, когда в течение нескольких часов в организме идут процессы восстановления, наблюдается общее снижение активности тела и мозга.
Такое понимание соответствовало особенностям сонного поведения, когда человек или животное, приняв удобную позу, многие часы находились без движения, а также регистрируемой при этом ЭЭГ, в которой появлялись медленные волны.
Позже представление о падении активности мозга во время сна было отвергнуто прежде всего благодаря открытию особой стадии сна, так называемого парадоксального или быстрого сна. Его открытие в 1953 г. связано с именами аспиранта Э. Азеринского и исследователя Н. Клейтмена, которые во время ночного сна у здоровых людей зарегистрировали быстрые движения глаз. Периоды таких движений в течение ночи появлялись 4-5 раз и, как в дальнейшем было показано У. Дементом и Н. Клейтменом, они совпадали с десинхронизацией в ЭЭГ. По данным М. Жуве, десинхронизация ЭЭГ во время сна сочетается с фазическими двигательными реакциями в вице подергивания конечностей, вибрис, хвоста.
Таким образом, сон — это не просто углубление и распространение торможения. Это сложный процесс, имеющий свои стадии, каждая из которых сама по себе уникальна. Различие между ними не может быть описано количественно. При этом мозг во время сна характеризуется высоким уровнем активности и в некотором смысле даже более высоким, чем при спокойном бодрствовании (рис. 46).
высоковольтными дельта-волнами; на IV стадии развивается наиболее глубокий медленный сон: в ЭЭГ дельта-волны без веретен.
Переход от бодрствования ко сну — это особое состояние, которое осуществляется через ряд гипнотических фаз, выделенных и описанных И.П. Павловым
Нейрогуморальная регуляция активности мозга. Нейрогормональные системы мозга человека
а) Нейрогормональная регуляция мозговой активности. Для регуляции мозговой активности помимо проведения специфических нервных сигналов из нижних областей мозга к его корковым регионам очень часто используется другой физиологический механизм. Он связан с секрецией в вещество мозга возбуждающих и тормозных агентов, нейромедиаторов и нейрогормонов. Эти нейрогормоны часто сохраняются в течение нескольких минут или часов, обеспечивая долговременную регуляцию вместо мгновенного действия в виде активации или торможения.
На рисунке ниже показаны три нейрогуморальные системы, детально изученные в мозге крыс:
(1) норадреналиновая система;
(2) дофаминовая система;
(3) серотониновая система.
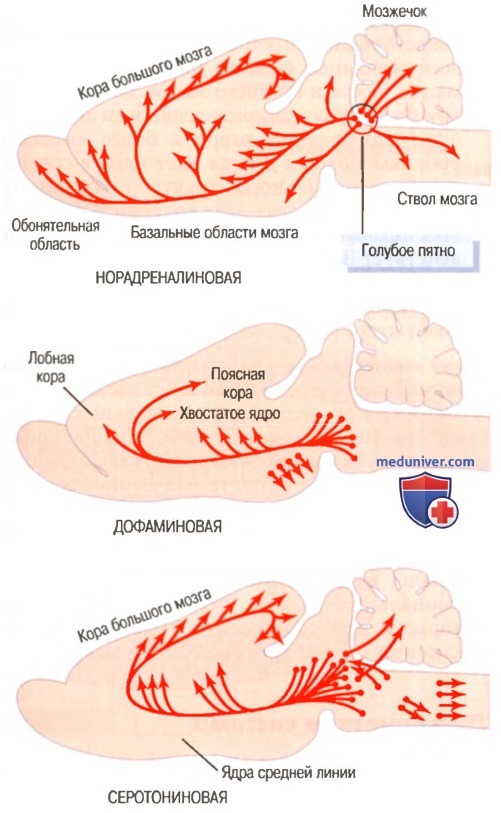
Три нейрогуморальные системы в мозге крысы: норадреналиновая, дофаминовая и серотониновая
Норадреналин обычно функционирует как возбуждающий гормон, тогда как серотонин — как правило, тормозной агент, а дофамин в одних областях является возбуждающим, но в других — тормозящим. Следовательно, можно ожидать, что эти системы оказывают разное действие на уровень возбудимости разных отделов мозга. Областью распределения норадреналиновой системы является практически весь мозг, тогда как серотониновая и дофаминовая системы имеют гораздо более специфическое распределение: дофаминовая система в основном направляется к регионам базальных ганглиев, а серотониновая система больше влияет на структуры, расположенные по средней линии.
б) Нейрогормональные системы мозга человека. На рисунке ниже представлены области мозгового ствола головного мозга человека для активации четырех нейрогуморальных систем, три из которых соответствуют выявленным у крыс, и одна — ацетилхолиновая система.
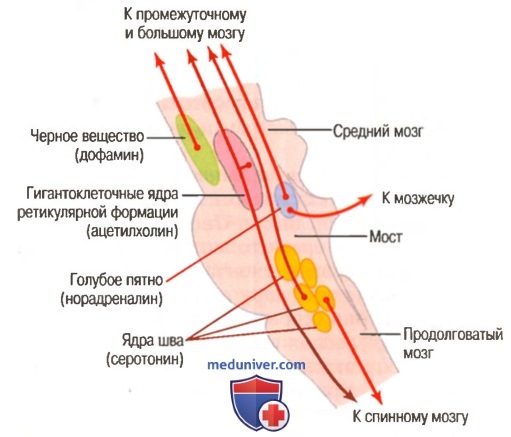
Совокупность центров ствола мозга, нейроны которых секретируют различные медиаторы (указаны в скобках). Эти нейроны посылают регулирующие сигналы вверх (в промежуточный и большой мозг) и вниз (в спинной мозг)
Далее перечислены некоторые специфические функции этих систем.
1. Голубое пятно и норадреналиновая система. Голубое пятно — небольшая область, расположенная с обеих сторон в задней части места соединения моста и среднего мозга. Нервные волокна, выходящие из этой области, секретируют норадреналин и распространяются по всему мозгу так же. В большинстве случаев норадреналин возбуждает мозг, увеличивая его активность. Однако в некоторых областях мозга его эффекты — тормозные из-за тормозных рецепторов в некоторых нервных синапсах. Эта система, вероятно, играет важную роль в возникновении сновидений, участвуя в развитии особого типа сна, называемого сном с быстрыми движениями глаз (REM-сон).
2. Черное вещество и дофаминовая система. Оно лежит в передне-верхней части среднего мозга, а его нейроны посылают нервные окончания главным образом к хвостатому ядру и скорлупе большого мозга, где они секретируют дофамин. Другие нейроны, расположенные в прилежащих областях, также секретируют дофамин, но посылают свои окончания в области мозга, расположенные вентральнее, особенно к гипоталамусу и лимбической системе. Полагают, что дофамин действует как тормозной медиатор в базальных ганглиях, но в некоторых других областях мозга его эффект, возможно, возбуждающий. Разрушение дофаминергических нейронов черного вещества является основной причиной болезни Паркинсона.
3. Ядра шва и серотониновая система. По средней линии моста и продолговатого мозга есть несколько тонких ядер, называемых ядрами шва. Многие нейроны этих ядер секре-тируют серотонин. Они посылают волокна в промежуточный мозг, некоторые из них идут к коре большого мозга, другие спускаются вниз к спинному мозгу. В спинном мозге серотонин, выделяемый из окончаний этих волокон, способен подавлять боль. В области промежуточного мозга и большого мозга серотонин почти наверняка оказывает мощное тормозное действие, помогая развитию нормального сна, что мы обсудим в отдельной статье на сайте (просим вас пользоваться формой поиска выше).
4. Гигантоклетонные нейроны возбуждающей области ретикулярной формации и ацетилхолиновая система. Ранее мы обсуждали гиган-токлеточные нейроны (гигантские клетки) возбуждающей области ретикулярной формации моста и среднего мозга. Волокна этих крупных нейронов сразу делятся на две ветви: одна направляется вверх к более высоким уровням мозга, а другая идет вниз в составе ретикулоспинального тракта к спинному мозгу. В терминалах этих волокон секретируется ацетилхолин. В большинстве случаев он действует как возбуждающий нейромедиатор. Активация этих холинергических нейронов ведет к внезапному пробуждению и возбуждению нервной системы.
в) Другие нейромедиаторы и нейрогормоны, секретируемые в мозге. Далее перечислены (без описания их функций) другие нейрогормональные вещества, функционирующие либо в специфических синапсах, либо путем выделения в жидкости мозга: энкефалины, гамма-аминомасляная кислота, глутамат, вазопрессин, адренокортикотропный гормон, адреналин, гистамин, эндорфины, ангиотензин II, нейротензин. Следовательно, в мозге имеется множество нейрогормональных систем, активация каждой из которых играет собственную роль в регуляции различных сторон мозговой деятельности.
Видео физиология гуморальной регуляции и ее отличие от гормональной - профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Читайте также: