
Физиологическая характеристика мышечной силы кратко
Обновлено: 02.07.2024
Мышечное сокращение. Характеристика и механизм мышечного сокращения
Выделяют несколько последовательных этапов запуска и осуществления мышечного сокращения.
1. Потенциал действия распространяется вдоль двигательного нервного волокна до его окончаний на мышечных волокнах.
2. Каждое нервное окончание секретирует небольшое количество нейромедиатора ацетилхолина.
3. Ацетилхолин действует на ограниченную область мембраны мышечного волокна, открывая многочисленные управляемые ацетилхолином каналы, проходящие сквозь белковые молекулы, встроенные в мембрану.
4. Открытие управляемых ацетилхолином каналов позволяет большому количеству ионов натрия диффундировать внутрь мышечного волокна, что ведет к возникновению на мембране потенциала действия.
5. Потенциал действия проводится вдоль мембраны мышечного волокна так же, как и по мембране нервного волокна.
6. Потенциал действия деполяризует мышечную мембрану, и большая часть возникающего при этом электричества течет через центр мышечного волокна. Это ведет к выделению из саркоплазматического ретикулума большого количества ионов кальция, которые в нем хранятся.
7. Ионы кальция инициируют силы сцепления между актиновыми и миозиновыми нитями, вызывающие скольжение их относительно друг друга, что и составляет основу процесса сокращения мыщц.
8. Спустя долю секунды с помощью кальциевого насоса в мембране саркоплазматического ретикулума ионы кальция закачиваются обратно и сохраняются в ретикулуме до прихода нового потенциала действия. Удаление ионов кальция от миофибрилл ведет к прекращению мышечного сокращения.
Далее мы обсудим молекулярные механизмы этого процесса.
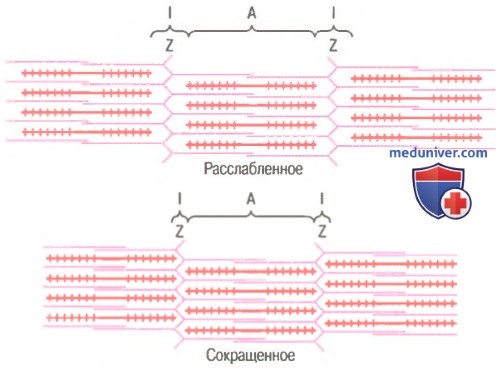
Миофибрилла в расслабленном и сокращенном состоянии. Показано (вверху), что актиновые нити (розовые) вдвинуты в пространства между миозиновыми нитями (красные). Сближение Z-дисков друг с другом (внизу).
Молекулярные механизмы мышечного сокращения
Механизм скольжения нитей для мышечного сокращения. На рисунке показан основной механизм мышечного сокращения. Показано расслабленное состояние саркомера (вверху) и сокращенное состояние (внизу). В расслабленном состоянии концы актиновых нитей, отходящие от двух последовательных Z-дисков, лишь незначительно перекрываются. Наоборот, в сокращенном состоянии актиновые нити втягиваются внутрь между миозиновыми так сильно, что их концы максимально перекрывают друг друга. При этом Z-диски притягиваются актиновыми нитями к концам миозиновых. Таким образом, мышечное сокращение осуществляется путем механизма скольжения нитей.
Что заставляет нити актина скользить внутрь среди нитей миозина? Это связано с действием сил, генерируемых при взаимодействии поперечных мостиков, исходящих от нитей миозина, с нитями актина. В условиях покоя эти силы не проявляются, однако распространение потенциала действия вдоль мышечного волокна приводит к выделению из саркоплазматическо-го ретикулума большого количества ионов кальция, которые быстро окружают миофи-бриллы. В свою очередь, ионы кальция активируют силы взаимодействия между нитями актина и миозина, в результате начинается сокращение. Для осуществления процесса сокращения необходима энергия. Ее источником являются высокоэнергетические связи молекулы АТФ, которая разрушается до АДФ с высвобождением энергии. В следующих разделах мы приведем известные детали молекулярных процессов сокращения.
Молекулярные особенности сократительных нитей
Миозиновая нить. Она состоит из множества молекул миозина, молекулярная масса каждой составляет около 480000. На рисунке показана отдельная молекула; и также — объединение многих молекул миозина в миозиновую нить, а также взаимодействие одной стороны этой нити с концами двух актиновых нитей.
В состав молекулы миозина входят 6 полипептидных цепей: 2 тяжелые цепи с молекулярной массой около 200000 каждая и 4 легкие цепи с молекулярной массой около 20000 каждая. Две тяжелые цепи спирально закручиваются вокруг друг друга, формируя двойную спираль, которую называют миозиновым хвостом. С одного конца обе цепи изгибаются в противоположных направлениях, формируя глобулярную полипептидную структуру, называемую миозиновой головкой. Таким образом, на одном конце двойной спирали молекулы миозина образуются 2 свободные головки; 4 легкие цепи также включены в состав миозиновой головки (по 2 в каждой). Они помогают регулировать функцию головки во время мышечного сокращения.
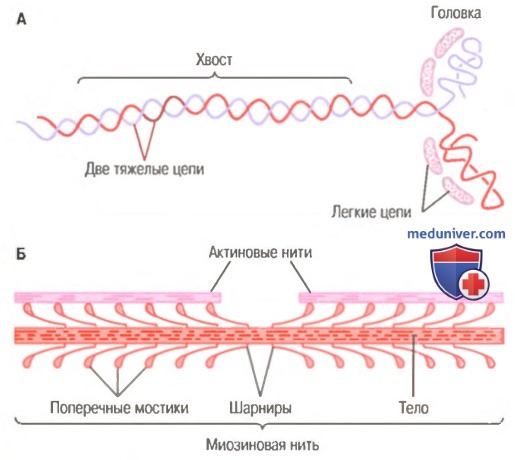
А. Молекула миозина.
Б. Объединение многих молекул миозина в одну миозиновую нить.
Показаны также тысячи миозиновых поперечных мостиков и взаимодействие их головок с прилежащими актиновыми нитями.
Миозиновая нить состоит из 200 или более отдельных молекул миозина. Видно, что хвосты молекул миозина объединяются, формируя тело нити, а многочисленные головки молекул выдаются наружу по сторонам тела. Кроме того, наряду с головкой в сторону выступает часть хвоста каждой миозиновой молекулы, образуя плечОу которое выдвигает головку наружу от тела, как показано на рисунке. Выступающие плечи и головки вместе называют поперечными мостиками. Каждый поперечный мостик может сгибаться в двух точках, называемых шарнирами. Один из них расположен в месте, где плечо отходит от тела миозиновой нити, а другой — где головка крепится к плечу. Движение плеча позволяет головке или выдвигаться далеко наружу от тела миозиновой нити, или приближаться к телу. В свою очередь, повороты головки участвуют в процессе сокращения, что обсуждается в следующих разделах.
Общая длина каждой миозиновой нити остается постоянной и равна почти 1,6 мкм. В самом центре миозиновой нити на протяжении 0,2 мкм поперечных мостиков нет, поскольку снабженные шарнирами плечи отходят в стороны от центра.
Сама миозиновая нить сплетена таким образом, что каждая последующая пара поперечных мостиков смещена в продольном направлении относительно предыдущей на 120°, что обеспечивает распределение поперечных мостиков во всех направлениях вокруг нити.
АТФ-азная активность миозиновой головки. Есть и другая особенность миозиновой головки, необходимая для мышечного сокращения: миозиновая головка функционирует как фермент АТФ-аза. Как объясняется далее, это свойство позволяет головке расщеплять АТФ и использовать энергию расщепления высокоэнергетической связи для процесса сокращения.
Актиновая нить. Актиновая нить состоит из трех белковых компонентов: актина, тропомиозина и тропонина.
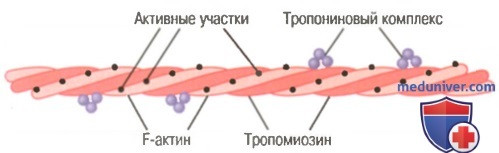
Актиновая нить, состоящая из двух спиралевидных цепочек молекул F-актина и двух цепочек молекул тропомиозина, расположенных в желобках между цепочками актина.
К одному концу каждой молекулы тропомиозина прикреплен тропониновый комплекс, который запускает сокращение.
Основой актиновой нити являются две цепи белковой молекулы F-актина. Обе цепи закручиваются в спираль так же, как и молекула миозина.
Каждая цепь двойной спирали F-актина состоит из полимеризованных молекул G-актина с молекулярной массой около 42000. К каждой молекуле G-актина прикреплена 1 молекула АДФ. Полагают, что эти молекулы АДФ являются активными участками на актиновых нитях, с которыми взаимодействуют поперечные мостики миозиновых нитей, обеспечивая мышечное сокращение. Активные участки на обеих цепях F-актина двойной спирали расположены со смещением таким образом, что вдоль всей поверхности актиновой нити встречается один активный участок примерно через каждые 2,7 нм.
Длина каждой актиновой нити — около 1 мкм. Основания актиновых нитей прочно встроены в Z-диски; концы этих нитей выступают в обоих направлениях, располагаясь в пространствах между миозиновыми молекулами.
Молекулы тропомиозина. Актиновая нить также содержит другой белок — тропомиозин. Каждая молекула тропомиозина имеет молекулярную массу 70000 и длину 40 нм. Эти молекулы спирально оплетают спираль из F-актина. В состоянии покоя молекулы тропомиозина располагаются поверх активных участков актиновых нитей, препятствуя их взаимодействию с миозиновыми нитями, лежащему в основе сокращения.
Тропонин и его роль в мышечном сокращении. По ходу молекул тропомиозина к ним периодически прикреплены другие белковые молекулы, называемые тропонином. Они представляют собой комплексы трех слабосвязанных белковых субъединиц, каждая из которых играет специфическую роль в регуляции мышечного сокращения. Одна из субъединиц (тропонин I) имеет высокое сродство к актину, другая (тропонин Т) — к тропомиозину, третья (тропонин С) — к ионам кальция. Считают, что этот комплекс прикрепляет тропомиозин к актину. Высокое сродство тропонина к ионам кальция, как полагают, инициирует процесс сокращения, о чем говорится в следующей статье.
Видео физиология мышц и мышечного сокращения - профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Биохимия и физиология мышечной активности при выполнении физической работы может быть описана следующим образом. Покажем с помощью имитационного моделирования как разворачиваются физиологические процессы в мышце при выполнении ступенчатого теста.
При достижении внешней мощности некоторого значения наступает момент, когда в работу вовлекаются все ММВ и начинают рекрутироваться промежуточные мышечные волокна (ПМВ). Промежуточными мышечными волокнами можно назвать те, в которых массы митохондрий недостаточно для обеспечения баланса между образованием пирувата и его окислением в митохондриях. В ПМВ после снижения концентрации фосфогенов активизируется гликолиз, часть пирувата начинает преобразовываться в молочную кислоту (точнее говоря, в лактат и ионы водорода), которая выходит в кровь, проникает в ММВ. Попадание в ММВ (ОМВ) лактата ведет к ингибированию окисления жиров, субстратом окисления становится в большей мере гликоген. Следовательно, признаком рекрутирования всех ММВ (ОМВ) является увеличение в крови концентрации лактата и усиление легочной вентиляции. Легочная вентиляция усиливается, в связи с образованием и накоплением в ПМВ ионов водорода, которые при выходе в кровь взаимодействуют с буферными системами крови и вызывают образование избыточного (неметаболического) углекислого газа. Повышение концентрации углекислого газа в крови приводит к активизации дыхания (Физиология человека, 1998).
Таким образом, при выполнении ступенчатого теста имеет место явление, которое принято называть аэробным порогом (АэП). Появление АэП свидетельствует о рекру-тировании всех ОМВ. По величине внешнего сопротивления можно судить о силе ОМВ, которую они могут проявить при ресинтезе АТФ и КрФ за счет окислительного фосфори-лирования (Селуянов В. Н. с соав., 1991).
Дальнейший рост внешней мощности делает необходимым вовлечение все более высокопороговых ДЕ, иннервирующих гликолитические МВ. Динамическое равновесие нарушается, продукция Н, лактата начинает превышать скорость их устранения. Это сопровождается дальнейшим увеличением легочной вентиляции, ЧСС и потребления кислорода. После АнП потребление кислорода в основном связано с работой дыхательных мышц и миокарда. При достижении предельных величин легочной вентиляции и ЧСС или при локальном утомлении мышц потребление кислорода стабилизируется, а затем начинает уменьшаться. В этот момент фиксируют МПК.
Таким образом, МПК есть сумма величин потребления кислорода окислительными МВ (ММВ), дыхательными мышцами и миокардом.
Описаны физиологические факторы, определяющие силу скелетных мышц человека: частота импульсации ДЕ, количество активных ДЕ, синхронизация активности ДЕ. Показано, как принцип активации ДЕ (принцип размера или правило Хенеманна) влияет на количество ДЕ, которые активны при выполнении силовых упражнений с различными отягощениями.
От чего зависит сила мышц? (физиологические факторы)
Итак, мы разобрались с анатомическими факторами, определяющими силу мышц. Можно сказать, что это тот морфологический потенциал, который зависит от генетики человека (например, число мышечных волокон), его пола, возраста и функционального состояния, которое определяется его образом жизни (малоподвижный или систематичные тренировки).
Однако вы хорошо понимаете, что каждый человек может сам, произвольно регулировать силу, которую проявляют его мышцы. Это означает, что наша центральная нервная система обладает такими механизмами. Назовем их физиологическими механизмами регуляции силы и скорости сокращения мышц.
Физиологические механизмы регуляции силы и скорости сокращения мышц
Учеными доказано, что управлять уровнем силы и скоростью сокращения мышц мы можем посредством следующих параметров работы центральной нервной системы (ЦНС):
- частоты разрядов двигательной единицы (ДЕ);
- числа активных ДЕ;
- синхронизации работы ДЕ.
Частота разрядов ДЕ
Частота разрядов ДЕ представляет собой не что иное как частоту разрядов мотонейрона, иннервирующего скелетную мышцу. Именно посредством увеличения частоты импульсации мотонейрона мышца получает команду о том, что она должна сокращаться сильнее.
Одиночное сокращение мышцы
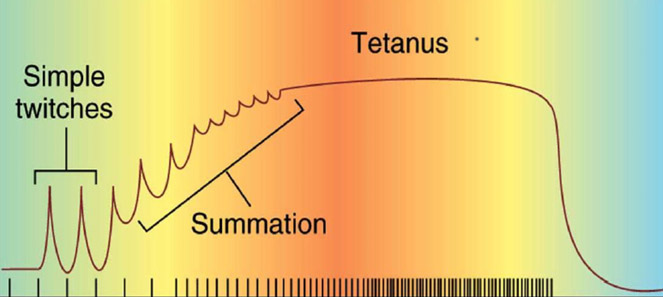
Для того, чтобы понять, как влияет частота импульсации мотонейрона на мышечные волокна, ученые стимулировали скелетную мышцу или нерв, идущий к мышце, током различной частоты. Было установлено, (рис.1), что при низкой частоте стимуляции (до трех импульсов в секунду, то есть до 3 Гц мышечные волокна, входящие в ДЕ производят одиночное сокращение. Это означает, что они успевают сократиться и расслабиться. Установлено, что фаза расслабления в 1,5-2 раза более продолжительна, чем фаза сокращения. Сила, которую развивает мышца небольшая (рис.1).
Зубчатый тетанус
Гладкий тетанус
При частоте импульсации 20-30 имп./с (30 Гц) каждый последующий импульс, поступающий от мотонейрона попадает на фазу сокращения мышечных волокон. Возникает их сильное сокращение. Сила, развиваемая мышцей достигает 30-60 % от максимальной. Это называется гладким тетанусом.
Было установлено, что зависимость силы мышцы от частоты стимуляции нелинейная. Вначале при частотах от 5 до 30 Гц сила мышцы резко возрастает до 90% от максимальной. Дальнейшее повышение частоты стимуляции (до 60 Гц), дает прибавку силы мышцы в только 10%.
Число активных ДЕ
Вначале рекрутируются ДЕ S типа, иннервируемые мотонейронами, имеющими небольшой диаметр аксона. В состав этих ДЕ входят самые медленные мышечные волокна типа I. Мышца развивает небольшую силу.
Затем активируются ДЕ типа FR, содержащие быстрые неутомляемые волокна типа IIА . Увеличение количества активных ДЕ приводит к росту силы мышцы.
Последними рекрутируются ДЕ типа FF, в состав которых входят быстрые, быстроутомляемые волокна типа IIВ. Эти волокна иннервируют мотонейроны, имеющие самый большой диаметр аксона. Сила, которую может развить мышца при произвольном сокращении достигает максимума.
Считается, что принцип размера позволяет осуществлять тонкую градацию силы мышцы во всем физиологическом диапазоне.
Принцип размера объясняет почему, используя небольшие отягощения, невозможно эффективно наращивать силу мышц. Это связано с тем, что при применении небольших отягощений, рекрутируются только ДЕ типа S, в состав которых входят медленные мышечные волокна. То есть активны только медленные мышечные волокна. Те мышечные волокна, которые активны, те и тренируются. Если мышце необходимо развить большую силу, в сокращение вовлекаются все типы ДЕ, в состав которых входят мышечные волокна всех типов. Это вызывает процессы адаптации в мышце (мышечные волокна повреждаются, а потом восстанавливаются). В результате возрастает поперечное сечение мышцы и она способна развивать большую силу.
Синхронизация активности ДЕ
Установлено, что при длительной, но не очень интенсивной работе, отдельные ДЕ сокращаются попеременно (асинхронно). Такой тип активности имеет место, например, при беге на длинные дистанции или марафонском беге (А.С. Солодков, Е.Б. Сологуб, 2005). Утомление в этом случае развивается медленно, так как работая по очереди, ДЕ в промежутках между активацией успевают восстанавливаться.
Однако, если мышце требуется развить мощное, кратковременное усилие (например, выполнить жим лежа штанги большой массы), требуется синхронизация активности отдельных ДЕ. Естественно одновременная синхронная работа большого количества ДЕ приводит к значительному увеличению силы мышцы.
Ю. Хартман и Х. Тюннеманн (1988) показали, что новичок одновременно может задействовать до 60% ДЕ, в то время как спортсмен высокой квалификации может обеспечить синхронную активность до 85% ДЕ. Из этого следует, что тренировка силовой направленности повышает степень синхронной активности ДЕ.
Литература
- Мак-Комас А. Дж. Скелетные мышцы (Строение и функции). – Киев: Олимпийская литература, 2001.– 408 с.
- Солодков А.С., Сологуб Е.С. Физиология человека. Общая. Спортивная. Возрастная: Учебник. М.: Олимпия Пресс, 2005.– 528 с.
- Хартманн Ю. Тюннеманн Х. Современная силовая тренировка. Берлин: Шпортферлаг. 1988. 335 с.
Похожие записи:
Тест времени реакции на сигнал
Представлена программа расчета времени реакции на сигнал, предназначенная для использования в учебных целях, например на занятиях по…
Саркоплазматическая гипертрофия мышц
Дано определение и описаны механизмы саркоплазматической гипертрофии скелетных мышц. Показано, что этот вид гипертрофии мышц широко…
Классификация типов конституции человека М.В. Черноруцкого
Рассмотрена классификация типов конституции человека, разработанная выдающимся терапевтом М.В. Черноруцким в 1925 году. Классификация типов конституции человека М.В.
Типы гипертрофии скелетных мышц человека
В статье дается классификация различных видов гипертрофии скелетных мышц человека на основе ряда классификационных признаков: времени проявления…
Миомейкер: Мембранный активатор слияния миобластов и образования мышц
Ученые установили, что для образования мышечных волокон необходимо слияние клеток-предшественников, которые называются миобластами. Эти клетки имеют только…
Отсроченное начало болезненности мышц. Стратегии лечения и факторы эффективности
Описаны симптомы, причины, теории отсроченного начала болезненности мышц (запаздывающих болезненных ощущений, DOMS), а также способы уменьшения этих болей:…
Срочные гормональные ответы у элитных тяжелоатлетов-юниоров
Изучалось изменение концентрации в крови: тестостерона, кортизола, гормона роста, бета-эндорфина и лактата у тяжелоатлетов-юниоров…
Effect of KAATSU-training on the maximum voluntary isometric contraction of lower extremity muscles of qualified football players
Изучалось влияние KAATSU-тренинга на изометрическую силу мышц квалифицированных футболистов. Установлено, что интенсивный рост максимальной силы мышц…
Читайте также: