
За счет чего при тренировках возрастает точность мышечных сокращений кратко
Обновлено: 05.07.2024
Самсонова, А.В. Гипертрофия скелетных мышц человека под воздействием различных средств и методов силовой тренировки /А.В.Самсонова // Научно-педагогические школы университета. Научные труды. Ежегодник 2014.- СПб: НГУ им. П.Ф.Лесгафта.- С.11-23.
Самсонова А.В.
Гипертрофия скелетных мышц человека под воздействием различных средств и методов силовой тренировки
Введение
Гипертрофия скелетных мышц (увеличение их массы или объема) является предметом исследования многих научных дисциплин, таких как: анатомия, гистология, биохимия, физиология, спортивная медицина, биомеханика и конечно, атлетизм. Благодаря тому накоплен большой арсенал научных знаний об этом феномене. Столь пристальное внимание к этому феномену связано с тем, что гипертрофия скелетных мышц лежит в основе увеличении их силы. Однако анализ существующей научно-методической литературы свидетельствует о том, что до настоящего времени отсутствует целостная концепция, объясняющая механизм воздействия физической нагрузки на гипертрофию скелетных мышц. В связи с этим цель исследования состояла в разработке концепции гипертрофии скелетных мышц под воздействием различных средств и методов силовой тренировки с позиций системного подхода.
Результаты
- Влияние на гипертрофию скелетных мышц отягощений различной массы методом повторных непредельных усилий
Уже в середине ХХ века в реабилитационной медицине был установлен факт быстрого восстановления массы и силы скелетных мышц после их атрофии посредством тренировки с использованием значительных внешних отягощений (70-80% от максимума). Тренировка с малыми отягощениями (30-40% от максимума), такого эффекта не вызывала (T.L. DeLorme, 1945). Найденная эмпирическим путем закономерность в настоящее время активно используется в атлетизме (А.Н. Воробьев, 1988; V.M. Zatsiorsky, W.J. Kraemer, 2006; Г.П. Виноградов, 2009). Чтобы понять механизм воздействия отягощений различной массы на увеличение объема скелетных мышц необходимо привлечь знания из ряда медико-биологических дисциплин. Из анатомии и гистологии известно, что скелетные мышцы человека состоят из мышечных волокон (МВ) различного типа: медленных (I типа) и быстрых (IIA и IIВ типа). МВ I типа характеризуются невысокой скоростью и силой сокращения, однако способность сопротивляться утомлению у них очень высока. Мышечные волокна IIВ типа способны сокращаться с большой силой и скоростью, однако сопротивление утомлению у них низкое. Волокна IIA типа характеризуются промежуточными свойствами. Основу сократительного аппарата мышечных волокон скелетных мышц человека составляют миофибриллы, которые состоят из секций – саркомеров, отделенных друг от друга Z-дисками. Гистологами установлено, что медленные волокна I типа имеют более толстые Z-диски по сравнению с быстрыми волокнами II типа (H. Hoppeler, 1986). Между собой миофибриллы на уровне Z-дисков связаны цитоскелетными белками. Помимо этого периферические миофибриллы прикреплены к внутренней оболочке мышечного волокна посредством костамеров (рис.1). Саркомер подобен связке шестигранных карандашей (в саркомере их больше 300), грифелем которых служит толстый филамент, а ребрами – тонкие филаменты. При сокращении мышцы тонкие филаменты скользят относительно толстых, расстояние между Z-дисками уменьшается, саркомер укорачивается. Одновременное сокращение всех саркомеров приводит к уменьшению длины миофибриллы и мышечного волокна. Ввиду того, что саркомер представляет собой не плоскую, а объемную структуру, при его сокращении происходит также увеличение площади его поперечного сечения (когда тонкие нити входят в промежутки между толстыми), площади поперечного сечения мышечных волокон и всей мышцы. Физиологами найдено, что управление мышцей со стороны ЦНС осуществляется посредством активации двигательных единиц (ДЕ). По классификации Р. Берка с соавт. (R.E. Burke et all. 1973) ДЕ делятся на три типа: S (slow) – медленные, устойчивые к утомлению; FR (fast resistant) – быстрые, устойчивые к утомлению, FF – fast fatigable – быстрые, быстроутомляемые. ДЕ различных типов соответствуют различные виды мышечных волокон. В состав двигательных единиц типа S входят мышечные волокна I типа. В состав двигательных единиц типа FR – IIA типа. В состав двигательных единиц типа FF – входят мышечные волокна IIB типа.
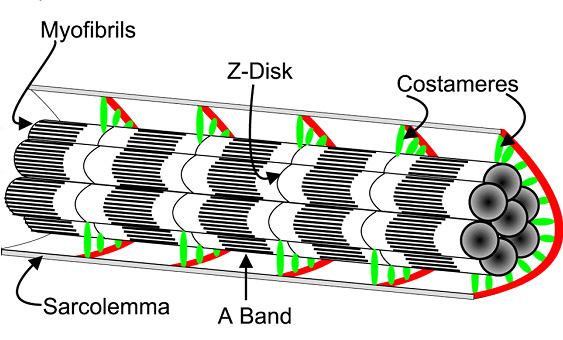
Рис.1. Прикрепление миофибрилл посредством костамеров к сарколемме мышечного волокна (J.M.Ervasty, 2003)
Количество ДЕ, активных в процессе сокращения мышцы, определяется посредством центральных и рефлекторных механизмов регуляции силы мышц (А.Г. Фельдман, 1979). Установлено, что имеется стабильный порядок вовлечения в работу (рекрутирования) ДЕ: вначале рекрутируются ДЕ S типа, затем FR типа, последними в сокращение вовлекаются ДЕ FF типа (E. Henneman, C.B. Olson, 1965). Концепция, объясняющая воздействие больших отягощений на увеличение силы и гипертрофию скелетных мышц состоит в следующем. Если внешнее отягощение небольшое (например, масса штанги составляет менее 20% максимума) при выполнении силового упражнения рекрутируются только мышечные волокна I типа, при этом уровень силы, развиваемый мышцей, невысокий (В.С. Гурфинкель, Ю.С. Левик, 1985). Для преодоления большого внешнего отягощения (более 70% от максимума) мышца должна развить большую силу. Поэтому в сокращение последовательно вовлекаются все типы мышечных волокон, в том числе и IIB типа (рис.2).
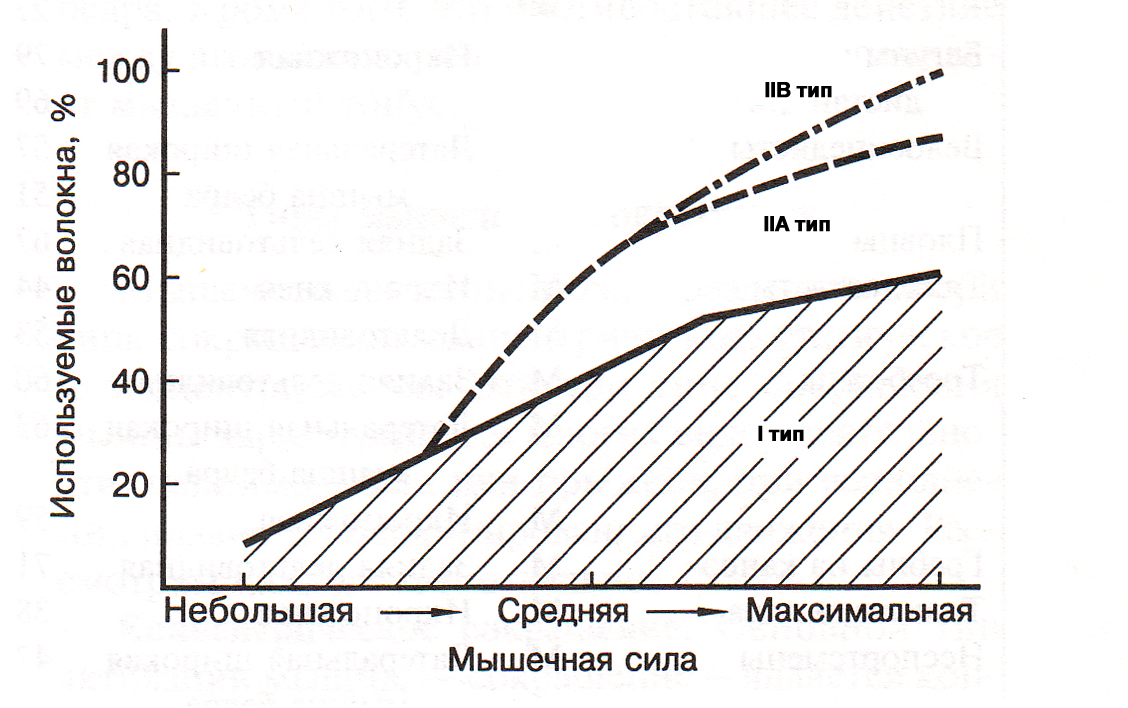
Рис.2. Зависимость между развиваемой силой мышцы и вовлечением в работу медленных и быстрых мышечных волокон (Дж. Х.Уилмор, Д.Л. Костилл, 1997)
Специалистами в области спортивной медицины установлено, что тренировка с применением больших отягощений вызывает мышечные боли у спортсменов, как во время, так и после ее окончания (Б.И. Прилуцкий, 1989; В.И. Морозов, Г.А. Сакута, М.И. Калинский, 2006; Г.А. Макарова, 2008). В настоящее время накоплено достаточное количество фактов, свидетельствующих о том, что болезненные ощущения в мышцах в первую очередь связаны с их повреждением. Повреждение мышечных волокон вызывает воспаление, что ощущается как боль в мышцах через 24 часа и более. Последующая регенерация мышечных волокон приводит к их гипертрофии.
- Влияние на гипертрофию скелетных мышц тренировки с использованием различных режимов мышечного сокращения
Различают несколько режимов сокращения мышц: преодолевающий (концентрический) – длина мышцы уменьшается, уступающий (эксцентрический) – длина мышцы увеличивается, изометрический (статический) – длина мышцы не изменяется. P.J. Rasch и L.J. Morehouse (1957) одними из первых показали, что динамические упражнения по сравнению с изометрическими вызывают больший прирост силы и площади поперечного сечения мышц. Показано, что тренировка с использованием эксцентрического режима приводит к несколько большей гипертрофии мышц, чем другие режимы сокращения (M.J.Gibala et al. 1995; Е. Hagbie et al. 1996; J.Y. Seger, B. Arvidsson, A. Thorstensson, 1998; M.J. Gibala et al. 2000). Установлено, что гипертрофия мышечных волокон при работе в эксцентрическом режиме больше, чем при концентрическом (S.L.Lindstedt, P.C. LaStayo, T.E. Reich, 2001).
Исследования, проведенные на гистологическом уровне, свидетельствуют о том, что при выполнении упражнений в эксцентрическом режиме в большей степени повреждается цитоскелет и Z-диски мышечного волокна по сравнению с тренировкой в других режимах (J. Friden, M. Sjostrom, B. Ekblom, 1983; J.Friden, R.L. Lieber, 2001), рис.3. Установлено, что степень повреждения Z-дисков мышечных волокон II типа в три раза больше, чем у волокон I типа (J. Friden, M. Sjostrom, B. Ekblom, 1983).

Рис.3. Электронная фотография мышечного волокна человека после выполнения эксцентрических упражнений. Обозначения: * — миофибриллы с разрушенными Z-дисками; ○ – миофибрилла с неповрежденным Z диском. (J.G. Yu, L.Carlsson, L.E. Thomell, 2004)
Исследованиями M.J. Gibala etal. (1995) показано, что даже однократная силовая тренировка в эксцентрическом режиме вызывает у начинающих спортсменов повреждение более 82% мышечных волокон, а в концентрическом – только 33%, при этом во время эксцентрических сокращений суммарная ЭАМ на 40% меньше, чем во время концентрических. У хорошо тренированных спортсменов аналогичная тренировка приводит к 45% повреждений мышечных волокон при работе в эксцентрическом режиме и 27% при работе в концентрическом режиме (M.J. Gibala et al. 2000). Биохимические данные свидетельствуют о том, что на третий-пятый день после эксцентрических сокращений, в крови исследуемых значительно возрастает уровень креатинкиназы и миоглобина, что свидетельствует о сильных повреждениях мышечных волокон. При этом он значительно превышает аналогичные показатели, полученные при выполнении движений в концентрическом режиме (J.A. Faulkner, S.V. Brooks, J.A. Opiteck, 1993; A.P. Lavender, K. Nosaka, 2006; K. Nosaka, 2008). На основе изучения биомеханических характеристик развития усилия мышцей M.J. Gibala et al. (1995) было показано (рис.4), что после одного тренировочного занятия силовыми упражнениями значения максимального момента силы двуглавой мышцы плеча, измеренного в изометрическом режиме, понизились как у руки, выполнявшей движения в концентрическом, так и у руки, выполнявшей движения в эксцентрическом режимах сокращения (р≤0,05). Однако через 24 часа этот показатель, измеренный у руки, работающей в концентрическом режиме достоверно не отличался от базового уровня (р>0,05). В то же время значения максимального момента силы, у руки, выполняющей эксцентрические сокращения, достоверно отличались от начального уровня через 24, 48, 72 и даже 96 часов (р≤0,05). Следует отметить, что растяжение пассивной мышцы не приводит к ее гипертрофии (J.R. Fowles et al. 2000).
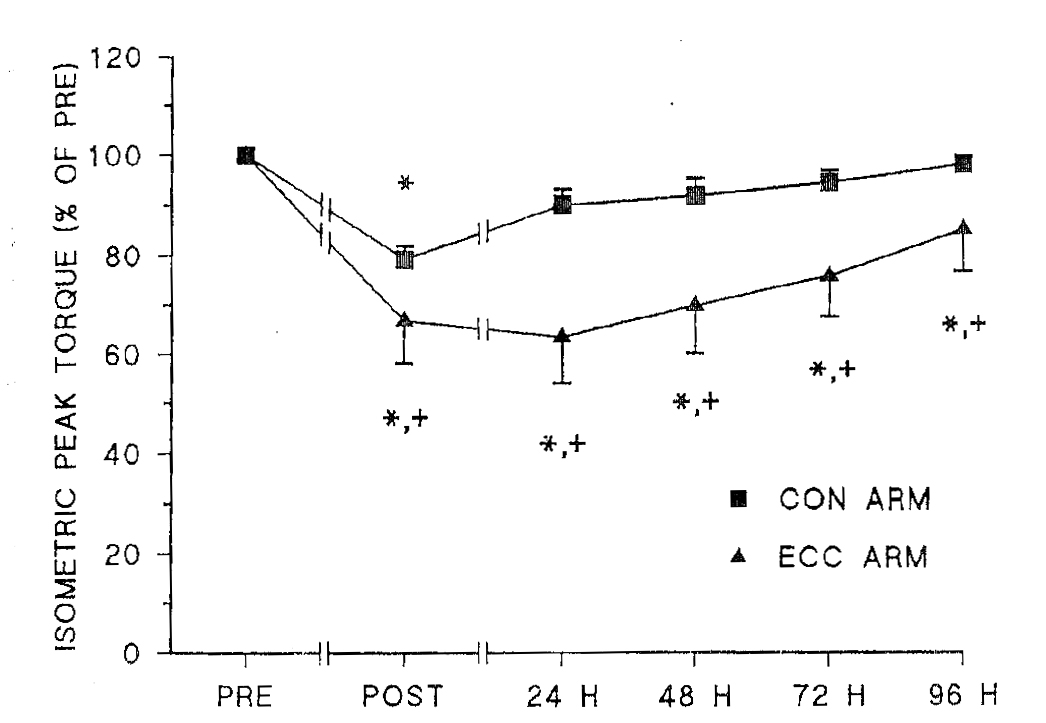
Рис. 4. Максимальные значения момента силы, полученного в изометрическом режиме для руки, выполнявшей концентрические сокращения (квадраты) и руки, выполнявшей эксцентрические сокращения (треугольники) в различные моменты времени: до (PRE), сразу после (POST),через 24 часа (24H),48 часов (48H), 72 часа (72H) и 96 часов (96H) после выполнения упражнения. Представлены средние значения и ошибка среднего до и после выполнения упражнения * – различия между значениями, полученными до и после проведения упражнений достоверны на уровне значимости р≤0,05. + – различия статистически достоверны на уровне значимости р≤0,05 между CON рукой и Eсс рукой (по: M.J.Gibala et al. (1995).
Выводы и заключение
Литература
Похожие записи:
Саркоплазматическая гипертрофия мышц
Дано определение и описаны механизмы саркоплазматической гипертрофии скелетных мышц. Показано, что этот вид гипертрофии мышц широко…
Эффективность мышечного сокращения. Сокращение целой мышцы
Эффективность двигателя или автомашины рассчитывают как процент потребляемой энергии, которая превращается в работу вместо тепла. В мышцах количество энергии, способной превращаться в работу, даже при наилучших условиях составляет менее 25% всей энергии, доставляемой к мышце (химической энергии питательных веществ), а остальная энергия превращается в тепло. Причина этой низкой эффективности связана с тем, что примерно половина энергии питательных веществ теряется во время образования АТФ, и только 40-45% энергии самой АТФ может позднее превратиться в работу.
Максимальная эффективность реализуется лишь при условии сокращения мышцы с умеренной скоростью. При медленном сокращении мышцы или без какого-либо ее укорочения во время сокращения освобождается небольшое количество поддерживающего тепла, хотя работа практически не выполняется, что снижает эффективность преобразования до нуля. Напротив, если сокращение слишком быстрое, большая доля энергии используется на преодоление вязкого трения внутри самой мышцы, и это также снижает эффективность сокращения. Обычно максимальная эффективность развивается, когда скорость сокращения составляет около 30%.
Характеристики сокращения целой мышцы
Многие особенности сокращения мышцы можно продемонстрировать на примере одиночных мышечных сокращений. Такие сокращения вызывают с помощью одиночного электрического возбуждения, иннервирующего мышцу нерва, или короткого электрического раздражения самой мышцы, что ведет к развитию одиночного сокращения, продолжающегося долю секунды.
Изотоническая и изометрическая системы для регистрации мышечного сокращения. Длительность изометрических сокращений различных типов скелетных мышц млекопитающих. Показан также латентный период между потенциалом действия (деполяризацией) и мышечным сокращением.
Изометрическое и изотоническое сокращение. Мышечное сокращение называют изометрическим, если мышца не укорачивается во время сокращения, и изотоническим — если мышца укорачивается, но ее напряжение на протяжении всего сокращения остается постоянным.
В изометрической системе мышца сокращается без уменьшения своей длины, а в изотонической системе мышца укорачивается против фиксированной нагрузки: мышца поднимает чашу весов с разновесом. Изометрическая система строго регистрирует изменения силы самого мышечного сокращения, а параметры изотонического сокращения зависят от нагрузки, против которой мышца сокращается, а также от инерции нагрузки. В связи с этим при сравнении функциональных особенностей различных типов мышц чаще всего используют изометрическую систему.
На рисунке показаны кривые регистрации изометрических сокращений трех типов скелетных мышц: глазной мышцы (длительность изометрического сокращения менее 1/40 сек), икроножной мышцы (длительность сокращения около 1/15 сек) и камбаловиднй мышцы (длительность сокращения примерно 1/3 сек). Интересно, что эти длительности сокращений приспособлены к функциям соответствующих мышц. Движения глаз должны быть чрезвычайно быстрыми, чтобы поддерживать фиксацию глаз на объекте для обеспечения ясного видения. Икроножная мышца должна сокращаться умеренно быстро, чтобы обеспечить скорость движения нижней конечности, достаточную для бега или прыжков. А камбаловидная мышца имеет дело в основном с медленными сокращениями для непрерывной длительной поддержки тела против силы тяжести.
Быстрые и медленные мышечные волокна. Как обсуждается в предыдущих статьях, посвященных спортивной физиологии, каждая мышца тела состоит из совокупности так называемых быстрых и медленных мышечных волокон, а также других волокон с переходными свойствами. В состав быстрореагирующих мышц входят в основном быстрые волокна и лишь небольшое число медленных. И наоборот, медленнореагирующие мышцы составлены главным образом из медленных волокон. Различия между этими двумя типами волокон следующие.
Быстрые волокна: (1) крупные волокна, обеспечивающие большую силу сокращения; (2) имеют хорошо развитый саркоплазматический ретикулум для быстрого выделения ионов кальция, инициирующих сокращение; (3) содержат большое количество гликолитических ферментов для быстрого освобождения энергии путем гликолиза; (4) имеют сравнительно бедное кровоснабжение, поскольку окислительный метаболизм имеет второстепенное значение; (5) содержат немного митохондрий также в связи со второстепенностью окислительного метаболизма.
Медленные волокна: (1) более мелкие волокна; (2) иннервируются также более мелкими нервными волокнами; (3) имеют хорошо развитую систему кровеносных сосудов и капилляров для доставки большого количества кислорода; (4) содержат значительно больше митохондрий для обеспечения высоких уровней окислительного метаболизма; (5) содержат большое количество миоглобина — железосодержащего белка, подобного гемоглобину эритроцитов. Миоглобин связывается с кислородом и хранит его до момента, когда в нем возникнет потребность (это также значительно увеличивает скорость транспорта кислорода в митохондрии). Миоглобин придает медленным волокнам красноватый вид, поэтому их называют красными волокнами, а из-за дефицита красного миоглобина в быстрых волокнах их называют белыми волокнами.
Видео физиология мышц и мышечного сокращения - профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
В статье изложен обзор некоторых литературных данных о механизмах работы мышц во время различных по интенсивности нагрузок. Проведён анализ адаптивности, пластичности и степени изменения мышц на разных уровнях под влиянием различных факторов. Эти изменения связаны с увеличением мускульной силы и сопровождаются трансформацией одних типов мышечных волокон в другие, изменением метаболических процессов и увеличением синтеза сократительных белков, что в основном и обуславливает процессы мышечной гипертрофии. Главным пусковым механизмом данных процессов является синтез ростовых факторов, которые выполняют, по сути, основополагающую роль. Кроме усиления синтеза белка, данные факторы способны влиять и на миосателлиты. Именно эти клетки, располагающиеся на периферии симпласта, способны делиться в постнатальном периоде. В условиях наличия нагрузки и как следствия появления развивающихся микротравм происходит их активация. Они составляют основу регенерации поврежденного мышечного участка. За счёт своего деления они формируют новые мышечные волокна. Данный процесс очень напоминает воспалительный ввиду участия провоспалительных клеток. Их цитокины также влияют на пролиферацию. Поэтому при мышечной работе происходит множество процессов, которые способствуют к непосредственному увеличению мышечного волокна путём как за счёт гипертрофии, так и гиперплазии.

3. Lin Y. et al. PGC-1? is associated with C2C12 Myoblast differentiation // Central European Journal of Biology. 2014. Т. 9. № 11. Р. 1030–1036.
4. Scharf M. et al. Mitogen-activated protein kinase-activated protein kinases 2 and 3 regulate SERCA2a expression and fiber type composition to modulate skeletal muscle and cardiomyocyte function // Molecular and cellular biology. 2013. Т. 33. № 13. Р. 2586–2602.
5. Meissner J.D. et al. The p38?/? mitogen-activated protein kinases mediate recruitment of CREB-binding protein to preserve fast myosin heavy chain IId/x gene activity in myotubes // Journal of Biological Chemistry. 2007. Т. 282. № 10. Р. 7265–7275.
6. Potthoff M.J., Olson E.N. MEF2: a central regulator of diverse developmental programs // Development. 2007. Т. 134. № 23. Р. 4131–4140.
На протяжении многих лет изучение процессов синтеза белков в скелетных мышцах при выполнении различных физических нагрузок остаётся актуальной проблемой биохимии и физиологии. Мышцы и их силовые характеристики очень важная составляющая организма каждого спортсмена, которая позволяет достигать результатов. В связи с прогрессивным развитием спорта и вовлечением большого количества людей в физическую культуру, тема здоровья спортсменов становится все более актуальной, интересной и увлекательной. Учитывая существующую сильную корреляцию между площадью поперечного сечения мышц и мышечной силой, стремление увеличить мышечную массу тела есть у каждого человека, занимающегося спортом. Кроме этого, необходимо помнить, что преобладание мышечной массы в организме благоприятно влияет на метаболические процессы.
Скелетная мышца – одна из наиболее пластичных структур в организме млекопитающих. При повышенной активности и нагрузке часто происходит увеличение её размеров, объёмов миофибриллярного аппарата, повышение сократительных возможностей (силы, мощности). Процесс прироста мышечной массы зависит от различных факторов: наследственных, конституциональных, а также пола, возраста, метаболизма, гормонального фона. Кроме того, с приобретением опыта тренировок становится все труднее увеличить мышечную массу, поэтому важно понимать и активно использовать все возможные механизмы этого процесса.
Клетки поперечно-полосатой мускулатуры отличаются от гладкомышечных миоцитов. Клетки скелетных мышц образуют многоядерный синцитий, основное вещество которого формируют миофибриллы, состоящие из толстых и тонких миофиламентов. Первый тип образуют молекулярные единицы и миозин, а второй тип содержит тропомиозин с тропонином и F-актин. Многие авторы считают скелетную мускулатуру гетерогенной системой относительно устройства и выполняемых функций, несмотря на её строгую организацию. Данное свойство помогает мышцам соответствовать возлагаемой на них функции. Так путём изменения количества саркомеров и миофибрилл обеспечивается их функциональная реорганизация [1].
Работа мышц проявляется их сокращением, которое начинается с появления очага возбуждения на нейромышечных окончаниях. Наружная мембрана деполяризуется, открываются кальциевые каналы, и концентрация кальция внутри клетки возрастает. Ионы кальция связываются с тропонином, при этом конформируется тропониновый комплекс. Участки цепей миозина связываются с актином, что сопровождается высвобождением энергии вследствие расщепления АТФ до АДФ и остатка фосфорной кислоты. Угол между лёгкой и тяжёлой цепями миозина изменяется и актиновый филамент перемещается к центру саркомера, что приводит к изменению длины мышцы, её сокращению [1, 2].
Клетки скелетных мышц подразделяются на два типа:
А) Миосателлиты – взрослые стволовые клетки мышечной ткани. Представляют собой основу для обновления мышц и прироста их массы;
Б) Миосимпласты – формируют многоядерный синцитий. Сами по себе являются мышечными тубами с миофибриллами внутри, по периферии которых располагаются ядра.
Нагрузки, оказываемые на мышцы, и само мышечное сокращение имеют некую зависимость. Предполагается, что первое будет напрямую соответствовать второму. Это достигается за счёт усиления экспрессии генов сократительных белков и энзимов обменных процессов. Мышечная активность сопровождается количественными и качественными изменениями в миоцитах того типа, которые необходимы для наиболее эффективного осуществления выполняемой работы [2].
Мышечные волокна делятся на медленные (I тип) и быстрые (II тип). Оба этих типа имеют различный состав, включающий в себя сократительные белки, ферменты энергетического обмена и внутриклеточный кальций.
Увеличение силы мышц проявляется структурными перестройками, которые затрагивают нервную и мышечные системы. Изменения в нервной системе проявляются трансформацией величины кортикальных полей, которые регулируют выполнение определённого вида движения, влиянием на синхронизацию моторных единиц и на обучение определенных мышц, отвечающих за выполнение данного вида движений. Таким образом, наибольшая активность мышц наблюдается именно тогда, когда она необходима для достижения максимального эффекта (активность мышц агонистов при одновременной пассивности антагонистов). Также наблюдается изменение частоты и устойчивости генерируемых импульсов и порога возбудимости мотонейронов. Изменения в мышечной системе могут быть связаны с гипертрофией скелетных мышц (увеличение размеров мышечного волокна) и с их гиперплазией (увеличение количества миоцитов) [3].
Но прежде чем переходить к последним двум процессам, необходимо разобраться с изменениями, происходящими в самих мышцах. В момент выполнения работы миоцит подвергается действию физических и гуморальных факторов (пассивные механические силы, гипоксемия, факторы роста, и т.д.). Они являются причиной запуска путей передачи сигнала внутри клеток, опосредуя транскрипцию и трансляцию генов, ответственных за синтез белков [2]. Изменения данных путей сопровождаются реорганизацией мышечных волокон, точнее их типов.
Одним из основных исходных сигналов является повышенная концентрация кальция внутри клетки и кальцинейрина. Кальцинейрин дефосфорилирует факторы транскрипции – NFAT (nuclear factor of activated T-cells), которые находятся в фосфорилированном состоянии [4]. Данные факторы в дефосфорилированной форме активируют гены-мишени, что способствует перестроению быстрых волокон в медленные.
По мере приспособления мышц к нагрузкам изменяются и процессы метаболизма в них. Существуют различные параметры, влияющие на формирование адаптивных механизмов в миоцитах при выполнении работы. Важнейшим является гипоксия, которая, в свою очередь активирует ферментные системы (фумараза, цитратсинтаза, ЛДГ) и запускает работу факторов транскрипции (PGC1). При недостатке кислорода происходит активация одной изоформы семейства гипоксия-индуцированных факторов (HIF; hypoxia inducible factor), которая проникает в ядро, связывается с определенным участком ДНК и активирует гены, отвечающие за гликолиз, потребление кислорода и ангиогенез, увеличивая данные процессы. Некоторые гормоны также способны влиять на экспрессию генов в мышечных клетках. Это такие гормоны, как инсулин, гормон роста, которые вместе с кортизолом запускают катаболические реакции в условиях метаболического и энергетического истощения [3].
Стоит напомнить, что мышцы не являются постоянными клетками, а заменяются в течение жизни. Пролиферация необходима для предотвращения апоптоза клеток (регулируемый процесс клеточной гибели) и поддержания массы скелетных мышц. Это осуществляется через динамический баланс между синтезом белков в мышцах и их распадом. Мышечная гипертрофия возникает тогда, когда синтез белков превышает их распад.
Что же наблюдается при гипертрофии и гиперплазии мышечного волокна? При растяжении и сокращении мышц происходит образование факторов роста IGF и MGF, которые могут действовать как паракринно, так и аутокринно. С одной стороны, их действие проявляется в увеличении синтеза сократительных белков мышечных волокон. Основным участником данного механизма является фосфорилированная PKB [5]. Её активация начинается с влияния на мышцу нагрузки, которая приводит к синтезу гена, запускающего путь IGF/PI3K. В ткани имеется несколько изоформ, некоторые из них (IGF-1 и MGF), взаимодействуя с рецепторами приводят к конформационным изменениям. Через фосфорилирование ряда рецепторов и происходит активация PKB, способствующая развитию анаболических реакций [6].
С другой же стороны, происходит усиление пролиферации миосателлитов, их митотическая активность приводит к формированию новых клеток, а также сопровождается слиянием их с имеющимися мышечными волокнами или даёт возможность формировать новые. Миосателлиты расположены между базальной мембраной и сарколеммой. Покоящиеся клетки активируются непосредственно травмированием мышцы и в ответ на это начинают активно делиться и соединяться с частями поврежденного волокна. Под влиянием тяжёлой изнурительной работы происходит также активация данных клеток из-за образования многочисленных микротравм мышечного волокна. Вследствие этого наблюдается явление подобное процессам, происходящим при воспалении. В зону повреждения активно мигрируют нейтрофилы и макрофаги, которые активируют синтез ранее упомянутых факторов роста, регулирующих пролиферацию и дифференцировку миосателлитов. Мышечная гипертрофия отличается от мышечной гиперплазии. При гипертрофии мышц, увеличиваются сократительные элементы, и межклеточный матрикс расширяется для поддержки роста. Гиперплазия приводит к увеличению количества мышечных волокон. Гипертрофия сократительных элементов может происходить путем добавления саркомеров либо последовательно или параллельно.
В отечественной литературе не утихают споры о патогенетических аспектах мышечного роста. Чаще всего гипертрофию скелетных мышц человека рассматривают как их долговременную адаптацию к физическим нагрузкам различной направленности. Но существует понятие о кратковременной гипертрофии скелетных мышц – то есть изменение объема мышцы в результате одной силовой тренировки. Спортсмены, выступающие в соревнованиях по бодибилдингу или бодифитнесу хорошо знают, что объем мышц можно немного увеличить за счет собственной крови и осмотического давления, если использовать специальный метод тренировки – пампинг.
Неоспоримым является факт увеличения объёма мышечных волокон. Это так называемая миофибриллярная гипертрофия, при которой происходит изменение объёма миофибрилл и плотность их укладки. Механизм связан с увеличением количества саркомеров в миофибриллах. Значительная роль при этом отводится активированным клеткам-сателлитам. Миогенные стволовые клетки начинают пролифелировать, а затем сливаются с существующими клетками или взаимодействуют между собой для формирования новых мышечных волокон. Этот механизм актуален при восстановлении травмированных клеток и при спортивной гипертрофии.
Существует множество данных, доказывающих идущий параллельно с этим процесс увеличения объёма несократительной части мышцы – саркоплазматическая гипертрофия. Это тонкие перестройки на биохимическом уровне клетки, а так же увеличение количества митохондрий. Многие авторы считают, что трансформации в саркоплазме повышают выносливость мышц. Ряд исследователей утверждает, что увеличение различных неконтрактильных элементов и жидкости действительно может привести к приросту мышечной массы, но без сопутствующего увеличения силы. Саркоплазматическая гипертрофия достигается специальными тренировками и часто описывается как нефункциональная. Однако ряд специалистов предполагают, что отек мышечных волокон вызывает увеличение синтеза белка и таким образом способствует росту сократительной ткани.
Эти процессы редко бывают сбалансированными и зависят от характера и интенсивности нагрузки. В скелетных мышцах при этом синтез мышечных белков преобладает над их распадом. Причиной такого метаболизма сторонники гипотезы ацидоза считают накопление молочной кислоты. С точки зрения другой теории – временная гипоксия запускает реперфузию мышц и активирует деление клеток-сателлитов. Последнее время широкое распространение получила гипотеза механического повреждения мышечных волокон. Микроразрывы сократительных белков и повреждения саркоплазмы сопровождается увеличением концентрации ионов кальция, что и стимулирует пролиферацию сателлитов.
Из этого следует, что механизмы мышечной гипертрофии известны и неоспоримы. Очень дискутабельным остается вопрос о наличии процесса гиперплазии мышц. Большинство авторов сходится во мнении, что увеличение количества мышечных волокон у человека не доказано, но при этом описывается возможность получения гиперплазии мышц в экспериментальных условиях у животных (млекопитающих и птиц). Некоторые исследователи допускают частичное увеличения числа волокон. На основании проведенного мета-анализа экспериментальных работ отмечено, что количество мышечных элементов увеличилось в экспериментах на птицах значительнее, чем при использовании в качестве подопытных млекопитающих. Примечательно также, что эффект гиперплазии наблюдался там, где использовались постоянные растяжения, а не упражнения, сочетающие его с расслаблением. Ряд исследователей (Kraemer, William J. и MacDougall J.) утверждают, что этот механизм может осуществляться под влиянием силовых тренировок. Однако доказательств увеличения мышечных волокон у людей недостаточно. Длительных исследований (более года) добровольцев и спортсменов не проводилось. Высказывается мнение, что это слишком короткий период для этого процесса. Гиперплазия подтверждается в биопсийном материале, а погрешность этого метода составляет около 10 %, что делает результат очень сомнительным.
Общее число волокон предопределяется генетически и практически не меняется в течение жизни без применения специальных стимуляторов. Российские ученые подтверждают, что вклад гиперплазии в процесс увеличения объема мышц составляет не более 5 % и, как правило, потенцирован использованием анаболических стероидов. Также гиперплазию могут вызывать блокаторы миостатина. Гормон роста при этом не вызывает гиперплазии.
Таким образом, при мышечной работе происходит множество процессов на разных уровнях. Начиная с изменений интенсивности обменных процессов и заканчивая изменениями механизмов нервной и гуморальной регуляции. Реорганизация мышц, лежащая в основе этих процессов, приводит к изменению многочисленных характеристик деятельности спортсменов.
Проанализировав все данные и изучив все возможные гипотезы, становится очевидным, что в увеличении мышечных волокон играют некую роль всё-таки два процесса. Первый – гипертрофия с ёе подвидами для сократительной и несократительной части мышцы (миофибриллярная и саркоплазматическая), которая, по мнению многих исследователей, занимает основополагающую роль. И второй это гиперплазия с её минимальным, но существенным вкладом.
Вопрос 1. Почему в начале тренировок происходит значительное улучшение спортивных результатов, а потом они нарастают медленнее?
На начальных этапах значительное улучшение спортивных результатов происходит за счет того, что увеличивается число двигательных единиц, которые единовременно включаются в действие. Позднее результаты нарастают гораздо медленнее, так как они определяются уже перестройкой самих мышечных волокон, в которых увеличивается число сократительных нитей и митохондрий, но не меняется число самих волокон и их ядер (тренировочный эффект).
Вопрос 2. За счет чего при тренировках возрастает точность мышечных сокращений?
Возрастание точности мышечных сокращений при тренировках связано с адаптацией к выполняемым физическим упражнениям.
Вопрос 3. Поясните различие между динамической и статической работой.
Работа, связанная с перемещением тела или определенного груза, называется динамической (бег, ходьба, колка дров).
Работа, связанная с удержанием определенной позы или груза, называется статической (держать книгу в руках во время чтения).
Читайте также: