
В результате каких процессов возникают новообразования при скрещивании кратко
Обновлено: 05.07.2024
Иногда на формирование признака влияют две или несколько пар неаллельных генов. Проявление признака в этом случае зависит от характера их взаимодействия в процессе развития организма. В первом поколении появляется новый признак, которого не было у исходных родительских форм, и соотношение фенотипов во втором поколении будет иным. Рассмотрим несколько примеров различных типов взаимодействия генов.
Новообразование.Новообразованием называется такой тип взаимодействия генов, когда при их сочетании в одном организме развивается совершенно новая форма признака. Известно,, что у кур гены розовидного и стручковидного гребня не являются аллельными. И стручковидный, и розовидный гребень доминирует над листовидным. При скрещивании кур породы виан* дот, имеющих розовидный гребень (RRcc), с петухами породы брама со стручковидным гребнем (rrCQ у потомков первого поколения (RrCc) в результате взаимодействия двух доминантных генов R и С развивается новая форма гребня — ореховидная (рис. 11). Скрещивание потомков Fi между собой ведет к получению в F2 четырех разных фенотипов в соотношении: 9 — с генами R и С с гребнем ореховидной формы, 3Rcc — с гребнем розовидной формы, ЪггС— со стручковидным гребнем и \rrcc
пигмента в результате включения в генотип птиц Fi обоих доминантных генов — С (обусловливающего синтез тирозина) и О (обусловливающего синтез фермента). Во втором поколении окрашенных птиц ожидается 9 частей (СО..), а белых — 7 частей (С.оо—3, ccO..—З и ссоо—1).
Белые Белые
минорки шелковистые
Р J ССоо х (fccOO
| СО | Со | сО | со | |
| со | ССОО окраш. | ССОо окраш. | СсОО окраш. | СсОо окраш. |
| Со | ССОо окраш. | ССоо | СсОо окраш. | Ссоо Ш1 |
| сО | СсОО окраш. | СсОо окраш. | ссОО [бел\ | ссОо \Ш\ |
| со | СсОо окраш. | Ссоо | ссОо | ССОО |
Комплементарным взаимодействием генов обусловлен, очевидно, особый тип паралича задних конечностей у помесных собак, полученных от скрещивания датского дога с сенбернаром. Генетический анализ, проведенный Стокардом, показал, что при чистопородном разведении как у датских догов, так и у сенбернаров паралич не наблюдается. В то же время из 57 помесей Fi, полученных от реципрокных скрещиваний указанных пород и доживших до 3-месячного возраста, только 3 или 4 не имели такого дефекта. Среди помесей F2 из 66 щенков, доживших до 3 мес, была парализована почти треть. Болезнь проявляется внезапно в возрасте около 3 мес. Тяжесть болезни может быть различной: от слабой парализованности до полной утраты способности к самостоятельному передвижению. Аналогичное заболевание встречается у некоторых помесных собак-ищеек.

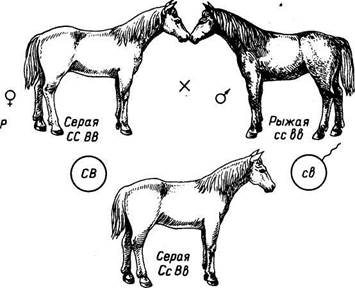
Эпистаз.При этом типе взаимодействия доминантный ген одной пары аллелей, например ген С, подавляет действие другого неаллельного доминантного гена В. При генотипе ССВВ проявляются признаки, обусловленные геном С. Ген, подавляющий развитие другого признака, называется эпистатичным, а подавляемый — гипостатичным. Например, у лошади серая доминирующая масть, связанная с ранним поседением, перекрывает все другие масти. При скрещивании серой лошади генотипа ССВВ с
масти. При скрещивании серой лошади генотипа ССВВ с рыжей генотипа ссЪЪ в Fi все потомки будут серыми с генотипом СсВЪ. При скрещивании Fi между собой в F2 наблюдается расщепление по фенотипу: 12 серых, 3 вороные и 1 рыжая. Аллель серой масти (С) перекрывает действие других независимых генов окраски. Все лошади, имеющие в генотипе аллель С, будут серыми. Если аллель С отсутствует, при наличии в генотипе аллеля В лошадь будет вороной (ссВВ, ссВЬ), и лошадь с генотипом ccbb, двойным рецессивом будет рыжей окраски (рис. 12).
Полимерия.При полимерии, или полимерном (полигенном) наследовании, на один и тот же признак влияют несколько разных, но сходно действующих неаллельных генов. Каждый из них усиливает развитие признака. Такие однозначно действующие гены называются аддитивными. Впервые этот тип взаимодействия генов установлен Нильсоном-Эле при изучении наследования окраски чешуи овса и зерен пшеницы.
Рассмотрим пример наследования окраски зерен пшеницы при взаимодействии двух пар полимерных генов. Различают две основные окраски зерен: красную и белую. Полимерные гены, действующие на один и тот же признак, обозначают одинаковой буквой. Разные аллельные пары обозначают цифрами внизу букв. Исходя из этого, генотип пшеницы с темно-красным зерном будет А1А1А2А2, с белым зерном — ата&г- У первого родителя образуются гаметы А1А2, у второго — в/д* В результате потомки Fi будут иметь генотип Aiayi^ (двойная гетерозигота) и промежуточную окраску зерен — светло-красную, так как имеется два доминантных гена (Ai и Аг), влияющих на проявление признака. Потомки первого поколения образуют по четыре сорта гамет и при спаривании между собой дадут F2, в котором расщепление по фенотипу и генотипу будет таким: из 16 частей 1 часть темно-красных (А1А1А2А2), четыре красных QA1A1A2CI2, lAiatAiAi), 6 светло-красных (4AiOiA2O2, \AiA1a2a2, ХаыААг), 4 бледно-красных (2Aiaia2O2, 20101X202) и 1 часть белых (ауД/ДгДг). В этом легко убедиться, составив решетку Пеннета. Как видим, степень развития окраски зависит от количества доминантных генов, влияющих на формирование этого признака. При отсутствии доминантных генов окраска зерна пшеницы белая.
Полимерный тип взаимодействия генов имеет большое значение для понимания наследования количественных признаков. Эти признаки не обладают фенотипической дискретностью, и их невозможно распределить по четким фенотипическим классам. Их оценивают с помощью количественных методов учета. К количественным относятся признаки, характеризующие продуктивность животных: удой за лактацию, масса животного, настриг шерсти, масса яйца. В некоторых случаях полигенно наследуется резистентность к неблагоприятным условиям внешней среды.
 |
| Гаметы |
| Св |
| сВ |
| СВ |
 |
| СВ |
сВ
| Серая ССВв |
| Серая СсВВ |
| ссвв |
Серая СсВВ
| Св |
| Серая СсВВ |
| Серая |
| Серая Сс88 |
 |

Серая ССВВ
| ■!■. |
| СВ |
.-.-ж
| Серая СсВВ |
 |
вороная ее ев

сВ
Вороная се Вв
Рис. 12. Эпистаз при наследовании серой, вороной и рыжей масти у лошадей
Все эти признаки формируются под влиянием многих генов, каждый из которых усиливает развитие признака.
Гены-модификаторы.Гены, не проявляющие собственного действия, но усиливающие или ослабляющие эффект действия других генов, называются генами-модификаторами. Гены-модификаторы играют, по-видимому, определенную роль в формировании у животных резистентное™ к инфекционным болезням. Например, скот герефордской породы имеет белую голову, и при пастбищном содержании в условиях сильной солнечной инсоляции животные с непигментированными и слабопигментирован-ными веками болеют раком глаз. При усилении пигментации век частота заболевания уменьшается, а при интенсивной пигментации в тех же условиях болезнь не возникает. Оказалось, что интенсивность пигментации кожи вокруг глаз у белоголовых животных наследственна. Это говорит о существовании генов — модификаторов основного гена, обусловливающего белую окраску головы. Таким образом, путем селекции можно избавиться от заболевания глаз раком.
Пенетрантность гена — это доля особей, у которых проявляется ожидаемый фенотип. При полной пенетрантности (100 %) мутантный ген проявляет свое действие у каждой особи. При неполной пенетрантности (меньше 100 %) ген проявляется фе-нотипически не у всех особей. Экспрессивность и пенетрантность гена в значительной степени зависят, по-видимому, от влияния генов-модификаторов и условий развития особей.
Плейотропия.Это влияние одного гена на развитие двух и более признаков (множественное действие гена). Так, Д. К. Беляев и А. И. Железнова установили, что у норок большинство мутаций, сопровождающихся изменением окраски волосяного покрова, рецессивно и в силу плейотропии при этом снижаются плодовитость и жизнеспособность животных.
Явление плейотропии объясняется тем, что гены плейотроп-ного действия контролируют синтез ферментов, которые участвуют в многочисленных обменных процессах в клетке и в организме в целом и тем самым одновременно влияют на проявление и развитие других признаков.
На основании рассмотренного действия генов-модификаторов, взаимодействия генов и плейотропного действия генов можно видеть, что формирование признака — очень сложное яв-
Иногда на формирование признака влияют две или несколько пар неаллельных генов. Проявление признака в этом случае зависит от характера их взаимодействия в процессе развития организма. В первом поколении появляется новый признак, которого не было у исходных родительских форм, и соотношение фенотипов во втором поколении будет иным. Рассмотрим несколько примеров различных типов взаимодействия генов.
Новообразование.Новообразованием называется такой тип взаимодействия генов, когда при их сочетании в одном организме развивается совершенно новая форма признака. Известно,, что у кур гены розовидного и стручковидного гребня не являются аллельными. И стручковидный, и розовидный гребень доминирует над листовидным. При скрещивании кур породы виан* дот, имеющих розовидный гребень (RRcc), с петухами породы брама со стручковидным гребнем (rrCQ у потомков первого поколения (RrCc) в результате взаимодействия двух доминантных генов R и С развивается новая форма гребня — ореховидная (рис. 11). Скрещивание потомков Fi между собой ведет к получению в F2 четырех разных фенотипов в соотношении: 9 — с генами R и С с гребнем ореховидной формы, 3Rcc — с гребнем розовидной формы, ЪггС— со стручковидным гребнем и \rrcc
Кроме наследования в виде доминантности и рецессивности генов, может иметь место совместное воздействие разных аллелей данного локуса: кодоминантное действие генов. Например, синтез белка гемоглобина обусловлен генами А и В, которые дают гемоглобин трех типов АА, ВВ и АВ, и каждый из генотипов обеспечивает синтез нормальных гемоглобинов. Проявляются различия совместного действия А и В только в биохимической структуре молекулы соответствующими методами путем электрофореза образцов крови.
Кроме отмеченных закономерностей в наследовании признаков потомства, обусловленных взаимодействием аллелей одного локуса, наблюдается такая особенность, как появление нового состояния признака у потомство", которое отсутствовало у его родителей. Этот тип наследования называется "новообразованием при скрещивании". Примером такого наследования служит скрещивание кофейного (коричневого) добермана с голубым доберманом. В результате их скрещивания получаются доберманы черного цвета, так как у исходных типов доберманов различны аллели генотипа локуса. Генотип кофейного добермана включает ген D, определяющий интенсивность окраски и ген b, как рецессивный аллель гена черного цвета. Поэтому кофейный доберман имеет генотип bbDD. Генотип голубого добермана включает доминантный ген черной окраски B, но эта черная окраска не может полностью проявиться из-за отсутствия гена D (усилителя). В результате получается голубой доберман с генотипом BBdd. При скрещивании доберманов обоих типов bbDD x BBdd их потомство будет иметь гетерозиготный генотип BbDd, а по окраске все потомство будет черного цвета. От скрещивания таких гетерозиготных особей будет происходить расщепление по фенотипам в соотношении: черные 9BD+голубые 3Bd и кофейные разных оттенков 3bD+lbd, т. е. соотношение, как при обычном дигибридном скрещивании 9: 3: 3: 1.
Взаимодействие неаллельных генов (находящихся в разных участках хромосом) также приводит к новообразованию: комплементарному взаимодействию генов. При таком типе наследования расщепление по фенотипам во втором поколении будет отмечаться по их соотношению от вышеописанных. Например, в F2 может быть соотношение 9: 7 или 9: 3: 4, 12: 3: 1 при наличии разных аллелей в локусе A и локусе E. У собак соотношение 9: 7 прослежено при скрещивании гетерозиготных черных собак (генотип AsAYEe) между собой. Фенотипы их потомства были следующие: 9 черных и 7 желтых. Скрещивание черных гетерозиготных собак с генотипом As'a'Ee дает расщепление в потомстве следующего типа: 9 черных, 3 желтых, 4 рыжевато-коричневых. А при наличии других аллелей у черных гетерозиготных собак с генотипом AsAYEbrE соотношение фенотипов с новообразованием будет еще более отличающимся, а именно: 12 черных, 3 полосато-тигровых (DYDYEbrE) и 1 желтая (DYDYEE).
Генотип - это система взаимодействующих генов, а не механическая их совокупность. В результате взаимодействия генов в потомстве могут возникать новые признаки, отсутствовавшие у родительских организмов. Это явление называют новообразованием при скрещиваниях. Оно очень часто наблюдается в природе. Например, формирование такого, казалось бы, элементарного признака, как окраска цветков, зависит от взаимодействия, по крайней мере, двух неаллельных генов, продукты которых взаимно дополняют друг друга.
В рассмотренных выше примерах имело место относительно независимое проявление действия генов. Доминантный ген желтой окраски семян гороха вызывает развитие этого признака как в присутствии доминантного гена гладкой формы семян, так ипри наличии аллельного ему рецессивного гена морщинистой формы семян. У морских свинок ген черной или белой окраски действует независимо от генов, определяющих характер развития шерстного покрова. На основании знакомства с этими примерами может сложиться впечатление, что генотип организма слагается из суммы отдельных, независимо действующих генов. Такое представление ложно. Хотя в некоторых случаях действие генов, принадлежащих к разным аллелям, относительно независимо, но чаще между ними осуществляются разные формы взаимодействия.
Развитие того или иного признака организма обычно находится под контролем многих генов.
У разных пород кур имеются разнообразные формы гребня. Четыре типа формы гребня: гороховидный, розовидный, ореховидный и простой. При скрещивании между собой гомозиготных птиц с розовидным и простым гребнем признак розовидного гребня оказывается доминантным. Первое поколение гибридов будет иметь розовидный гребень, а в F2 произойдет расщепление в отношении 3: 1 (три розовидных, один простой).
Аналогичный результат получается при скрещивании птиц сгороховидным и простым гребнем. В F1 будет доминировать гороховидный гребень, а в F2 происходит расщепление в отношении 3 гороховидных, 1 простой. Ну, а как будут выглядеть гибриды, если скрестить между собой птиц с гороховидным и розовидным гребнем, т. е. с двумя разными, неаллельными доминантными признаками? Опыт показывает, что при этом все потомство в F1 будет иметь совершенно новую форму гребня — ореховидную. При скрещивании этих гибридов между собой расщепление в F2 пойдет по дигибридной схеме, а именно: 9 ореховидных, 3 розовидных,. 3 гороховидных, 1 простой. Если мы сопоставим этот результат с уже известным нам ходом дигибридного расщепления у горохов и морских свинок, то придем к заключению, что ореховидный гребень развивается в том случае, когда в зиготе присутствуют одновременно два доминантных гена. Следовательно, простой гребень — результат взаимодействия двух рецессивных генов. Обозначим аллель, доминантный ген которой вызывает развитие гороховидного гребня, латинскими буквами Р—р, а аллель роговидного гребня как R—г.
Множественное действие генов.На рассмотренных только что примерах было показано, что большинство наследственно обусловленных признаков организма находится под контролем не одного, а многих генов. Наряду с этим имеет место и другое явление. Во многих случаях ген оказывает свое действие не на один, а на ряд признаков организма. При этом особенно отчетливо выступает одна сторона действия гена, по которой его обычно и называют. Приведем примеры. У большинства растений с красными цветками (наследственный признак) в стеблях (особенно в междоузлиях) тоже имеется красный пигмент. У растений с белыми цветками стебли чисто зеленые. У растения водосбора ген, обусловливающий красную окраску цветка, имеет множественное действие. Он определяет фиолетовый оттенок листьев, удлинение стебля и больший вес семян. Множество аналогичных примеров можно привести и из животного мира. Ограничимся только одним. У излюбленного объекта генетических исследований — плодовой мушки дрозофилы, генотип которой изучен очень полно, ген, определяющий отсутствие пигмента в глазах, вместе с тем снижает плодовитость, влияет на окраску некоторых внутренних органов и уменьшает продолжительность жизни.
Накопившийся в настоящее время в генетике обширный материал по изучению наследственности у самых различных растений, животных, микроорганизмов говорит о том, что множественное действие гена — широко распространенное явление.
Приведенные факты и наблюдения, касающиеся взаимодействия генов и их множественного действия, позволяют нам значительно углубить общее представление о природе наследственной основы организмов — генотипе. Факт расщепления в потомстве гибридов позволяет утверждать, что генотип слагается из отдельных элементов, которые называются генами и могут отделяться друг от друга и наследоваться независимо (вспомним второй закон Менделя). Наряду с прерывистым характером генотип обладает целостностью и не может рассматриваться как простая механическая сумма отдельных генов. Эта целостность генотипа выражается, прежде всего, втом, что отдельные компоненты его (гены) находятся в тесном взаимодействии друг с другом. Развитие признаков организма определяется взаимодействием множества генов. С другой стороны, каждый ген обладает множественным действием, оказывая влияние на развитие не одного, а многих признаков организма. Генотип организма связан с определенными структурами клетки, с ее хромосомным аппаратом.
Генетика пола
Хорошо известно, что у раздельнополых организмов (в том числе и у человека) соотношение полов обычно составляет 1:1. Какие причины определяют пол развивающегося организма? Вопрос этот издавна интересовал человечество ввиду его большого теоретического и практического значения. Хромосомный комплекс самцов и самок у большинства раздельнополых организмов неодинаков. Познакомимся с этими различиями на примере набора хромосом у дрозофилы. Хромосомные комплексы самца и самки, они не вполне одинаковы. По трем парам хромосом самцы и самки не отличаются друг от друга. Но в отношении одной пары имеются существенные различия. У самки присутствуют две одинаковые (парные) палочковидные хромосомы, у самца же имеется только одна такая хромосома, пару которой составляет особая двуплечая хромосома. Те хромосомы, в отношении которых между самцами и самками нет различий, носят название аутосом. Хромосомы, по которым самцы и самки отличаются друг от друга, называются половыми. Таким образом, хромосомный комплекс дрозофилы слагается из шести аутосом и двух половых хромосом. Половая палочковидная хромосома, присутствующая у самки в двойном числе, а у самца в единичном, называется Х-хромосомой; вторая половая (двуплечая) хромосома самца
Каким же образом рассмотренные половые различия в хромосомных комплексах самцов и самок поддерживаются в процессе размножения? Для ответа на этот вопрос необходимо выяснить поведение хромосом в мейозе и при оплодотворении. При созревании половых клеток у самки каждая яйцеклетка в результате мейоза получает гаплоидный комплект из четырех хромосом, в том числе по одной Х-хромосоме. При мейозе у самца образуются сперматозоиды двух сортов. Во всех сперматозоидах имеется по 3 аутосомы. Половые хромосомы расходятся к противоположным полюсам — веретенам. Таким образом, Х-хромосома отходит к одному полюсу, а Y-хромосома — к противоположному. Благодаря этому у самцов в равных количествах образуются сперматозоиды двух сортов. Одни несут 3 аутосомы и Х-хромосому, другие — 3 аутосомы и Y-хромосому. При оплодотворении равновероятны 2 комбинации. Яйцеклетка может быть оплодотворена спермием с Х- или Y-хромосомой. В первом случае из оплодотворенного яйца разовьется самка, во втором — самец. Таким образом, пол организма определяется в момент оплодотворения и зависит от хромосомного комплекса зиготы.
У человека хромосомный механизм определения пола тот же, что и у дрозофилы. Диплоидное число хромосом человека — 46. В это число входят 22 пары аутосом (одинаковые у мужчин и женщин) и 2 половые хромосомы. У женщин это две Х-хромосомы, у мужчин — одна Х- и одна Y-хромосома. Соответственно у мужчин образуются сперматозоиды двух сортов с Х- и Y-хромосомами.
У некоторых раздельнополых организмов (например, насекомых) Y-хромосома вообще отсутствует. В этих случаях у самца оказывается на одну хромосому меньше (вместо Х- и Y- у него имеется одна Х-хромосома). Тогда при образовании мужских гамет в процессе мейоза Х-хромосома не имеет партнера для конъюгации и отходит в одну из клеток. В результате половина всех сперматозоидов имеет Х-хромосому, а другая половина лишена ее. При оплодотворении яйца спермием с Х-хромосомой получается комплекс с двумя Х-хромосомами, и из такого яйца развивается самка. Если яйцеклетка будет оплодотворена спермием без Х-хромосомы, то разовьется организм с одной Х-хромосомой (полученной через яйцеклетку от самки), который будет самцом.
Во всех рассмотренных выше примерах развиваются спермии двух категорий: либо с Х- и Y-хромосомами (дрозофила, человек), либо половина спермиев несет Х-хромосому, а другая совсем лишена половой хромосомы. Яйцеклетки же в отношении половых хромосом все одинаковы. Во всех этих случаях мы имеем мужскую гетерогаметность (разногаметность). Женский же пол гомогаметен (равногаметен). Наряду с этим в природе встречается и другой тип определения пола, характеризующийся женской гетерогаметностью. Здесь имеют место отношения как раз обратные только что рассмотренным. Разные половые хромосомы или только одна Х-хромосома свойственны женскому полу. Мужской пол обладает парой одинаковых Х-хромосом. Очевидно, что в этих случаях будет иметь место женская гетерогаметность. После мейоза образуются яйцевые клетки двух сортов, тогда как в отношении хромосомного комплекса все спермин одинаковы (все несут одну Х-хромосому). Следовательно, пол зародыша будет определяться тем, какое яйцо (с Х- или Y-хромосомой) будет оплодотворено.
Женская гетерогаметность имеет место у некоторых насекомых, например у бабочек. Среди позвоночных животных она характерна для птиц и пресмыкающихся.
Сцепленная с полом наследственность. Врассмотренных выше различных примерах хода расщепления мы не учитывали, от кого гибрид получает тот или иной ген — от самца или от самки. Никаких различий в этом отношении не наблюдается. Это вполне понятно, ибо гаметы, как мужские, так и женские, несут гаплоидные комплексы хромосом, которые, объединяясь при оплодотворении, дают диплоидный комплекс зиготы. Сказанное вполне справедливо в отношении аутосом. В отношении же половых хромосом дело, как мы только что видели, обстоит иначе. Гетерогаметный пол (у человека, например, мужской пол) получает Х-хромосому только от гомогаметного пола, и она присутствует у него в единственном числе. Например, все мужчины имеют только материнскую Х-хромосому, тогда как женщины получают по одной Х-хромосоме и от отца и от матери. Очевидно, что в отношении генов, локализующихся в Х-хромосомах, наследование будет осуществляться несколько иначе, чем в отношении генов, локализованных в аутосомах. Рассмотрим пример, поясняющий особенности наследования через хромосому. Человеку свойственно тяжелое наследственное заболевание — гемофилия, одним из проявлений которой является несвертываемость крови. Болеют гемофилией только мужчины. Гемофилия возникла как рецессивное наследственное изменение в Х-хромосоме. Обычно носителями гемофилии являются внешне совершенно здоровые (по фенотипу) женщины, одна из Х-хромосом которых несет ген гемофилии. Однако фенотипически он не проявляется, так как подавляется аллельным ему доминантным геном, обусловливающим нормальную свертываемость крови. Такая женщина-носительница передает гемофилию половине своих сыновей — тем, кому попадает Х-хромо-сома, несущая рецессивный ген гемофилии. Так как у мужчины Х-хромосо-ма присутствует в единственном числе у доминантного аллельного гена, доминирующего над геном гемофилии, гемофилия проявляется фенотипически. Вероятность заболевания гемофилией женщины ничтожно мала. Это может произойти в случае брака гетерозиготной по гемофилии женщины с больным гемофилией мужчиной. Можно говорить, что гемофилия — признак, сцепленный с полом. По такой же точно схеме у человека наследуется и дальтонизм (неспособность различать красный цвет).
Не вдаваясь в разбор других случаев сцепленной с полом наследственности, укажем, что таких признаков известно довольно много у домашних животных и птиц.
Список литературы
1. Азимов А. Краткая история биологии. М.,1997.
2. Кемп П., Армс К. Введение в биологию. М.,2000.
3. Либберт Э. Общая биология. М.,1978 Льоцци М. История физики. М.,2001.
4. Найдыш В.М. Концепции современного естествознания. Учебное пособие. М.,1999.
Читайте также:
- Какие спасательные службы обычно организуются в учреждениях и на предприятиях кратко
- Дмитрий самозванец анализ кратко
- Какими инструментами пользуются для измерения расстояний геометрия 7 класс ответы кратко
- Какие объекты базы данных microsoft access вы знаете кратко
- Покажите суть школьных проектов екатерининской эпохи таблица