
В чем значение биогенетического закона кратко
Обновлено: 03.07.2024
впервые сформулированное Фрицем Мюллером и Эрнстом Геккелем положение о том, что ряд форм, которые живое существо проходит в процессе своего индивидуального развития от яйцеклетки до развитого состояния (онтогенез), представляет собой краткое, сжатое, отличающееся многообразными изменениями повторение длинного ряда форм, пройденных предками данного организма или осн. формами данного вида с древних времен до настоящего времени (филогенез). Этого взгляда, раскритикованного еще К. Бэром, в настоящее время придерживаются лишь очень немногие.
Философский энциклопедический словарь . 2010 .
(от греч. βίος – жизнь и γένεσις – происхождение) – принимаемое эволюционистами-дарвинистами положение, согласно к-рому каждая органич. форма в своем индивидуальном развитии (онтогенезе) в нек-рой мере повторяет черты и особенности тех форм, от к-рых она произошла.Термин "Б. з." введен нем. биологом Э. Геккелем (1866), к-рый сформулировал "основной Б. з.": онтогенез, или развитие органич. индивидуумов как ряд изменений форм, через к-рые проходит каждый индивидуальный организм за время своего индивидуального существования, непосредственно обусловливается филогенезом, т. е. развитием органич. группы, возникшей от общего корня, к к-рому данный организм относится. По Геккелю, "онтогения представляет собой краткое и быстрое повторение (р е к а п и т у л я ц и ю) филогении, повторение, обусловленное физиологическими функциями н а с л е д с т в е н н о с т и (воспроизведения) и п р и с п о с о б л я е м о с т и (питания). В течение быстрого и краткого хода своего онтогенетического развития особь повторяет важнейшие из тех изменений формы, через которые прошли ее предки в течение медленного и длительного хода их палеонтологического развития по законам наследственности и приспособления" (Мюллер Ф. и Геккель Э., Основной биогенетический закон, М.–Л., 1940, с. 169). Геккель различал в индивидуальном развитии п а л и н г е н е з ы, к к-рым относил явления, соответствующие Б. з. и повторяющие те или иные особенности предковых форм, т. е. явления рекапитуляции (повторения), и ц е н о г е н е з ы, к-рые представляют нарушения рекапитуляции, т. е. отступление от повторения филогенеза. На основе палингенезов Геккель считал возможным восстанавливать некогда существовавшие формы, от к-рых происходит данный вид.
Б. з. в трактовке Геккеля был грубым упрощением биологич. фактов, хотя и содержал рациональное зерно. На основании данных биологии и эволюционной палеонтологии, или палеобиологии, можно говорить лишь об элементах рекапитуляции в индивидуальном развитии организмов; ни в одной стадии онтогенеза организм не представляет сколько-нибудь полного повторения предковых форм данного филогенетич. ряда. Основная идея Б. з. существовала задолго до Геккеля. О подобии между ступенями зародышевого развития организмов и классификационными рядами менее высокоорганизованных групп писали еще в конце 18 в. и в начале 19 в. как авторы, не имевшие ясного представления об эволюции органич. мира или придерживавшиеся весьма ограниченного понимания эволюции (напр., нем. натурфилософы Л. Окен и И. Меккель), так и нек-рые эволюционисты додарвиновских времен (напр., русский естествоиспытатель К. Ф. Рулье). Впервые проблему соотношения между онтогенезом и филогенезом, проблему рекапитуляции во всей ее полноте поставил Ч. Дарвин. Еще в "Очерке 1844 года", разрабатывая эволюц. теорию, Дарвин писал, что "зародыши ныне существующих позвоночных отражают строение некоторых взрослых форм этого большого класса, существовавших в более ранние периоды истории земли" (Соч., т. 3, 1939, с. 215). В "Происхождении видов. " основная идея теории рекапитуляции и Б. з. выражена Дарвином как закон сходства древних форм с эмбриональными стадиями новейших форм. Дарвин указал многие факты, свидетельствующие в пользу Б. з. (напр., наличие у зародышей млекопитающих и птиц жаберных щелей, у теленка – никогда не прорезывающихся зубов в верхней челюсти там, где у взрослого зубов нет и где они функционально замещаются другими органами). Однако Дарвин отмечал и такие факты, к-рые не укладываются в примитивное понимание Б. з., напр. явления гетерохронии (изменения во времени появления признаков) и, в частности, случаи, когда определенные признаки возникают в онтогенезе потомков раньше, чем в онтогенезе предковых форм. Изменения признаков могут иметь место, по Дарвину, на всех стадиях (яйцо, семя, молодой и взрослый организм); это он объяснял естественным отбором. Явления рекапитуляции рассматривал с дарвинистских позиций нем. ученый Ф. Мюллер (1864). Ф. Энгельс подчеркнул важное значение Б. з. для эволюционного учения.
Основная причина явлений рекапитуляции, подчиняющихся Б. з., заключается в том, что эволюция – не внезапная и одновременная полная перестройка всего организма, а процесс последовательного изменения с сохранением, до нек-рого историч. момента, многих основных черт, процесс, к-рый может привести к коренному преобразованию всего тела лишь через многочисленные относительно мелкие изменения, осуществление к-рых требует значительного времени. Многие черты строения и деятельности тела сохраняются на протяжении длинных филогенетич. рядов в виде рудиментарных органов или удерживаются только у юных особей; в обоих случаях они представляют собой прямое доказательство эволюции и того конкретного пути, к-рый она совершала в данной группе.
Убедительные примеры рекапитуляции дает изучение многих групп ископаемых животных – плеченогих, пластинчатожаберных и брюхоногих моллюсков, фораминифер и особенно аммоноидей (из класса головоногих моллюсков); в последнем случае рекапитуляция связана с тем, что молодь была близка к взрослым особям по образу жизни. Явления, соответствующие Б. з., имеют значение для систематики организмов, напр. в систематике животных для установления таксономич. положения нек-рых групп низших ракообразных (ракушковых, усоногих); эти явления отмечались в многочисленных группах животных (напр., рудиментарные коренные зубы у юных утконосов, впоследствии совершенно исчезающие; многочисленные зубы у зародышей беззубых китов, затем полностью атрофирующиеся). Рекапитуляция и тесно связанные с ней явления известны не только у животных, но и у растений, на что неоднократно указывал, напр., И. В. Мичурин. Так, рассматривая "влияние атавизма", он отмечал, что оно "является неустранимым во всех без исключения сеянцах различных видов и разновидностей растений только в начальных стадиях развития их из семени, выражаясь тем, что все они в молодости имеют в своем наружном виде кажущееся сходство с дикими формами родоначального вида" (Соч., т. 1, 1948, 185–86).
Нек-рые зоологи и палеонтологи совершенно отвергают Б. з., ссылаясь на то, что онтогенез никогда не представляет собой даже краткого повторения важнейших этапов филогенеза. Это возражение имеет силу по отношению к первоначальному геккелевскому определению "основного Б. з.", но не может быть выдвинуто против дарвиновского понимания теории рекапитуляции, согласно к-рому онтогенез отнюдь не представляет краткого повторения филогенеза, но на разных стадиях содержит элементы такого повторения. Столь же неосновательно то возражение против основной идеи Б. з., к-рое основывается на случаях, когда эмбриональные структуры, бесспорно соответствующие некогда существовавшим органам предков данной формы, несут какие-то определенные функции у зародышей, т. е. не являются полностью бесполезными: напр., появление в онтогенезе рыб жаберных щелей, обслуживающих у зародышей дыхательную систему, но в дальнейшем исчезающих, надо рассматривать как рекапитуляцию, так как эти щели соответствуют жаберным щелям предков современных рыб; зубы у зародышей беззубых китов имеют определенное значение в индивидуальном развитии этих животных, но это не опровергает того положения, что такие эмбриональные зубы соответствуют зубам предков, у к-рых они служили для захвата пищи. Нек-рые противники теории рекапитуляции утверждают, что в животном мире господствующее положение занимают процессы, как бы противоположные Б. з. Эти ученые утверждают, что эмбриональные структуры предков часто становятся структурами взрослых особей у потомков – у более поздних представителей данного филогенетич. ряда (нем. палеонтолог О. Шиндевольф, англ. зоолог-эмбриолог Г. де Бир). Крайние представители этого направления приходят к мысли, что эволюция шла в основном от высшего состояния к низшему. Такое понимание соотношения между онтогенезом и филогенезом направлено к дискредитации научной, материалистической эволюционной теории в целом.
Лит.: Энгельс Ф., Анти-Дюринг, М., 1957, с. 70; Дарвин Ч., Происхождение видов путем естественного отбора, Соч., т. 3, М.–Л.,1939,с. 627–36; Мичурин И. В., Принципы и методы работы, Соч., Т.1, 2 изд., М., 1948, с. 651; Мюллер Ф., Геккель Э., Основной биогенетический закон, Избр. работы, М.–Л. 1940.
Философская Энциклопедия. В 5-х т. — М.: Советская энциклопедия . Под редакцией Ф. В. Константинова . 1960—1970 .
Смотреть что такое БИОГЕНЕТИЧЕСКИЙ ЗАКОН в других словарях:
БИОГЕНЕТИЧЕСКИЙ ЗАКОН
термин, введенный в биологию Геккелем и означающий, что ряд форм, проходимый организмом при его индивидуальном развитии (онтогении), есть "краткое повт. смотреть
БИОГЕНЕТИЧЕСКИЙ ЗАКОН
закономерность в живой природе, сформулированная немецким учёным Э. Геккелем (1866) и состоящая в том, что индивидуальное развитие особи (Онтог. смотреть
БИОГЕНЕТИЧЕСКИЙ ЗАКОН
БИОГЕНЕТИЧЕСКИЙ ЗАКОН, закономерность в живой природе, сформулированная нем. учёным Э. Геккелем (1866) и состоящая в том, что индивидуальное развитие. смотреть
БИОГЕНЕТИЧЕСКИЙ ЗАКОН
Биогенетический закон — термин, введенный в биологию Геккелем и означающий, что ряд форм, проходимый организмом при его индивидуальном развитии (онтогении), есть "краткое повторение длинного ряда форм, через который прошли предки этого организма (племенные формы данного вида) от первоначального их появления на земле до настоящего времени". Таким образом, "онтогения есть краткое повторение филогении (т. е. племенной истории развития)". См. слова Онтогения и Филогения.
БИОГЕНЕТИЧЕСКИЙ ЗАКОН
от греч. . – жизнь и . – происхождение) – принимаемое эволюционистами-дарвинистами положение, согласно к-рому каждая органич. форма в своем индивидуальном развитии (онтогенезе) в нек-рой мере повторяет черты и особенности тех форм, от к-рых она произошла. Термин "Б. з." введен нем. биологом Э. Геккелем (1866), к-рый сформулировал "основной Б. з.": онтогенез, или развитие органич. индивидуумов как ряд изменений форм, через к-рые проходит каждый индивидуальный организм за время своего индивидуального существования, непосредственно обусловливается филогенезом, т. е. развитием органич. группы, возникшей от общего корня, к к-рому данный организм относится. По Геккелю, "онтогения представляет собой краткое и быстрое повторение (р е к а п и т у л я ц и ю) филогении, повторение, обусловленное физиологическими функциями н а с л е д с т в е н н о с т и (воспроизведения) и п р и с п о с о б л я е м о с т и (питания). В течение быстрого и краткого хода своего онтогенетического развития особь повторяет важнейшие из тех изменений формы, через которые прошли ее предки в течение медленного и длительного хода их палеонтологического развития по законам наследственности и приспособления" (Мюллер Ф. и Геккель Э., Основной биогенетический закон, М.–Л., 1940, с. 169). Геккель различал в индивидуальном развитии п а л и н г е н е з ы, к к-рым относил явления, соответствующие Б. з. и повторяющие те или иные особенности предковых форм, т. е. явления рекапитуляции (повторения), и ц е н о г е н е з ы, к-рые представляют нарушения рекапитуляции, т. е. отступление от повторения филогенеза. На основе палингенезов Геккель считал возможным восстанавливать некогда существовавшие формы, от к-рых происходит данный вид. Б. з. в трактовке Геккеля был грубым упрощением биологич. фактов, хотя и содержал рациональное зерно. На основании данных биологии и эволюционной палеонтологии, или палеобиологии, можно говорить лишь об элементах рекапитуляции в индивидуальном развитии организмов; ни в одной стадии онтогенеза организм не представляет сколько-нибудь полного повторения предковых форм данного филогенетич. ряда. Основная идея Б. з. существовала задолго до Геккеля. О подобии между ступенями зародышевого развития организмов и классификационными рядами менее высокоорганизованных групп писали еще в конце 18 в. и в начале 19 в. как авторы, не имевшие ясного представления об эволюции органич. мира или придерживавшиеся весьма ограниченного понимания эволюции (напр., нем. натурфилософы Л. Окен и И. Меккель), так и нек-рые эволюционисты додарвиновских времен (напр., русский естествоиспытатель К. Ф. Рулье). Впервые проблему соотношения между онтогенезом и филогенезом, проблему рекапитуляции во всей ее полноте поставил Ч. Дарвин. Еще в "Очерке 1844 года", разрабатывая эволюц. теорию, Дарвин писал, что "зародыши ныне существующих позвоночных отражают строение некоторых взрослых форм этого большого класса, существовавших в более ранние периоды истории земли" (Соч., т. 3, 1939, с. 215). В "Происхождении видов. " основная идея теории рекапитуляции и Б. з. выражена Дарвином как закон сходства древних форм с эмбриональными стадиями новейших форм. Дарвин указал многие факты, свидетельствующие в пользу Б. з. (напр., наличие у зародышей млекопитающих и птиц жаберных щелей, у теленка – никогда не прорезывающихся зубов в верхней челюсти там, где у взрослого зубов нет и где они функционально замещаются другими органами). Однако Дарвин отмечал и такие факты, к-рые не укладываются в примитивное понимание Б. з., напр. явления гетерохронии (изменения во времени появления признаков) и, в частности, случаи, когда определенные признаки возникают в онтогенезе потомков раньше, чем в онтогенезе предковых форм. Изменения признаков могут иметь место, по Дарвину, на всех стадиях (яйцо, семя, молодой и взрослый организм); это он объяснял естественным отбором. Явления рекапитуляции рассматривал с дарвинистских позиций нем. ученый Ф. Мюллер (1864). Ф. Энгельс подчеркнул важное значение Б. з. для эволюционного учения. Основная причина явлений рекапитуляции, подчиняющихся Б. з., заключается в том, что эволюция – не внезапная и одновременная полная перестройка всего организма, а процесс последовательного изменения с сохранением, до нек-рого историч. момента, многих основных черт, процесс, к-рый может привести к коренному преобразованию всего тела лишь через многочисленные относительно мелкие изменения, осуществление к-рых требует значительного времени. Многие черты строения и деятельности тела сохраняются на протяжении длинных филогенетич. рядов в виде рудиментарных органов или удерживаются только у юных особей; в обоих случаях они представляют собой прямое доказательство эволюции и того конкретного пути, к-рый она совершала в данной группе. Убедительные примеры рекапитуляции дает изучение многих групп ископаемых животных – плеченогих, пластинчатожаберных и брюхоногих моллюсков, фораминифер и особенно аммоноидей (из класса головоногих моллюсков); в последнем случае рекапитуляция связана с тем, что молодь была близка к взрослым особям по образу жизни. Явления, соответствующие Б. з., имеют значение для систематики организмов, напр. в систематике животных для установления таксономич. положения нек-рых групп низших ракообразных (ракушковых, усоногих); эти явления отмечались в многочисленных группах животных (напр., рудиментарные коренные зубы у юных утконосов, впоследствии совершенно исчезающие; многочисленные зубы у зародышей беззубых китов, затем полностью атрофирующиеся). Рекапитуляция и тесно связанные с ней явления известны не только у животных, но и у растений, на что неоднократно указывал, напр., И. В. Мичурин. Так, рассматривая "влияние атавизма", он отмечал, что оно "является неустранимым во всех без исключения сеянцах различных видов и разновидностей растений только в начальных стадиях развития их из семени, выражаясь тем, что все они в молодости имеют в своем наружном виде кажущееся сходство с дикими формами родоначального вида" (Соч., т. 1, 1948, 185–86). Нек-рые зоологи и палеонтологи совершенно отвергают Б. з., ссылаясь на то, что онтогенез никогда не представляет собой даже краткого повторения важнейших этапов филогенеза. Это возражение имеет силу по отношению к первоначальному геккелевскому определению "основного Б. з.", но не может быть выдвинуто против дарвиновского понимания теории рекапитуляции, согласно к-рому онтогенез отнюдь не представляет краткого повторения филогенеза, но на разных стадиях содержит элементы такого повторения. Столь же неосновательно то возражение против основной идеи Б. з., к-рое основывается на случаях, когда эмбриональные структуры, бесспорно соответствующие некогда существовавшим органам предков данной формы, несут какие-то определенные функции у зародышей, т. е. не являются полностью бесполезными: напр., появление в онтогенезе рыб жаберных щелей, обслуживающих у зародышей дыхательную систему, но в дальнейшем исчезающих, надо рассматривать как рекапитуляцию, так как эти щели соответствуют жаберным щелям предков современных рыб; зубы у зародышей беззубых китов имеют определенное значение в индивидуальном развитии этих животных, но это не опровергает того положения, что такие эмбриональные зубы соответствуют зубам предков, у к-рых они служили для захвата пищи. Нек-рые противники теории рекапитуляции утверждают, что в животном мире господствующее положение занимают процессы, как бы противоположные Б. з. Эти ученые утверждают, что эмбриональные структуры предков часто становятся структурами взрослых особей у потомков – у более поздних представителей данного филогенетич. ряда (нем. палеонтолог О. Шиндевольф, англ. зоолог-эмбриолог Г. де Бир). Крайние представители этого направления приходят к мысли, что эволюция шла в основном от высшего состояния к низшему. Такое понимание соотношения между онтогенезом и филогенезом направлено к дискредитации научной, материалистической эволюционной теории в целом. Лит.: Энгельс Ф., Анти-Дюринг, М., 1957, с. 70; Дарвин Ч., Происхождение видов путем естественного отбора, Соч., т. 3, М.–Л.,1939,с. 627–36; Мичурин И. В., Принципы и методы работы, Соч., Т.1, 2 изд., М., 1948, с. 651; Мюллер Ф., Геккель Э., Основной биогенетический закон, Избр. работы, М.–Л. 1940. Л. Давиташвили. Тбилиси. . смотреть
БИОГЕНЕТИЧЕСКИЙ ЗАКОН
БИОГЕНЕТИЧЕСКИЙ ЗАКОН обобщение в области взаимоотношений онтогенеза и филогенеза организмов, установленное Ф. Мюллером (1864) и сформулированное Э. Г. смотреть


Сыграл важную роль в истории развития науки, однако в настоящее время в своем исходном виде не признается современной биологической наукой. [2] [3] По современной трактовке биогенетического закона, предложенной русским биологом А. Н. Северцовым в начале 20 века, в онтогенезе происходит повторение признаков не взрослых особей предков, а их зародышей. [4]
Содержание
История создания
Немецкий анатом и эмбриолог Мартин Ратке (1793—1860) в 1825 г. описал жаберные щели и дуги у эмбрионов млекопитающих и птиц — один из наиболее ярких примеров рекапитуляции [источник не указан 469 дней] .
Примеры выполнения биогенетического закона
Яркий пример выполнения биогенетического закона — развитие лягушки, включающее в себя стадию головастика, который по своему строению гораздо больше похож на рыб, чем на земноводных:
У головастика, как и у низших рыб и рыбьих мальков, основой скелета служит хорда, только впоследствии в туловищной части обрастающая хрящевыми позвонками. Череп у головастика хрящевой, и к нему примыкают хорошо развитые хрящевые дуги; дыхание жаберное. Кровеносная система также построена по рыбьему типу: предсердие ещё не разделилось на правую и левую половины, кровь в сердце поступает только венозная, а оттуда через артериальный ствол идёт к жабрам. Если бы развитие головастика остановилось на этой стадии и не шло дальше, мы должны были бы без всяких колебаний отнести такое животное к надклассу рыб. [7]
Зародыши не только земноводных, но и всех без исключения позвоночных животных также имеют на ранних стадиях развития жаберные щели, двухкамерное сердце и другие признаки, характерные для рыб. Например, птичий зародыш в первые дни насиживания также представляет собой хвостатое рыбообразное существо с жаберными щелями. На этой стадии будущий птенец обнаруживает сходство и с низшими рыбами, и с личинками амфибий, и с ранними стадиями развития других позвоночных животных (в том числе и человека [7] [8] ). На последующих стадиях развития зародыш птицы становится похожим на пресмыкающихся:
И пока у куриного зародыша до конца первой недели и задние, и передние конечности имеют вид одинаковых лапок, пока хвост ещё не успел исчезнуть, а из сосочков ещё не сформировались перья, он по всем своим признакам стоит ближе к пресмыкающимся, чем к взрослым птицам. [7]
Зародыш человека в ходе эмбриогенеза проходит через аналогичные стадии. Затем, за период примерно между четвертой и шестой неделями развития он превращается из рыбоподобного организма в организм, неотличимый от зародыша обезьяны, и только потом приобретает человеческие черты. [9] [10]
Такое повторение признаков предков в ходе индивидуального развития особи Геккель назвал рекапитуляция.
- a1 — a2
- a1' — a2' — a3'
- a1" — a2" — a3" — a4"
(На этой схеме сверху вниз расположены виды-предки и виды-потомки, а слева направо — стадии их онтогенеза.)
Факты, противоречащие биогенетическому закону
Уже в XIX веке было известно достаточно фактов, противоречащих биогенетическому закону. Так, были известны многочисленные примеры неотении, при которой в ходе эволюции происходит укорочение онтогенеза и выпадение его конечных стадий. В случае неотении взрослая стадия вида-потомка напоминает личиночную стадию вида-предка, а не наоборот, как этого следовало бы ожидать при полной рекапитуляции.
Связь биогенетического закона с дарвинизмом
Биогенетический закон часто рассматривается как подтверждение дарвиновской теории эволюции, хотя он вовсе не следует из классического эволюционного учения.
Например, если вид А3 возник путём эволюции из более древнего вида А1 через ряд переходных форм (A1 =>А2 => A3), то, в соответствии с биогенетическим законом (в его модифицированном варианте), возможен и обратный процесс, при котором вид А3 превращается в А2 путем укорочения развития и выпадения его конечных стадий (неотения или педогенез).
Дарвинизм и синтетическая теория эволюции, напротив, отрицают возможность полного возврата к предковым формам (Закон необратимости эволюции Долло). Причиной этого, в частности, являются перестройки эмбрионального развития на его ранних стадиях (архаллаксисы по А. Н. Северцову), при которых генетические программы развития меняются настолько существенно, что их полное восстановление в ходе дальнейшей эволюции становится практически невероятным.
Научная критика биогенетического закона и дальнейшее развитие учения о связи онтогенеза и филогенеза
Накопление фактов и теоретические разработки показали, что биогенетический закон в формулировке Геккеля в чистом виде никогда не выполняется. Рекапитуляция может быть только частичной.
Филэмбриогенезы — это такие изменения онтогенеза, которые в ходе эволюции приводят к изменению признаков взрослых особей. Северцов разделил филэмбриогенезы на анаболии, девиации и архаллаксисы. Анаболия — удлинение онтогенеза, сопровождающееся надставкой стадий. Только при этом способе эволюции наблюдается рекапитуляция — признаки зародышей или личинок потомков напоминают признаки взрослых предков. При девиации происходят изменения на средних стадиях развития, что приводят к более резким изменениям в строении взрослого организма, чем при анаболии. При этом способе эволюции онтогенеза рекапитулировать признаки предковых форм могут лишь ранние стадии потомков. При архаллаксисах изменения происходят на самых ранних стадиях онтогенеза, изменения в строении взрослого организма наиболее часто существенны, а рекапитуляции невозможны.

Наблюдение двух независимых биологов за онтогенезом организмов позволило сформировать биогенетический закон Геккеля-Мюллера. Впервые формулировка прозвучала в 1866 году. Однако предпосылки становления закона были выявлены ещё в 1820-х годах.
Закон и его значение
Суть закона заключается в том, что в процессе онтогенеза (индивидуального развития организма) особь повторяет формы своих предков и от зачатия до становления проходит стадии филогенеза (исторического развития организмов).
Через два года естествоиспытатель Эрнст Геккель сформулировал закон более кратко: онтогенез – быстрое повторение филогенеза. Другими словами, каждый организм проходит эволюционное изменение вида в процессе развития.
Свои выводы учёные сделали при изучении эмбрионов разных видов на основе ряда схожих признаков. Например, у зародышей млекопитающих и рыб формируются жаберные дуги. Эмбрионы амфибий, рептилий и млекопитающих проходят одинаковые стадии развития и внешне похожи. Схожесть эмбрионов является одним из доказательств теории эволюции и происхождения животных от одного предка.
Сайт учителей биологии МБОУ Лицей № 2 г. Воронежа, РФ
Site biology teachers lyceum № 2 Voronezh city, Russian Federation
Закон зародышевого сходства — полученное опытным путём (эмпирическое) обобщение Карла Максимовича Бэра в 1828 году, которое звучит следующим образом: в онтогенезе всех животных сначала выявляются признаки высших категорий (типа, класса), в ходе дальнейшей эмбриональной дифференцировки развиваются особенности отряда, семейства, рода, вида и особи.
Представители разных групп организмов (например, классов подтипа позвоночных) на ранних стадиях эмбриогенеза обычно сходны друг с другом. Так, в онтогенезе курицы прежде всего обозначаются характерные черты типа хордовых, позднее — подтипа позвоночных, затем класса птиц, отряда курообразных и, наконец, индивидуальные качества.

Расхождение признаков зародышей в процессе развития называется эмбриональной дивергенцией. Это явление объясняется историей развития данного вида и отражает эволюцию той или иной систематической группы животных.
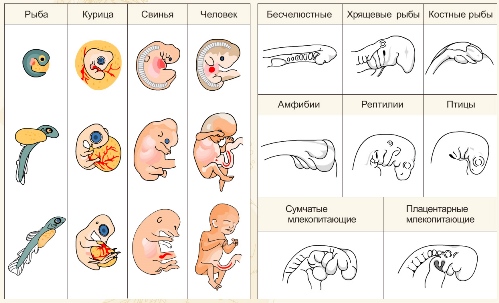
Биогенетический закон сформулирован немецким учёным Э. Геккелем в 1866 году, состоит в том, что индивидуальное развитие особи (онтогенез) является кратким и быстрым повторением важнейших этапов эволюции вида — филогенеза.
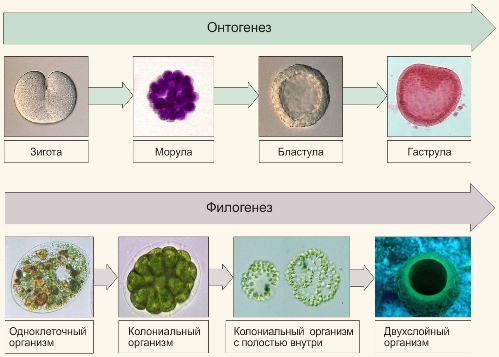
Факты, свидетельствующие о закладке у зародышей наземных позвоночных жаберных щелей, были известны ещё до появления эволюционного учения Дарвина. Но именно он дал этому объяснение, установив, что стадии развития зародышей воспроизводят древние предковые формы. Дарвин рассматривал этот факт как фундаментальную закономерность эволюции органического мира.
Немецкий эмбриолог Ф. Мюллер в 1864 году подтвердил это данными из истории развития ракообразных.
Геккель сформулировал биогенетический закон в 1866 году, используя представления Дарвина и закон зародышевого сходства, сформулированный Бэром.
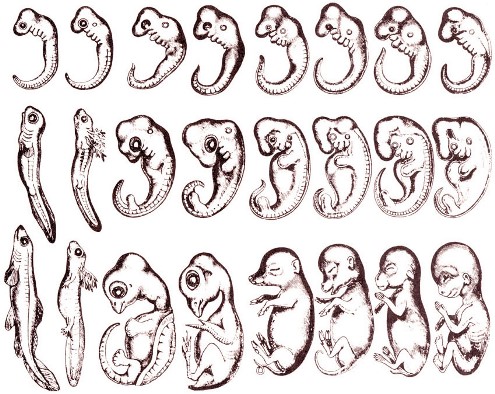
Биогенетический закон сыграл важную роль в биологии, обобщил эволюционные исследования в эмбриологии, сравнительной анатомии и палеонтологии.
Читайте также: