
Почему число мышечных волокон в двигательных единицах разных мышц различается кратко
Обновлено: 06.07.2024
Сила — векторная величина, являющаяся мерой механического воздействия на материальную точку или тело со стороны других тел или полей (Б. М. Яворский, А. А. Детлаф, 1974). Сила полностью задана, если указаны ее численное значение, направление и точка приложения.
В теории и методике физического воспитания рассматривают физическое качество силу как способность человека напряжением мышц преодолевать механические и биомеханические силы, препятствующие действию (Зациорский В. М., 1972; Л. П. Матвеев, 1991).
Мышцы могут проявлять силу: без изменения своей длины (изометрический режим), при уменьшении длины (изотонический режим), при удлинении (эксцентрический режим), при использовании специальной аппаратуры возможно соблюдение изокинетического режима (в ходе сокращения мышц соблюдается либо постоянная скорость, либо сила).
Силовое проявление мышцы зависит от:
— интенсивности активации мотнейронного пула спинного мозга данной мышцы;
— количества активированных двигательных единиц и мышечных волокон;
— количества миофибрилл в каждом мышечном волокне;
— скорости сокращения миофибрилл, которая зависти от активности миозиновой АТФ-азы и величины внешнего сопротивления;
— законов механики мышечного сокращения (сила — длина мышцы, сила — скорость сокращения);
— начального состояния исполнительного аппарата (утомленные мышечные волокна демонстрируют меньшую силу).
Наличие последовательной упругой компоненты в мышечных волокнах приводит к тому, что с ростом числа рекрутированных МВ увеличивается жесткость мышцы -коэффициент упругости (В. М. Зациорский с соав., 1981).
Растягивание активной мышцы приводит не только к накоплению энергии упругой деформации в последовательной упругой компоненте, но и к прекращению работы мостиков, а именно, они перестают отцепляться за счет энергии молекул АТФ. Разрыв мостиков происходит благодаря действию внешней — механической силы. В итоге отрицательная работа мышц выполняется с очень высоким коэффициентов полезного действия, с минимальными затратами АТФ, а значит и кислорода.
Различия связаны со степенью нагрузки на ту или иную мышцу. Нагрузка может быть как динамической, так и статической. Т.е. те мышцы, которые активны больше других, или на них оказывается нагрузка сильнее и продолжительнее, чем на другие, будут содержать большее количество волокон. Работа происходит в момент сокращения, которое приводит кости в движение. Во время сокращения мышца укорачивается и утолщается, затрачивая энергию и увеличивая потребление кислорода клетками. При длительной или избыточной нагрузке в мышцах наступает утомление, связанное с нехваткой кислорода и избытком продуктов обмена веществ.
Дано понятие двигательной единицы (ДЕ) и описана ее структура. Приведена классификация ДЕ и соответствие ДЕ и типов мышечных волокон. Описан принцип размера и правило Хеннемана. Приведены данные об активации ДЕ при выполнении силовых упражнений в зависимости от величины отягощения.
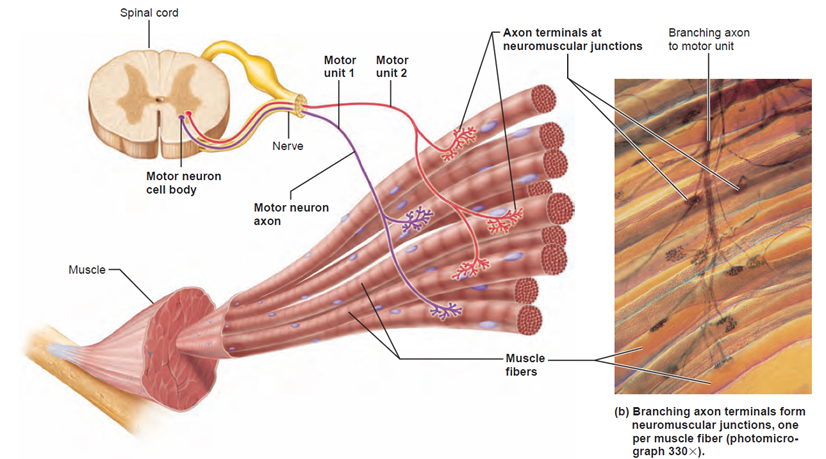
ДВИГАТЕЛЬНЫЕ ЕДИНИЦЫ (ДЕ)
Определение
В настоящее время под двигательной единицей (ДЕ) понимается элементарная функциональная единица мышцы, включающая в себя мотонейрон и иннервируемые им мышечные волокна.
Структура ДЕ
Войдя в мышцу, аксон мотонейрона разветвляется на множество веточек, каждая из которых иннервирует отдельное мышечное волокно. Таким образом, один мотонейрон иннервирует достаточно большое количество мышечных волокон (от нескольких единиц до нескольких тысяч), в то время как каждое мышечное волокно иннервируется только одним двигательным нейроном.
Размер ДЕ (иннервационное отношение, коэффициент иннервации)
Размер ДЕ — это количество мышечных волокон, которые иннервируются одним мотонейроном. Чтобы определить этот показатель определяют количество мышечных волокон в скелетной мышце и количество мотонейронов, которые инннервируют эти мышечные волокна (табл.1). Иногда в литературе размер ДЕ называют иннервационным отношением или коэффициентом иннервации.
Всякий раз, когда активируется мотонейрон, он посылает потенциалы действия ко всем мышечным волокнам, которые он иннервирует. Поэтому, чем ниже коэффициент иннервации, тем совершеннее контроль со стороны нервной системы за мышечными волокнами. По коэффициенту иннервации (размеру ДЕ) можно судить о количестве веточек, необходимых аксону мотонейрона, чтобы иннервировать все входящие в ДЕ мышечные волокна.
Табл. 1 — Количество мышечных волокон, количество ДЕ (мотонейронов) и размер ДЕ в различных скелетных мышцах человека
| Мышца | Количество мышечных волокон | Количество ДЕ | Размер ДЕ |
| Передняя большеберцовая | 250090 | 445 | 562 |
| Медиальная головка икроножной мышцы | 1120365 | 1934 | 579 |
| Наружная прямая мышца глаза | 26730 | 2970 | 9 |
| Плечелучевая | 136530 | 333 | 410 |
С возрастом количество ДЕ, приходящихся на одну мышцу уменьшается. Это связано с тем, что уменьшается количество мотонейронов, которые иннервируют отдельную мышцу. Вследствие этого количество мышечных волокон по мере старения организма человека также уменьшается.
Классификации ДЕ
Существуют различные классификации ДЕ. Исходя из значимости для организма, Р. Берк с соавт. (R.E. Burke, 1973) предложил разделять ДЕ по сочетанию двух признаков – скорости сокращения и устойчивости к утомлению. По этой классификации ДЕ делятся на три типа: S (slow) – медленные, устойчивые к утомлению; FR (fast resistant) – быстрые, устойчивые к утомлению, FF (fast fatigable) – быстрые, быстроутомляемые. Этим ДЕ соответствуют различные типы мышечных волокон (табл. 1).
Таблица 1 — Соответствие типов ДЕ и мышечных волокон
| Тип ДЕ | S | FR | FF |
| Тип мышечного волокна | I тип | IIA тип | IIB тип |
Строение и функции мотонейрона соответствуют морфологическим характеристикам мышечных волокон, которые он иннервирует. Так, мотонейрон ДЕ S типа имеет небольшое клеточное тело и иннервирует от 10 до 180 мышечных волокон, а мотонейрон ДЕ FF типа имеет большое клеточное тело и иннервирует от 300 до 800 мышечных волокон (Дж.Х. Уилмор, Д.Л. Костилл, 1997) (рис.1).
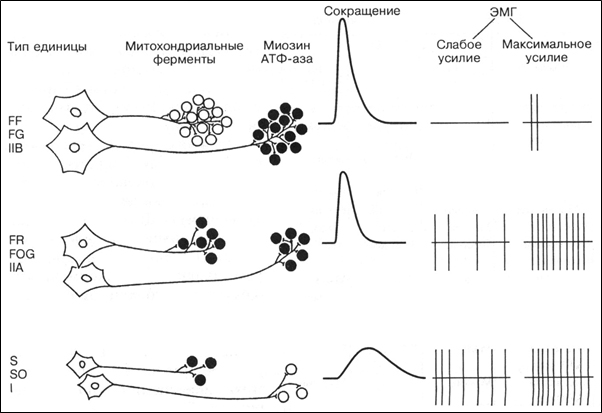
Рис. 1. Гистохимические и физиологические свойства трех основных типов ДЕ и мышечных волокон (R.E. Burke, 1973)
В табл. 2 представлено количество мышечных волокон и количество ДЕ в различных мышцах человека
Принцип размера или правило Хеннемана
ДЕ S типа имеют низкий порог активации, поэтому при развитии силы мышцы они включаются в работу первыми. После этого активируются ДЕ FR типа. ДЕ FF типа обладают высоким порогом активации, поэтому при развитии усилия в мышце они активируются последними.
Благодаря тому, что мышечные волокна, принадлежащие различным ДЕ, рассредоточены по всей мышце, а не находятся в одном пучке, развитие силы мышцы характеризуется плавностью. Однако из-за того, что между соседними мышечными волокнами существуют соединительнотканные связи, при сокращении одних мышечных волокон, например, входящих в состав ДЕ S типа, и расслабленном состоянии других (например, входящих в состав ДЕ FF типа) должны возникать силы трения, обусловливающие высокую вязкость мышцы. Г.В. Васюков (1967) показал, что при небольших напряжениях мышцы (30% от максимума) ее вязкость максимальна. При дальнейшем напряжении мышцы, когда одновременно возбуждено много мышечных волокон, вязкость мышцы скачкообразно уменьшается.
Активация ДЕ в зависимости от различной величины внешней нагрузки
В настоящее время установлено, что в зависимости от величины внешнего отягощения активируются разные ДЕ. Эти данные представлены в табл. 3
Таблица 33 – Активация ДЕ в зависимости от различной степени отягощения.
| Степень отягощения, % | Особенности активации ДЕ |
| 20-30% от максимального | Рекрутируются ДЕ S типа. |
| От 30 до 50% от максимального | Рекрутируются ДЕ S и FR типа |
| От 50 до 70 % от максимального | Рекрутируются ДЕ: S, FR FF типов. |
| Более 70% от максимума | Синхронизация активности ДЕ, то есть одновременное возбуждение большинства мышечных волокон. |
Похожие записи:
Тест времени реакции на сигнал
Представлена программа расчета времени реакции на сигнал, предназначенная для использования в учебных целях, например на занятиях по…
Саркоплазматическая гипертрофия мышц
Дано определение и описаны механизмы саркоплазматической гипертрофии скелетных мышц. Показано, что этот вид гипертрофии мышц широко…
Классификация типов конституции человека М.В. Черноруцкого
Рассмотрена классификация типов конституции человека, разработанная выдающимся терапевтом М.В. Черноруцким в 1925 году. Классификация типов конституции человека М.В.
Типы гипертрофии скелетных мышц человека
В статье дается классификация различных видов гипертрофии скелетных мышц человека на основе ряда классификационных признаков: времени проявления…
Миомейкер: Мембранный активатор слияния миобластов и образования мышц
Ученые установили, что для образования мышечных волокон необходимо слияние клеток-предшественников, которые называются миобластами. Эти клетки имеют только…
Основным морфо-функциональным элементом нервно-мышечного аппарата скелетных мышц является двигательная единица (ДЕ). Она включает мотонейрон спинного мозга с иннервируемыми его аксоном мышечными волокнами. Внутри мышцы этот аксон образует несколько концевых веточек. Каждая такая веточка образует контакт – нервно-мышечный синапс на отдельном мышечном волокне. Нервные импульсы, идущие от мотонейрона, вызывают сокращения определенной группы мышечных волокон. Двигательные единицы мелких мышц, осуществляющих тонкие движения (мышцы глаза, кисти), содержат небольшое количество мышечных волокон. В крупных их в сотни раз больше. Все ДЕ в зависимости от функциональных особенностей делятся на 3 группы:
I. Медленные неутомляемые. Они образованы "красными" мышечными волокнами, в которых меньше миофибрилл. Скорость сокращения и сила этих волокон относительно небольшие, но они мало утомляемы. Поэтому их относят к тоническим. Регуляция сокращений таких волокон осуществляется небольшим количеством мотонейронов, аксоны которых имеют мало концевых веточек. Пример – камбаловидная мышца.
IIВ. Быстрые, легко утомляемые. Мышечные волокна содержат много миофибрилл и называются "белыми". Быстро сокращаются и развивают большую силу, но быстро утомляются. Поэтому их называют фазными. Мотонейроны этих ДЕ самые крупные, имеют толстый аксон с многочисленными концевыми веточками. Они генерируют нервные импульсы большой частоты. Мышцы глаза.
IIA. Быстрые, устойчивые к утомлению. Занимают промежуточное положение.
30)(Виды и режимы сокращений скелетных мышц. Одиночное мышечное сокращение. Тетанус, его виды, механизм)
У скелетной мышцы выделяют одиночное сокращение и суммированное сокращение, или тетанус. Одиночное сокращение это сокращение, которое возникает на одиночный стимул, достаточный для вызова возбуждения мышцы. После короткого скрытого периода (латентный период) начинается процесс сокращения. При регистрации сократительной активности в изометрических условиях (два конца неподвижно закреплены) в первую фазу происходит нарастание напряжения (силы), а во вторую ее падение до исходной величины. Соответственно эти фазы называют фазой напряжения и фазой расслабления. При регистрации сократительной активности в изотоническом режиме (например, в условиях обычной миографической записи) эти фазы будут называться соответственно фазой укорочения и фазой удлинения. В среднем сократительный цикл длится около 200 мс (мышцы лягушки) или 30-80 мс (у теплокровных). Если на мышцу действует серия прямых раздражении (минуя нерв) или непрямых раздражении (через нерв), но с большим интервалом, при котором всякое следующее раздражение попадает в период после окончания 2-й фазы, то мышца будет на каждый из этих раздражителей отвечать одиночным сокращением. Суммированные сокращения возникают в том случае, если на мышцу наносятся 2 и более раздражения, причем всякое последующее раздражение (после предыдущего) наносится либо во время 2-й фазы (расслабления или удлинения), либо во время 1-й фазы (укорочения или напряжения).
(Режимы сокращения мышц)
Для скелетной мышцы характерны два основных режима сокращения изометрический и изотонический. Изометрический режим проявляется в том, что в мышце во время ее активности нарастает напряжение (генерируется сила), но из-за того, что оба конца мышцы фиксированы (например, мышца пытается поднять большой груз) она не укорачивается. Изотонический режим проявляется в том, что мышца первоначально развивает напряжение (силу), способную поднять данный груз, а потом мышца укорачивается меняет свою длину, сохраняя напряжение, равное весу поднимаемого груза. Так как изотоническое сокращение не является "чисто" изотоническим (элементы изометрического сокращения имеют место в самом начале сокращения мышцы), а изометрическое сокращение тоже не является "чисто" изотоническим (элементы смещения все-таки есть, несомненно), то предложено употреблять термин "ауксотоническое сокращение" смешанное по характеру. Понятия "изотонический", "изометрический" важны для анализа сократительной активности изолированных мышц и для понимания биомеханики сердца. Режимы сокращения гладких мышц. Целесообразно выделить изометрический и изотонический режимы (и, как промежуточный ауксотонический). Например,когда мышечная стенка полого органа начинает сокращаться, а орган содержит жидкость, выход для которой перекрыт сфинктером, то возникает ситуация изометрического режима: давление внутри полого органа растет, а размеры ГМК не меняются (жидкость не сжимается). Если это давление станет высоким и приведет к открытию сфинктера, то ГМК переходит в изотонический режим функционирования происходит изгнание жидкости, т.е. размеры ГМК уменьшаются, а напряжение или сила сохраняется постоянной и достаточной для изгнания жидкости.

(Одиночное сокращение)
Механический ответ отдельного мышечного волокна на одиночный потенциал действия называется одиночным сокращением. Основные характеристики одиночного изометрического сокращения показаны на , а. Начало мышечного напряжения запаздывает на несколько миллисекунд по отношению к потенциалу действия. В течение этого латентного периода проходят все этапы электромеханического сопряжения .
Интервал от начала развития напряжения до момента его максимума - это время сокращения . Оно различно для разных типов волокон скелетных мышц. Время сокращения быстрых волокон не превышает 10 мс, тогда как для более медленных волокон оно не меньше 100 мс. Длительность сокращения определяется тем, как долго цитоплазматическая концентрация Са2+ остается повышенной, обеспечивая продолжение циклической активности поперечных мостиков . Время сокращения обусловлено активностью Са2+-АТФ фазы саркоплазматического ретикулума , которая в быстрых волокнах выше, чем в медленных.
Сравнение одиночных сокращений одного и того же мышечного волокна при разных режимах его деятельности показывает, что латентный период больше для изотонического сокращения , чем для изометрического , тогда как длительность механического процесса меньше в случае изотонического сокращения (т.е. при укорочении), чем изометрического (т.е. при генерировании силы).
Характеристики изотонического сокращения зависят также от веса поднимаемой нагрузки). А именно, при более тяжелой нагрузке:
- латентный период продолжительнее;
- скорость укорочения (величина укорочения мышцы в единицу времени), длительность сокращения и величина укорочения мышцы меньше.
Рассмотрим подробнее последовательность явлений во время изотонического одиночного сокращения. При возбуждении мышечного волокна поперечные мостики начинают развивать силу, однако укорочение не начнется, пока мышечное напряжение не превысит нагрузку на волокно. Таким образом, укорочению предшествует период изометрического сокращения , в течение которого возрастает напряжение. Чем тяжелее нагрузка, тем больше потребуется времени, чтобы оно сравнялось с величиной нагрузки и началось укорочение. Если нагрузку повышать, то в конце концов мышечное волокно не сможет ее поднять, скорость и степень укорочения будут равны нулю и сокращение станет чисто изометрическим.
Основным морфо-функциональным элементом нервно-мышечного аппарата скелетных мышц является двигательная единица (ДЕ). Она включает мотонейрон спинного мозга с иннервируемыми его аксоном мышечными волокнами. Внутри мышцы этот аксон образует несколько концевых веточек. Каждая такая веточка образует контакт – нервно-мышечный синапс на отдельном мышечном волокне. Нервные импульсы, идущие от мотонейрона, вызывают сокращения определенной группы мышечных волокон. Двигательные единицы мелких мышц, осуществляющих тонкие движения (мышцы глаза, кисти), содержат небольшое количество мышечных волокон. В крупных их в сотни раз больше. Все ДЕ в зависимости от функциональных особенностей делятся на 3 группы:
I. Медленные неутомляемые. Они образованы "красными" мышечными волокнами, в которых меньше миофибрилл. Скорость сокращения и сила этих волокон относительно небольшие, но они мало утомляемы. Поэтому их относят к тоническим. Регуляция сокращений таких волокон осуществляется небольшим количеством мотонейронов, аксоны которых имеют мало концевых веточек. Пример – камбаловидная мышца.
IIВ. Быстрые, легко утомляемые. Мышечные волокна содержат много миофибрилл и называются "белыми". Быстро сокращаются и развивают большую силу, но быстро утомляются. Поэтому их называют фазными. Мотонейроны этих ДЕ самые крупные, имеют толстый аксон с многочисленными концевыми веточками. Они генерируют нервные импульсы большой частоты. Мышцы глаза.
IIA. Быстрые, устойчивые к утомлению. Занимают промежуточное положение.
30)(Виды и режимы сокращений скелетных мышц. Одиночное мышечное сокращение. Тетанус, его виды, механизм)
У скелетной мышцы выделяют одиночное сокращение и суммированное сокращение, или тетанус. Одиночное сокращение это сокращение, которое возникает на одиночный стимул, достаточный для вызова возбуждения мышцы. После короткого скрытого периода (латентный период) начинается процесс сокращения. При регистрации сократительной активности в изометрических условиях (два конца неподвижно закреплены) в первую фазу происходит нарастание напряжения (силы), а во вторую ее падение до исходной величины. Соответственно эти фазы называют фазой напряжения и фазой расслабления. При регистрации сократительной активности в изотоническом режиме (например, в условиях обычной миографической записи) эти фазы будут называться соответственно фазой укорочения и фазой удлинения. В среднем сократительный цикл длится около 200 мс (мышцы лягушки) или 30-80 мс (у теплокровных). Если на мышцу действует серия прямых раздражении (минуя нерв) или непрямых раздражении (через нерв), но с большим интервалом, при котором всякое следующее раздражение попадает в период после окончания 2-й фазы, то мышца будет на каждый из этих раздражителей отвечать одиночным сокращением. Суммированные сокращения возникают в том случае, если на мышцу наносятся 2 и более раздражения, причем всякое последующее раздражение (после предыдущего) наносится либо во время 2-й фазы (расслабления или удлинения), либо во время 1-й фазы (укорочения или напряжения).
(Режимы сокращения мышц)
Для скелетной мышцы характерны два основных режима сокращения изометрический и изотонический. Изометрический режим проявляется в том, что в мышце во время ее активности нарастает напряжение (генерируется сила), но из-за того, что оба конца мышцы фиксированы (например, мышца пытается поднять большой груз) она не укорачивается. Изотонический режим проявляется в том, что мышца первоначально развивает напряжение (силу), способную поднять данный груз, а потом мышца укорачивается меняет свою длину, сохраняя напряжение, равное весу поднимаемого груза. Так как изотоническое сокращение не является "чисто" изотоническим (элементы изометрического сокращения имеют место в самом начале сокращения мышцы), а изометрическое сокращение тоже не является "чисто" изотоническим (элементы смещения все-таки есть, несомненно), то предложено употреблять термин "ауксотоническое сокращение" смешанное по характеру. Понятия "изотонический", "изометрический" важны для анализа сократительной активности изолированных мышц и для понимания биомеханики сердца. Режимы сокращения гладких мышц. Целесообразно выделить изометрический и изотонический режимы (и, как промежуточный ауксотонический). Например,когда мышечная стенка полого органа начинает сокращаться, а орган содержит жидкость, выход для которой перекрыт сфинктером, то возникает ситуация изометрического режима: давление внутри полого органа растет, а размеры ГМК не меняются (жидкость не сжимается). Если это давление станет высоким и приведет к открытию сфинктера, то ГМК переходит в изотонический режим функционирования происходит изгнание жидкости, т.е. размеры ГМК уменьшаются, а напряжение или сила сохраняется постоянной и достаточной для изгнания жидкости.
(Одиночное сокращение)
Механический ответ отдельного мышечного волокна на одиночный потенциал действия называется одиночным сокращением. Основные характеристики одиночного изометрического сокращения показаны на , а. Начало мышечного напряжения запаздывает на несколько миллисекунд по отношению к потенциалу действия. В течение этого латентного периода проходят все этапы электромеханического сопряжения .
Интервал от начала развития напряжения до момента его максимума - это время сокращения . Оно различно для разных типов волокон скелетных мышц. Время сокращения быстрых волокон не превышает 10 мс, тогда как для более медленных волокон оно не меньше 100 мс. Длительность сокращения определяется тем, как долго цитоплазматическая концентрация Са2+ остается повышенной, обеспечивая продолжение циклической активности поперечных мостиков . Время сокращения обусловлено активностью Са2+-АТФ фазы саркоплазматического ретикулума , которая в быстрых волокнах выше, чем в медленных.
Сравнение одиночных сокращений одного и того же мышечного волокна при разных режимах его деятельности показывает, что латентный период больше для изотонического сокращения , чем для изометрического , тогда как длительность механического процесса меньше в случае изотонического сокращения (т.е. при укорочении), чем изометрического (т.е. при генерировании силы).
Характеристики изотонического сокращения зависят также от веса поднимаемой нагрузки). А именно, при более тяжелой нагрузке:
- латентный период продолжительнее;
- скорость укорочения (величина укорочения мышцы в единицу времени), длительность сокращения и величина укорочения мышцы меньше.
Рассмотрим подробнее последовательность явлений во время изотонического одиночного сокращения. При возбуждении мышечного волокна поперечные мостики начинают развивать силу, однако укорочение не начнется, пока мышечное напряжение не превысит нагрузку на волокно. Таким образом, укорочению предшествует период изометрического сокращения , в течение которого возрастает напряжение. Чем тяжелее нагрузка, тем больше потребуется времени, чтобы оно сравнялось с величиной нагрузки и началось укорочение. Если нагрузку повышать, то в конце концов мышечное волокно не сможет ее поднять, скорость и степень укорочения будут равны нулю и сокращение станет чисто изометрическим.
Строение мышечной ткани. Мышечное волокно в физиологии
Около 40% массы тела человека составляют скелегные мышцы, и, вероятно, 10% приходится на долю гладких мышц и мышцы сердца. Некоторые из основных принципов сокращения являются общими для разных типов мышц.
На рисунке показана схема организации скелетной мышцы. Все скелетные мышцы содержат множество волокон диаметром от 10 до 80 мкм. На рисунке также показано, что каждое из этих волокон состоит из последовательно соединенных более мелких элементов.
В большинстве скелетных мышц каждое волокно вытянуто во всю длину мышцы и, за исключением примерно 2%, обычно иннервируется лишь одним нервным окончанием вблизи середины волокна.
Сарколемма. Сарколеммой называют клеточную мембрану мышечного волокна. Она состоит из собственно мембраны клетки, называемой плазматической мембраной, и наружного покрытия из тонкого слоя полисахаридного материала, который содержит множество тонких коллагеновых нитей. На концах мышечного волокна этот поверхностный слой сарколеммы сливается с сухожильными волокнами. Сухожильные волокна, в свою очередь, собираются в пучки и формируют мышечные сухожилия, вплетающиеся в кости.
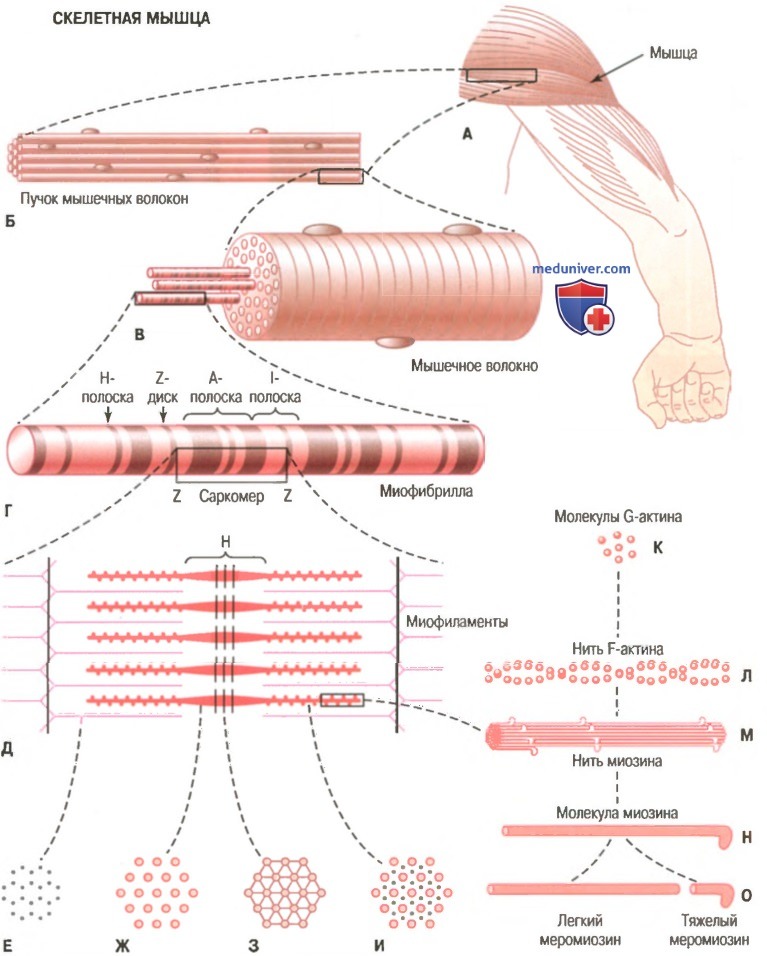
Организация скелетной мышцы от макроскопического до молекулярного уровня. Е, Ж 3, И - поперечное сечение на указанном уровне.
Миофибриллы. Нити актина и миозина. Каждое мышечное волокно содержит от нескольких сотен до нескольких тысяч миофибрилл, которые представлены на поперечном срезе в виде небольших светлых точек. Каждая миофибрилла собрана примерно из 1500 смежных нитей миозина и 3000 нитей актина, представляющих собой большие полимеризованные белковые молекулы, ответственные за мышечное сокращение. Они видны в продольном сечении на электронной микрофотографии. Толстые нити на схемах — миозиновые, тонкие — актиновые.
Обратите внимание, что на рисунке актиновые и миозиновые нити частично перекрываются, в связи с чем миофибриллы имеют чередующиеся светлые и темные полоски. Светлые полоски содержат только актиновые нити, их называют I-полосками, поскольку они изотропны в поляризованном свете. Темные полоски содержат миозиновые нити, а также концы актиновых нитей, которые частично накладываются на миозиновые нити. Эти полоски называют А-полосками, поскольку они анизотропны в поляризованном свете. На рисунке видны также маленькие выступы по сторонам миозиновых нитей. Это так называемые поперечные мостики. Их взаимодействие с актиновыми нитями лежит в основе сокращения мыщц.
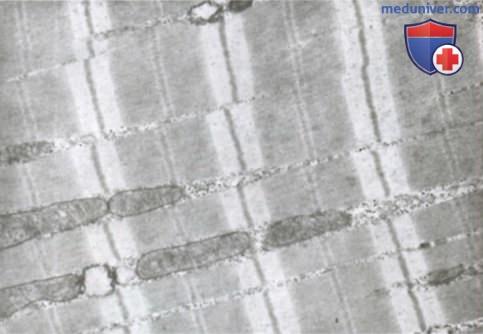
Электронная микрофотография миофибриллы, демонстрирующая детальную организацию актиновых и миозиновых нитей. Видны митохондрии, лежащие между миофибриллами.
На рисунке показано также, что концы актиновых нитей прикреплены к так называемому Z-диску (Z-мембране). Отсюда актиновые нити распространяются в обе стороны, проходя между миозиновыми нитями. Z-диск, состоящий из нитевидных белков, отличающихся от актиновых и миозиновых нитей, проходит в поперечном направлении через миофибриллу, а также через все мышечное волокно, переходя от одной миофибриллы к другой, скрепляя их между собой. В связи с этим целое мышечное волокно, как и отдельные миофибриллы, имеет светлые и темные полоски. Эти полоски придают скелетным и сердечной мышцам характерный полосатый вид.
Часть миофибриллы (или целого мышечного волокна), расположенную между двумя последовательными Z-дисками, называют саркомером. В сокращенном мышечном волокне длина саркомера равна примерно 2 мкм. При этой длине актиновые нити полностью перекрывают миозиновые, и кончики актиновых нитей начинают перекрывать друг друга. Мы увидим далее, что при этой длине мышца способна генерировать максимальную силу сокращения.
Что удерживает актиновые и миозиновые нити на месте? Удерживать актиновые и миозиновые нити рядом довольно трудно. Это обеспечивается с помощью большого числа нитевидных молекул белка, называемого тайтином. Каждая молекула тайтина имеет молекулярную массу около 3000000, что делает ее одной из самых крупных белковых молекул в организме. Кроме того, поскольку молекула тайтина волокнистая, она очень упруга. Упругие молекулы тайтина образуют каркас, удерживающий актиновые и миозиновые нити в положении, обеспечивающем нормальную работу сократительного аппарата саркомера. Это позволяет полагать, что сама молекула тайтина действует как матрица для начального формирования участков сократительных нитей саркомера, особенно миозиновых нитей.
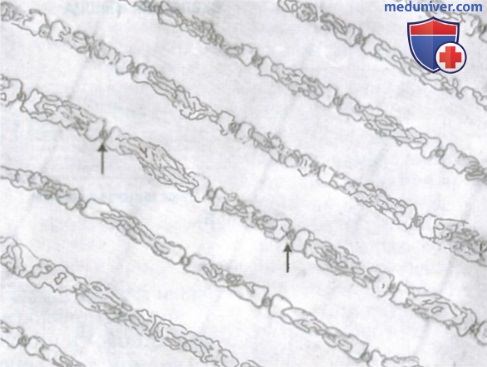
Саркоплазматический ретикулум во внутриклеточных пространствах между миофибриллами с системой продольных трубочек, параллельных миофибриллам. Показаны также в поперечном сечении Т-трубочки (стрелки), которые связаны с внеклеточным пространством и отвечают за проведение электрического сигнала в центр мышечного волокна.
Саркоплазма. В каждом мышечном волокне многочисленные миофибриллы расположены рядом, бок о бок. Пространство между ними заполнено внутриклеточной жидкостью, называемой саркоплазмой и содержащей большое количество калия, магния и фосфатов, а также множество ферментов. Кроме того, имеется огромное число митохондрий, расположенных параллельно миофибриллам. Они снабжают сокращающиеся миофибриллы большим количеством энергии в форме аденозинтрифосфата, синтезируемого митохондриями.
Саркоплазматический ретикулум. В саркоплазме, окружающей миофибриллы каждого мышечного волокна, имеется также хорошо развитый ретикулум, называемый саркоплазматическим ретикулумом. Он имеет специфическую организацию, исключительно важную для регуляции мышечного сокращения . Саркоплазматический ретикулум особенно сильно развит в очень быстро сокращающихся типах мышечных волокон.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Читайте также:
- Какие точки зрения существуют на возможность достижения равенства и справедливости в обществе кратко
- Анализ деятельности воспитателя старшей группы в детском саду
- Анализ дьяволиада булгакова кратко
- Что влияет на качество образования в начальной школе
- Чем по вашему мнению зеленый дом ф дольто отличается от детского сада