
Какую роль играют мышечные рецепторы в процессе движения кратко
Обновлено: 02.07.2024
Лекция 2
Функционирование рецепторного аппарата мышц
2.1. Рецепторы опорно-двигательного аппарата человека
При изучении анатомии и физиологии (А.С. Солодков, Е.Б. Сологуб, 2001) вы изучали двигательную сенсорную систему. Одним из отделов этой системы являются проприорецепторы, расположенные в мышцах, сухожилиях и суставных сумках. В мышцах расположены мышечные веретена, в сухожилиях – сухожильные органы Гольджи. В суставных сумках расположены рецепторы суставов.
Мышечные веретена
Еще в XIX веке В. Кюне обнаружил в скелетных мышцах структуры, напоминающие веретено. Затем, в начале XX века Нобелевский лауреат Чарльз Скотт Шеррингтон показал, что эти структуры служат чувствительными рецепторами. Мышечные веретена рассеяны по всем скелетным мышцам. Концы их обычно прикрепляются к мышечным волокнам параллельно. Каждое веретено покрыто капсулой, которая расширятся в центре и образует ядерную сумку. Внутри веретена содержатся интрафузальные мышечные волокна. Эти волокна в 2-3 раза тоньше обычных (экстрафузальных) волокон скелетных мышц.
Интрафузальные волокна подразделяются на два типа:
- Длинные и толстые (диаметр 20-25 мкм)[1], которые информируют ЦНС о динамическом компоненте движения – скорости изменения длины мышцы. Таких волокон в мышечном веретене не более двух.
- Короткие и тонкие (диаметр 10–12 мкм), которые информируют ЦНС о статическом компоненте движения – текущей длине мышцы. Таких волокон в мышечном веретене от 2 до 12.
Рецепторы Гольджи
Нервно-сухожильные веретена (рецепторы Гольджи) открыл в 1903 году Камилло Гольджи. Впоследствии за эти исследования ему была присуждена Нобелевская премия. Рецепторы Гольджи располагаются в месте перехода мышечных волокон в сухожилия. Их длина составляет 0,5-1,0 мм, а диаметр – 0,1-0,2 мм. Отдельный нервный аксон несет афферентные импульсы в спинной мозг и называется аксоном Ib. Он начинается в виде веточек, проходящих между коллагеновыми волокнами сухожилия. Когда мышечные волокна сокращаются, коллагеновые волокна сухожилия натягиваются и сжимают нервные веточки, которые начинают импульсировать (рис.2.1). Таким образом, в результате последовательного крепления сухожильных органов к мышечным волокнам они возбуждаются при укорочении возбужденной мышцы. Сухожильные рецепторы возбуждаются в 1,5 – 8 раз более эффективно при мышечном сокращении, нежели при пассивном растяжении.
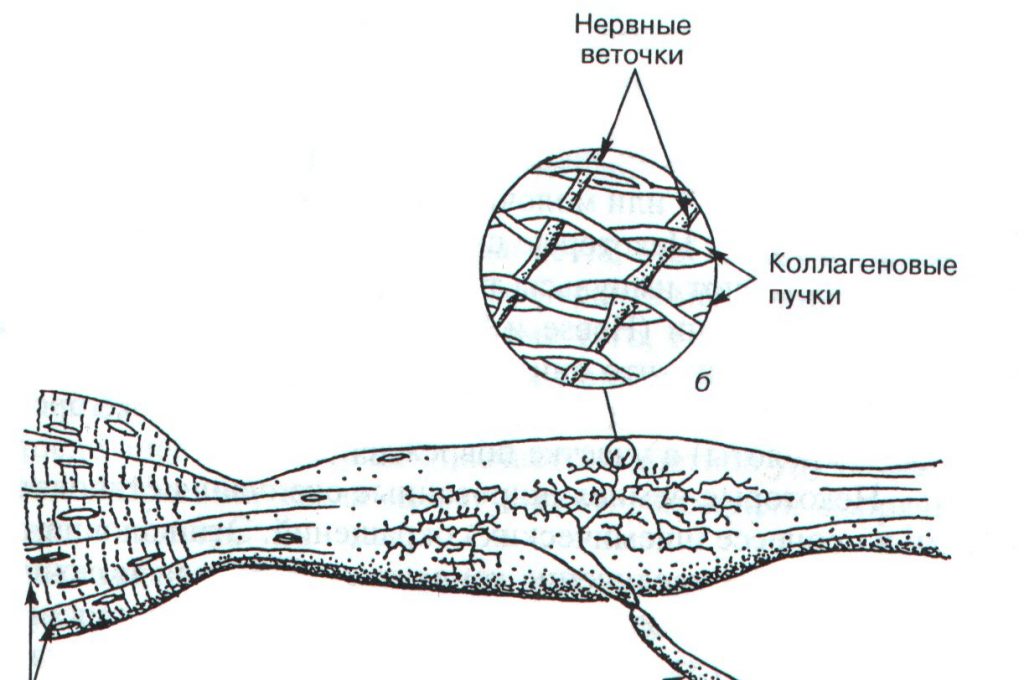
Рис. 2.1. Строение сухожильного органа Гольджи (А.Дж. Мак-Комас, 2001)
Рецепторы суставов
Суставные рецепторы подразделяются на несколько типов в зависимости от их реакции на амплитуду, скорость и направление движения в суставе.
Тельца Руффини находятся в капсуле сустава и воспринимают направление и скорость изменения межзвенного угла. Частота их импульсации возрастает с увеличением скорости изменения суставного угла.
Тельца Паччини посылают в ЦНС информацию о положении отдельных частей тела в пространстве и относительно друг друга. Эти рецепторы посылают в ЦНС информацию о значениях межзвенных углов, то есть о положении сустава. Их импульсация продолжается в течение всего периода сохранения межзвенного угла, и она тем больше, чем больше изменения угла.
2.2. Уровни построения движений и рецепторы опорно-двигательного аппарата
Афферентация уровня А основана на импульсации мышечных веретен (длина и скорость сокращения мышцы) и рецепторов Гольджи (уровень возбуждения мышцы при ее укорочении). Эта информация очень слабо осознается ЦНС, то есть, по гипотезе Н.А.Бернштейна, этот уровень почти никогда не бывает ведущим.
Афферентация уровня В опирается на информацию, поступающую от суставных рецепторов. Это уровень выступает как ведущий в ряде физических упражнений, таких как наклоны тела вперед и назад, а также циклические движения. Сигналы от суставных рецепторов хорошо осознаются.
Рекомендуемая литература
2. Самсонова А.В. Биомеханика мышц: учебно-методическое пособие /А.В.Самсонова, Е.Н. Комиссарова / Под ред А.В.Самсоновой .- СПб., 2008.- 217 с.
3. Самсонова А.В. Моторная и сенсорная функции мышц в биомеханике локомоций: монография /А.В.Самсонова; СПбГУФК им. П.Ф.Лесгафта, 2007.- 152 с.
Проприорецепторы
Необходимым условием нормальной мышечной деятельности является получение информации о положении тела в пространстве и о степени сокращения каждой из мышц. Эта информация поступает в центральную нервную систему от рецепторов вестибулярного аппарата, глаз, кожи, а также от проприорецепторов (мышечно-суставных рецепторов). К проприорецепторам относятся:
- мышечные веретена, находящиеся среди мышечных волокон,
- тельца Гольджи, расположенные в сухожилиях,
- пачиниевы тельца, находящиеся в фасциях, покрывающих мышцы, в сухожилиях, связках и периосте.
Все эти проприорецепторы относятся к группе механорецепторов. Мышечные веретена и тельца Гольджи возбуждаются при растяжении, а пачиниевы тельца — при давлении.
О важности афферентной иннервации мышц свидетельствует и тот факт, что 30-50% волокон в любом нерве, подходящем к мышце, являются афферентными, несущими импульсацию от проприорецепторов. К уже отмечалось, при выключении афферентной иннервации мышц, например при перерезке задних корешков спинного мозга, координированная мышечная деятельность нарушается, несмотря на сохранение двигательной иннервации мышц.
Мышечные веретена представляют собой высокодифференцированные рецепторные образования, снабженные афферентными и эфферентными нервными волокнами. Каждое веретено состоит из нескольких тонких так называемых интрафузальных поперечно-полосатых мышечных волокон ( рис. 195 ).
Oдиночное волокно состоит из центральной части — ядерной сумки и двух способных к сокращению участков. В ядерной сумке pacположены рецепторы, представляющие сообой спиралевидные окончания толстых афферентных покрытых миелиновой оболочкой нервных волокон. Сокращающиеся участки интрафузального волокна иннервированы тонкими моторными, так называемыми гамма-эфферентными, волокнами, образующими мелкие концевые пластинки.
Рис. 195. Схематическое изображение мышечного веретена кролика. 1 — проксимальный конец интрафузального мышечного волокна, прикрепленный к волокну скелетной мышцы; 2 — дистальный конец интрафузального волокна,прикрепленный к фасции; 3 — ядерная сумка; 4 — афферентные волокна, идущие от рецепторов интрафузального волокна; 5 — гамма-эфферентные волокна; 6 — моторное волокно, идущее к скелетной мышце.
Один конец интрафузального волокна прикрепляется к обычному мышечному волокну, а другой — к сухожилию. Таким образом, интрафузальные волокна расположены параллельно массе мышечных волокон, среди которых они находятся. Если мышца растянута или расслаблена, то мышечные веретена тоже растянуты и в рецепторах ядерной сумки возникают импульсы, идущие в центральную нервную систему. Если же мышца сокращена, то натяжение мышечных веретен ослабевает и импульсация прекращается.
Второй вид проприорецепторов — тельца Гольджи — находится в сухожилиях. При мышечном сокращении тельца Гольджи растягиваются и в них возникают нервные импульсы, поступающие к нервным центрам. Сухожильные рецепторы менее возбудимы, чем мышечные веретена.
Таким образом, в двигательном аппарате имеются рецепторы — мышечные веретена,— возбуждающиеся при удлинении (расслаблении и растяжении), и рецепторы — тельца Гольджи,— возбуждающиеся при сокращении мышечных волокон.
Импульсы, поступающие в центральную нервную систему от мышечных веретен, облегчают возникновение рефлекторной реакции данной мышцы и тормозят сокращение мышцы-антагониста. Импульсы, приходящие от сухожильных рецепторов — телец Гольджи, вызывают противоположные рефлекторные реакции ( рис. 180 ).
Импульсация из мышечных веретен, как показал Р. Гранит, может изменяться с помощью специального регуляторного механизма. Такой механизм образован сократительными элементами, расположенными по обе стороны от ядерной сумки интрафузального волокна. Их сокращение вызывает растяжение ядерной сумки и раздражение рецепторов веретена. Таким образом, эти рецепторы могут посылать в центральную нервную систему интенсивную импульсацию даже при небольшом сокращении или расслаблении поперечнополосатой мышцы.
Степень сокращения сократительных элементов веретена регулируется гамма-эфферентными нервными волокнами, которые являются отростками, гамма-мотонейронов спинного мозга. Импульсы, приходящие по гамма-эфферентным волокнам, вызывают сокращения интрафузальных мышечных волокон, что приводит к усилению потока афферентных импульсов от растягиваемых рецепторов ядерной сумки. Активность гамма-мотонейронов регулируется ретикулярной формацией среднего мозга .
Сократительные элементы мышечных веретен всегда находятся в состоянии некоторого тонуса, так как по гамма-эфферентам непрерывно поступают к ним импульсы из центральной нервной системы. Это влечет собой афферентную импульсацию от мышечных веретен, которая в свою ередь тонизирует мотонейроны спинного мозга, что является одной из причин рефлекторного тонуса скелетной мускулатуры. Отличительная особенность проприорецепторов — их малая способность к адаптации, благодаря чему центральная нервная система получает непрерывные сигналы о состоянии скелетной мускулатуры и может осуществлять непрерывную регуляцию двигательных актов.
Кинестетические сигналы, т. е. сигналы о движении отдельной части тела, играют чрезвычайно важную роль в развитии восприятий организма. Они являются главным контролем других органов чувств, например, зрения. Так, оценка зрением расстояния до какого-нибудь предмета вырабатывается при помощи мышечного чувства. Возбуждения, связанные с работой мышц, возникающие при хватании предмета руками или приближении к нему, сочетаются со зрительными ощущениями и служат важнейшим критерием для выработки суждения о том, насколько отдален предмет.
Функция поддержания мышечного тонуса обеспечивается по принципу обратной связи на различных уровнях регуляции организма Периферическая регуляция осуществляется с участием гамма-петли, в состав которой входят супраспинальные моторные пути, вставочные нейроны, нисходящая ретикулярная система, альфа- и гамма-нейроны.
Существует два типа гамма-волокон в передних рогах спинного мозга. Гамма-1-волокна обеспечивают поддержание динамического мышечного тонуса, т.е. тонуса, необходимого для реализации процесса движения. Гамма-2-волокна регулируют статическую иннервацию мышц, т.е. осанку, позу человека. Центральная регуляция функций гамма-петли осуществляется ретикулярной формацией через ретикулоспинальные пути. Основная роль в поддержании и изменении мышечного тонуса отводится функциональному состоянию сегментарной дуги рефлекса растяжения (миотатического, или проприоцептивного рефлекса). Рассмотрим его подробнее.
Рецепторным элементом его является инкапсулированное мышечное веретено. Каждая мышца содержит большое количество этих рецепторов. Мышечное веретено состоит из интрафузальных мышечных волокон (тонких) и ядерной сумки, оплетенной спиралевидной сетью тонких нервных волокон, представляющих собой первичные чувствительные окончания (анулоспинальная нить). На некоторых интрафузальных волокнах имеются также и вторичные, гроздевидные чувствительные окончания. При растяжении интрафузальных мышечных волокон первичные чувствительные окончания усиливают исходящую из них импульсацию, которая через быстропроводящие гамма-1-волокна проводится к альфа-большим мотонейронам спинного мозга. Оттуда, через также быстропроводящие альфа-1-эфферентные волокна, импульс идет к экстрафузальным белым мышечным волокнам, которые обеспечивают быстрое (фазическое) сокращение мышцы. От вторичных чувствительных окончаний, реагирующих на тонус мышцы, афферентная импульсация проводится по тонким гамма-2-волокнам через систему вставочных нейронов к альфа-малым мотонейронам, которые иннервируют тонические экстрафузальные мышечные волокна (красные), обеспечивающие поддержание тонуса и позы.
Интрафузальные волокна иннервируются гамма-нейронами передних рогов спинного мозга. Возбуждение гамма-нейронов, передаваясь по гамма-волокнам к мышечному веретену, сопровождается сокращением полярных отделов интрафузальных волокон и растяжением их экваториальной части, при этом изменяется исходная чувствительность рецепторов к растяжению (происходит снижение порога возбудимости рецепторов растяжения, и усиливается тоническое напряжение мышцы).
Гамма-нейроны находятся под влиянием центральных (супрасегментарных) воздействий, передающихся по волокнам, которые идут от мотонейронов оральных отделов головного мозга в составе пирамидного, ретикулоспинального, вестибулоспинального трактов.
При этом если роль пирамидной системы заключается преимущественно в регуляции фазических (т.е. быстрых, целенаправленных) компонентов произвольных движений, то экстрапирамидная система обеспечивает их плавность, т.е. преимущественно регулирует тоническую иннервацию мышечного аппарата. Так, по мнению J. Noth (1991), спастичность развивается после супраспинального или спинального поражения нисходящих двигательных систем при обязательном вовлечении в процесс кортикоспинального тракта [2] .
В регуляции мышечного тонуса принимают участие и тормозные механизмы, без которых невозможно реципрокное взаимодействие мышц-антагонистов, а значит, невозможно и совершение целенаправленных движений. Они реализуются с помощью рецепторов Гольджи, расположенных в сухожилиях мышц, и вставочных клеток Реншоу, находящихся в передних рогах спинного мозга. Сухожильные рецепторы Гольджи при растяжении или значительном напряжении мышцы посылают афферентные импульсы по быстропроводящим волокнам 1б-типа в спинной мозг и оказывают тормозящее воздействие на мотонейроны передних рогов. Вставочные клетки Реншоу активизируются через коллатерали при возбуждении альфа-мотонейронов, и действуют по принципу отрицательной обратной связи, способствуя торможению их активности. Таким образом, нейрогенные механизмы регуляции мышечного тонуса многообразны и сложны.
При поражении пирамидного пути растормаживается гамма-петля, и любое раздражение путем растяжения мышцы приводит к постоянному патологическому повышению мышечного тонуса. При этом поражение центрального мотонейрона приводит к снижению тормозных влияний на мотонейроны в целом, что повышает их возбудимость, а так же на вставочные нейроны спинного мозга, что способствует увеличению числа импульсов, достигающих альфа-мотонейронов в ответ на растяжение мышцы [4].
В качестве других причин спастичности можно указать структурные изменения на уровне сегментарного аппарата спинного мозга, возникающие вследствие поражения центрального мотонейрона: укорочение дендритов альфа-мотонейронов и коллатеральный спрауттинг (разрастание) афферентных волокон, входящих в состав задних корешков.
Возникают так же и вторичные изменения в мышцах, сухожилиях и суставах. Поэтому страдают механико-эластические характеристики мышечной и соединительной ткани, которые определяют мышечный тонус, что еще больше усиливает двигательные расстройства.
Следствием этого является спастичность мышц, гиперрефлексия, появление патологических рефлексов, а также первоочередная утрата наиболее тонких произвольных движений [5].
Наиболее значимым компонентом мышечного спазма является боль. Болевая импульсация активирует альфа- и гамма-мотонейроны передних рогов, что усиливает спастическое сокращение мышцы, иннервируемой данным сегментом спинного мозга. В то же время, мышечный спазм, возникающий при сенсомоторном рефлексе, усиливает стимуляцию ноцицепторов мышцы. Так, по механизму отрицательной обратной связи формируется замкнутый порочный круг: спазм – боль – спазм – боль [3].
Интерес представляют также данные о центральных холинергических механизмах регуляции мышечного тонуса. Показано, что клетки Реншоу активируются ацетилхолином как через коллатерали мотонейрона, так и через ретикулоспинальную систему.
M.Schieppati и соавт., (1989) установили, что фармакологическая активация центральных холинергических систем значительно снижает возбудимость альфа-мотонейронов путем повышения активности клеток Реншоу.
В последние годы исследователи регуляции мышечного тонуса придают огромное значение роли нисходящих адренергических супраспинальных путей, начинающихся в области голубого пятна. Анатомически эти образования тесно связаны со спинальными структурами, особенно с передними рогами спинного мозга. Норадреналин, высвобождаемый с терминалей бульбоспинальных волокон, активизирует адренорецепторы, располагающиеся во вставочных нейронах, первичных афферентных терминалях и мотонейронах и воздействует одновременно на альфа- и бета-адренорецепторы в спинном мозге (D.Jones et al., 1982). К ядерным образованиям ретикулярной формации ствола подходят многочисленные аксоны болевой чувствительности. На основе информации, поступающей в ретикулярную формацию ствола головного мозга, выстраиваются соматические и висцеральные рефлексы. От ядерных образований ретикулярной формации формируются связи с таламусом, гипоталамусом, базальными ядрами и лимбической системой, которые обеспечивают реализацию нейроэндокринных и аффективных проявлений боли, что особенно важно при хронических болевых синдромах [2].
В итоге формирующийся порочный круг включает в себя мышечный спазм, боль, локальную ишемию, дегенеративные изменения, которые самоподдерживают друг друга, усиливая первопричину патологических изменений.
Следует учитывать, что чем больше компонентов этого порочного круга становятся мишенями при лечении, тем выше вероятность его успеха. Поэтому современными требованиями к миорелаксирующей терапии являются: мощность миорелаксирующего действия, его селективность, наличие противосудорожного и антиклонического эффектов, мощность анальгетического действия, а так же безопасность и наличие широкого терапевтического диапазона доз препарата.
Согласно современным представлениям, большинство миорелаксантов воздействуют на трансмиттеры или нейромодуляторы ЦНС. Воздействие может включать супрессию возбуждающих медиаторов (аспартат и глутамат) и/или усиление тормозных процессов (ГАМК, глицин). Далее
Тормозная система клеток Реншоу. Мышечные сенсорные рецепторы
а) Тормозная система клеток Реншоу. В передних рогах спинного мозга расположено большое количество небольших нейронов, называемых клетками Реншоу, тесно связанных с мотонейронами. Как только аксон переднего мотонейрона покидает тело клетки, его коллатерали идут к прилежащим клеткам Реншоу. Это тормозные клетки, проводящие тормозные сигналы к окружающим мотонейронам. Таким образом, стимуляция каждого мотонейрона ведет к торможению прилежащих мотонейронов.
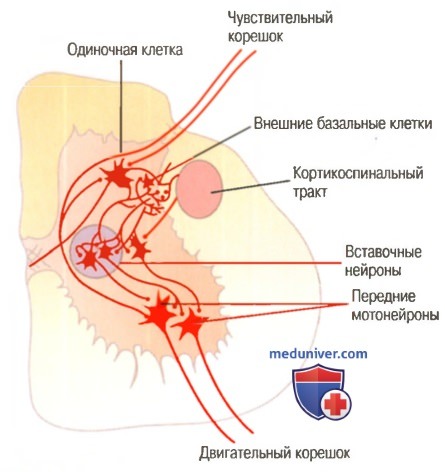
Связи периферических сенсорных и кортикоспинальных волокон со вставочными нейронами и передними мотонейронами спинного мозга
б) Множественные межсегментарные связи спинного мозга. Проприоспинальные волокна. Более половины восходящих и нисходящих нервных волокон спинного мозга являются проприоспиналъными волокна ми. Они проходят от одного сегмента спинного мозга к другому. Кроме того, когда сенсорные волокна входят в спинной мозг по его задним корешкам, они разветвляются, и ветви идут вверх и вниз вдоль спинного мозга; некоторые из них проводят сигналы лишь к одному или двум сегментам, тогда как другие проводят сигналы ко многим сегментам.
Эти восходящие и нисходящие проприоспинальные волокна обеспечивают пути для мультисегментарных рефлексов, изложенных в этой главе далее, включая рефлексы, координирующие одновременные движения передних и задних конечностей.
Мышечные сенсорные рецепторы: мышечные рецепторы и сухожильные рецепторы Гольджи. Их роль в регуляции мышечных сокращений
Для надлежащей регуляции мышечной функции требуется не только возбуждение мышцы мотонейронами передних рогов спинного мозга. Необходима также постоянная информация по принципу обратной связи между мышцей и спинным мозгом о функциональном состоянии каждой мышцы в данный момент: длине мышцы, ее напряжении, скорости изменения ее длины и напряжения в каждое мгновение.
Эту информацию обеспечивают два особых типа рецепторов, представленных в мышцах и их сухожилиях:
(1) мышечные веретена, которые распределены по всему протяжению брюшка мышцы и посылают информацию в нервную систему о длине мышцы или скорости ее изменения) (для облегчения понимания просим вас изучить первый рисунок ниже);
(2) сухожильные органы Гольджи, которые локализуются в мышечных сухожилиях и проводят информацию о напряжении сухожилия или скорости его изменения (для облегчения понимания просим вас изучить второй рисунок ниже).
Мышечное веретено, его связи с крупными экстрафузальными мышечными волокнами. Обратите внимание также на моторную и сенсорную иннервации мышечного веретена Сухожильный орган Гольджи
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Читайте также: