
Как и при каких условиях гликолиз может преобладать над аэробным процессом кратко
Обновлено: 04.07.2024
Гликолиз (от греч. glycys – сладкий и lysis – растворение, распад) – это последовательность ферментативных реакций, приводящих к превращению глюкозы в пируват с одновременным образованием АТФ.
При аэробных условиях пируват проникает в митохондрии, где полностью окисляется до СО2 и Н2О. Если содержание кислорода недостаточно, как это может иметь место в активно сокращающейся мышце, пируват превращается в лактат.
Итак, гликолиз – не только главный путь утилизации глюкозы в клетках, но и уникальный путь, поскольку он может использовать кислород, если
последний доступен (аэробные условия), но может протекать и в отсутствие кислорода (анаэробные условия).
Анаэробный гликолиз – сложный ферментативный процесс распада глюкозы, протекающий в тканях человека и животных без потребления кислорода. Конечным продуктом гликолиза является молочная кислота. В процессе гликолиза образуется АТФ. Суммарное уравнение гликолиза можно представить следующим образом:

В анаэробных условиях гликолиз – единственный процесс в животном организме, поставляющий энергию. Именно благодаря гликолизу организм человека и животных определенный период может осуществлять ряд физиологических функций в условиях недостаточности кислорода. В тех случаях, когда гликолиз протекает в присутствии кислорода, говорят об аэробном гликолизе .
Последовательность реакций анаэробного гликолиза, так же как и их промежуточные продукты, хорошо изучена. Процесс гликолиза катализируется одиннадцатью ферментами, большинство из которых выделено в гомогенном, клисталлическом или высокоочищенном виде и свойства которых достаточно известны. Заметим, что гликолиз протекает в гиало-плазме (цитозоле) клетки.
Первой ферментативной реакцией гликолиза является фосфорилирование, т.е. перенос остатка ортофосфата на глюкозу за счет АТФ. Реакция катализируется ферментом гексокиназой:
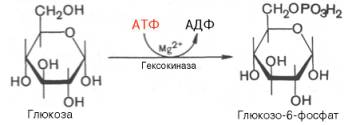
Образование глюкозо-6-фосфата в гексокиназной реакции сопровождается освобождением значительного количества свободной энергии системы и может считаться практически необратимым процессом.
Наиболее важным свойством гексокиназы является ее ингибирование глюкозо-6-фосфатом, т.е. последний служит одновременно и продуктом реакции, и аллостерическим ингибитором.
Фермент гексокиназа способен катализировать фосфорилирование не только D-глюкозы, но и других гексоз, в частности D-фруктозы, D-маннозы и т.д. В печени, кроме гексокиназы, существует фермент глюкокиназа, который катализирует фосфорилирование только D-глюкозы. В мышечной ткани этот фермент отсутствует (подробнее см. главу 16).
Второй реакцией гликолиза является превращение глюкозо-6-фос-фата под действием фермента глюкозо-6-фосфатизомеразы во фруктозо-6-фосфат:
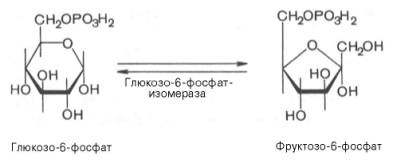
Эта реакция протекает легко в обоих направлениях, и для нее не требуется каких-либо кофакторов.
Третья реакция катализируется ферментом фосфофруктокиназой; образовавшийся фруктозо-6-фосфат вновь фосфорилируется за счет второй молекулы АТФ:
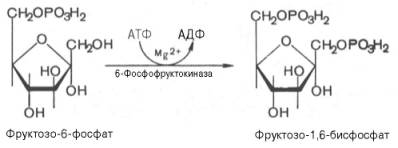
Данная реакция аналогично гексокиназной практически необратима, протекает в присутствии ионов магния и является наиболее медленно текущей реакцией гликолиза. Фактически эта реакция определяет скорость гликолиза в целом.
Фосфофруктокиназа относится к числу аллостерических ферментов. Она ингибируется АТФ и стимулируется АМФ . При значительных величинах отношения АТФ/АМФ активность фосфофруктокиназы угнетается и гликолиз замедляется. Напротив, при снижении этого коэффициента интенсивность гликолиза повышается. Так, в неработающей мышце активность фосфофруктокиназы низкая, а концентрация АТФ относительно высокая. Во время работы мышцы происходит интенсивное потребление АТФ и активность фосфофруктокиназы повышается, что приводит к усилению процесса гликолиза.
Четвертую реакцию гликолиза катализирует фермент альдолаза. Под влиянием этого фермента фруктозо-1,6-бисфосфат расщепляется на две фосфотриозы:
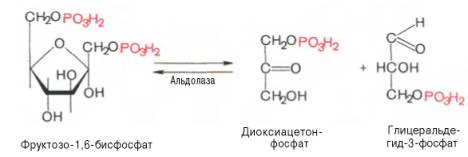
Эта реакция обратима. В зависимости от температуры равновесие устанавливается на различном уровне. При повышении температуры реакция сдвигается в сторону большего образования триозофосфатов (дигидро-ксиацетонфосфата и глицеральдегид-3-фосфата).
Пятая реакция – это реакция изомеризации триозофосфатов. Катализируется ферментом триозофосфатизомеразой:

Равновесие данной изомеразной реакции сдвинуто в сторону дигид-роксиацетонфосфата: 95% дигидроксиацетонфосфата и около 5% глице-ральдегид-3-фосфата. В последующие реакции гликолиза может непосредственно включаться только один из двух образующихся триозофосфатов, а именно глицеральдегид-3-фосфат. Вследствие этого по мере потребления в ходе дальнейших превращений альдегидной формы фосфотриозы ди-гидроксиацетонфосфат превращается в глицеральдегид-3-фосфат.
Образованием глицеральдегид-3-фосфата как бы завершается первая стадия гликолиза. Вторая стадия – наиболее сложная и важная. Она включает окислительно-восстановительную реакцию (реакция гликолитической оксидоредукции), сопряженную с субстратным фосфорилированием, в процессе которого образуется АТФ.
В результате шестой реакции глицеральдегид-3-фосфат в присутствии фермента глицеральдегидфосфатдегидрогеназы, кофермента НАД и неорганического фосфата подвергается своеобразному окислению с образованием 1,3-бисфосфоглицериновой кислоты и восстановленной формы НАД (НАДН). Эта реакция блокируется йод- или бромацетатом, протекает в несколько этапов:
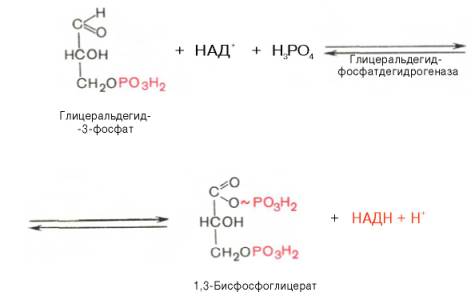
Седьмая реакция катализируется фосфоглицераткиназой, при этом происходит передача богатого энергией фосфатного остатка (фосфатной группы в положении 1) на АДФ с образованием АТФ и 3-фосфогли-цериновой кислоты (3-фосфоглицерат):
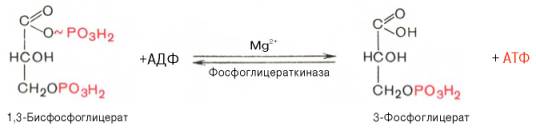
Таким образом, благодаря действию двух ферментов (глицеральде-гидфосфатдегидрогеназы и фосфоглицераткиназы) энергия, высвобождающаяся при окислении альдегидной группы глицеральдегид-3-фосфата до карбоксильной группы, запасается в форме энергии АТФ. В отличие от окислительного фосфорилирования образование АТФ из высокоэнергетических соединений называется субстратным фосфорилированием.
Восьмая реакция сопровождается внутримолекулярным переносом оставшейся фосфатной группы, и 3-фосфоглицериновая кислота превращается в 2-фосфоглицериновую кислоту (2-фосфоглицерат).
Реакция легкообратима, протекает в присутствии ионов Mg 2+ . Кофактором фермента является также 2,3-бисфосфоглицериновая кислота аналогично тому, как в фосфоглюкомутазной реакции роль кофактора выполняет глюкозо-1,6-бисфосфат:

Девятая реакция катализируется ферментом енолазой, при этом 2-фосфоглицериновая кислота в результате отщепления молекулы воды переходит в фосфоенолпировиноградную кислоту (фосфоенолпируват), а фосфатная связь в положении 2 становится высокоэргической:
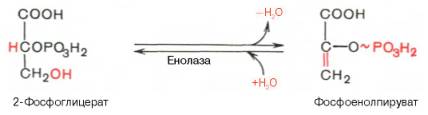
Енолаза активируется двухвалентными катионами Mg 2+ или Мn 2+ и ингибируется фторидом.
Десятая реакция характеризуется разрывом высокоэргической связи и переносом фосфатного остатка от фосфоенолпирувата на АДФ (субстратное фосфорилирование). Катализируется ферментом пируваткиназой:
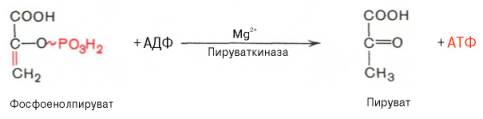
Для действия пируваткиназы необходимы ионы Mg 2+ , а также одновалентные катионы щелочных металлов (К + или др.). Внутри клетки реакция является практически необратимой.
В результате одиннадцатой реакции происходит восстановление пировиноградной кислоты и образуется молочная кислота. Реакция протекает при участии фермента лактатдегидрогеназы и кофермента НАДН, образовавшегося в шестой реакции:

Последовательность протекающих при гликолизе реакций представлена на рис. 10.3.
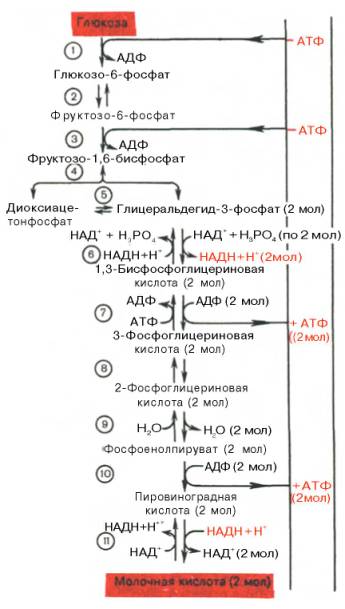
Рис. 10.3. Последовательность реакций гликолиза.
1 - гексокиназа; 2 - фосфоглюкоизоме-раза; 3 - фосфофруктокиназа; 4 - альдо-лаза; 5 - триозофосфатизомераза; 6 - гли-церальдегидфосфатдегидрогеназа; 7 -фосфоглицераткиназа; 8 - фосфоглицеромутаза; 9 - енолаза; 10 - пируватки-наза; 11 - лактатдегидрогеназа.
Реакция восстановления пирувата завершает внутренний окислительно-восстановительный цикл гликолиза. НАД + при этом играет роль промежуточного переносчика водорода от глицеральдегид-3-фосфата (6-я реакция) на пировиноградную кислоту (11-я реакция), при этом сам он регенерируется и вновь может участвовать в циклическом процессе, получившем название гликолитический оксидоредукции.
Биологическое значение процесса гликолиза заключается прежде всего в образовании богатых энергией фосфорных соединений. На первых стадиях гликолиза затрачиваются 2 молекулы АТФ (гексокиназная и фосфофрук-токиназная реакции). На последующих образуются 4 молекулы АТФ (фосфоглицераткиназная и пируваткиназная реакции). Таким образом, энергетическая эффективность гликолиза в анаэробных условиях составляет 2 молекулы АТФ на одну молекулу глюкозы.
Как отмечалось, основной реакцией, лимитирующей скорость гликолиза, является фосфофруктокиназная. Вторая реакция, лимитирующая скорость и регулирующая гликолиз – гексокиназная реакция. Кроме того, контроль гликолиза осуществляется также ЛДГ и ее изоферментами.
В тканях с аэробным метаболизмом (ткани сердца, почек и др.) преобладают изоферменты ЛДГ1 и ЛДГ2 (см. главу 4). Эти изоферменты инги-бируются даже небольшими концентрациями пирувата, что препятствует образованию молочной кислоты и способствует более полному окислению пирувата (точнее, ацетил-КоА) в цикле трикарбоновых кислот.
В тканях человека, в значительной степени использующих энергию гликолиза (например, скелетные мышцы), главными изоферментами являются ЛДГ5 и ЛДГ4. Активность ЛДГ5 максимальна при тех концентрациях пирувата, которые ингибируют ЛДГ1. Преобладание изоферментов ЛДГ4 и ЛДГ5 обусловливает интенсивный анаэробный гликолиз с быстрым превращением пирувата в молочную кислоту.
Как отмечалось, процесс анаэробного распада гликогена получил название гликогенолиза. Вовлечение D-глюкозных единиц гликогена в процесс гликолиза происходит при участии 2 ферментов – фосфорилазы а и фосфо-глюкомутазы. Образовавшийся в результате фосфоглюкомутазной реакции глюкозо-6-фосфат может включаться в процесс гликолиза. После образования глюкозо-6-фосфата дальнейшие пути гликолиза и гликогенолиза полностью совпадают:
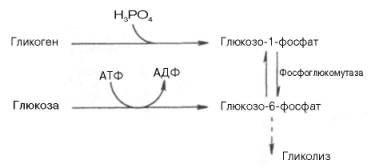
В процессе гликогенолиза в виде макроэргических соединений накапливаются не две, а три молекулы АТФ (АТФ не тратится на образование глюкозо-6-фосфата). Кажется, что энергетическая эффективность глико-генолиза выглядит несколько более высокой по сравнению с процессом гликолиза, но эта эффективность реализуется только при наличии активной фосфорилазы а. Следует иметь в виду, что в процессе активации фосфо-рилазы b расходуется АТФ (см. рис. 10.2).
Гликолиз - процесс окисления глюкозы, при котором из одной молекулы глюкозы образуются две молекулы пировиноградной кислоты, не является мембранозависимым процессом. Он происходит в цитоплазме. Однако ферменты гликолиза связаны со структурами цитоскелета. Суть гликолиза состоит в том, что молекула глюкозы (C6H12O6) без участия кислорода распадается на две молекулы пировиноградной кислоты (СН3СОСООН). При этом окисление идет за счет отщепления от молекулы глюкозы четырех атомов водорода, связывающихся со сложным органическим веществом НАД с получением двух молекул НАД•Н. Выделяющаяся при этом энергия запасается (40% от общего количества) в виде макроэргических связей двух молекул АТФ. 60% энергии выделяется в виде тепла. При последующем окислении НАД•Н получается еще 6 молекул АТФ. Таким образом, полный энергетический выход гликолиза в анаэробных условиях составляет 8 молекул АТФ.
Аэробный процесс гликолиза (10 реакций), уравнение (с образованием пирувата):
Анаэробный процесс гликолиза (11 реакций), уравнение (с образованием лактата):
Схема процесса гликолиза и его реакции

На схеме в рамках обозначены исходные субстраты и конечные продукты гликолиза, цифрами в скобках - число молекул.
ATP (АТФ) - это аденозинтрифосфорная кислота, универсальный источник энергии
ADP (АДФ) - это аденозиндифосфат, нуклеотид, участвует в энергетическом обмене
NAD (НАД) - никотинамидадениндинуклеотидфосфата
NADH (НАД•Н) - востановленная форма NAD
Таблица процесс гликолиза его реакции
Для распада и частичного окисления молекулы глюкозы требуется протекание 11 сложных последовательных реакций.
Гликолиз – это процесс превращения глюкозы в пируват (10 реакций).
Считается вторым этапом катаболизма (второй называется унификацией) в обмене углеводов – см. п.20.
Локализация процесса – гликолиз протекает во всех клетках организма, в гиалоплазме.
Глюкоза для гликолиза и других процессов поступает в клетки из крови, для чего в крови должна поддерживаться достаточная концентрация глюкозы (от 3 ммоль/л – см. п.37).
Транспорт глюкозы через мембраны клеток осуществляется специальными белками-переносчиками глюкозы. Транспорт глюкозы в клетки многих тканей (например, жировой и мышечной тканей, но не нейронов и эритроцитов) стимулируется инсулином (см. п.102, 103); и поэтому при снижении количества инсулина в крови или снижении действия инсулина (при диабете) транспорт глюкозы в клетки названных тканей снижен, что приводит к дефициту глюкозы в клетках и к избытку глюкозы в крови – см. п.37.
Кроме того, при расщеплении гликогена в мышцах (гликогенолизе – см.п.31) образуется метаболит гликолиза – глюкозо-6-фосфат, который сразу вступает во вторую реакцию гликолиза.
32.2. Конечные метаболиты гликолиза, значение и тканевые особенности
(то есть в каких клетках и тканях какие варианты бывают).
Центральным конечным метаболитом гликолиза является ПИРУВАТ (2 молекулы пирувата образуются из одной молекулы глюкозы).
Дальнейшие превращения пирувата зависят от условий, в которых протекает гликолиз – или в лактат, или в ацетилКоА, или в оксалоацетат (см. далее).
32.2.1. Превращения пирувата В АЭРОБНЫХ УСЛОВИЯХ
(АЭРОБНЫЙ ОБМЕН УГЛЕВОДОВ:
Пируват ; ацетилКоА + 2,5 АТФ
2.1 – АцетилКоА может поступить в ЦТК, что может привести к выработке 10 молекул АТФ от каждой молекулы ацетилКоА. И превращению ацетилКоА в углекислый газ для выдоха. Что завершает распад углеводородного скелета молекулы глюкозы.
ацетилКоА ; 2СО2 + 10 АТФ (при поступлении в ЦТК и ДЦ)
2.2 – В печени ацетилКоА может использоваться для синтеза жирных кислот (при сытости, см. п.46),кетоновых тел (при голоде и стрессе, см. п.47) и холестерина (см. п.49), а также (через реакции ЦТК) – аминокислот глутамата, глутамина и т.д. (см. п.67).
ацетилКоА ; жирные кислоты, кетоновые тела, холестерин, аминокислоты (с оксалоацетатом)
3 – Из пирувата может образоваться аминокислота АЛАНИН (см. п.64, 67 и 33) и кетокислота ОКСАЛОАЦЕТАТ (см. п.33), которая может 1) использоваться в ЦТК (см.п. 21) и 2) для синтеза аминокислоты аспартата для синтеза аспарагина, белков, для обезвреживания аммиака в синтезе мочевины (см.п.66) и для синтеза нуклеотидов (а затем РНК и ДНК).
4 – из метаболита гликолиза 3-фосфоглицерата (см. п.67) может синтезироваться аминокислота СЕРИН, из которой могут синтезироваться глицин, цистеин, липоиды (см. п.52).
Пируват ; аланин или оксалоацетат (; аспартат ; мочевина или нуклеотиды ; ДНК и РНК)
ОКСАЛОАЦЕТАТ для ЦТК или ; аспартат ; мочевина или нуклеотиды ; ДНК и РНК)
; (биотин!)
Аланин ; ПИРУВАТ ; (ПДГ) АЦЕТИЛ-КоА (+ 2,5 АТФ) ; (в ЦТК) 2СО2 + 10 АТФ (всего 12,5 АТФ)
; (под действием ПДГ, витамины – В1, В2, РР, пантотенат, липоевая)
ацетилКоА ; жирные кислоты (для жира и липоидов), кетоновые тела, холестерин
ЗНАЧЕНИЕ ГЛИКОЛИЗА в аэробных условиях – в получении всех перечисленных веществ, в том числе для синтезов (можно сказать, что это анаболическая роль гликолиза по аналогии с анаэробной ролью ЦТК). Участие гликолиза в выработке АТФ – это энергетическая роль гликолиза. Участие гликолиза в катаболизме глюкозы (превращение 6-тиуглеродной молекулы глюкозы в две трёхуглеродных молекулы пирувата) и поставка метаболитов для ЦТК (ацетилКоА и оксалоацетата) – катаболическая. Это достоинства гликолиза в аэробных условиях.
Недостатком аэробного обмена глюкозы является то, что его реакции протекают только в аэробных условиях и не протекают в анаэробных.
Использовать кислород в дыхательной цепи не могут ЭРИТРОЦИТЫ из-за отсутствия в них митохондрий и дыхательной цепи,
а также любые клетки, если в них не работает дыхательная цепь (см.п.22) из-за попадания в них ингибиторов дыхательной цепи.
Дефицит кислорода (гипоксия) возникает во всех клетках при нарушении доставки кислорода:
1) из-за нарушения кровотока (из-за тромбов, жгута, сдавливания, падения артериального давления, большой кровопотери, сердечной недостаточности и т.д.),
2) из-за нарушения работы эритроцитов (см. п.121),
3) из-за нарушения поступления кислорода в лёгкие (при эмфиземе лёгких у курильщиков, при бронхоспазме и т.д.).
Превращение глюкозы в лактат в анаэробных условиях могут называть анаэробным гликолизом.
Лактат опасен, поскольку его накопление приводит к АЦИДОЗУ, который называется лактацидозом (в отличие от кетоацидоза при накоплении кетоновых тел – см. п. 47). Лактацидоз и кетоацидоз относятся к метаболическим ацидозам, то есть к ацидозам, обусловленным накоплением метаболитов (кислот) и нарушением метаболизма. Есть ещё респираторный ацидоз – то есть связанный с нарушением дыхания (задержкой дыхания). При всех ацидозах портится самочувствие, нарушается работа белков (так как оптимум для большинства белков – рН около 7), возникает чувство тошноты. Сильный ацидоз может привести к коме, но для здоровых людей это не характерно.
32.3. ОБМЕН ЛАКТАТА.
Это процессы образования и использования лактата.
32.3.1. ОБРАЗОВАНИЕ лактата.
Как сказано выше, образуется лактат при анаэробном гликолизе в эритроцитах, мышцах и других клетках в анаэробных условиях. Из клеток он может поступать в кровь, что при избытке лактата приводит к ацидозу.
Ткани-клетки-источники лактата:
В покое основное количество лактата поступает в кровь из эритроцитов, а при наряжённой работе мышц – из мышц.
Кроме этого, лактат поступает в кровь из любых клеток, которым не хватает кислорода или в которых не работают дыхательная цепь, ЦТК и ПДГ. Причиной сниженной работы ЦТК бывает дефицит витаминов ЦТК, особенно В1. См. 32.2.2.
32.3.2. Использование и УТИЛИЗАЦИЯ ЛАКТАТА.
Из крови большая часть лактата поступает в клетки печени, где утилизируется. Это снижает ацидоз. Поэтому нарушение работы печени (печёночная недостаточность) может стать причиной избытка лактата в крови (из-за сниженного использования лактата клетками печени).
Пути метаболизма лактата – 25% и 75%.
25% лактата подвергаются так называемому ОКИСЛЕНИЮ ЛАКТАТА.
При этом:
1) лактат превращается в пируват под действием фермента ЛДГ, что даёт 2,5 АТФ благодаря образованию НАДН и его использованию в дыхательной цепи,
2) затем пируват превращается в ацетилКоА под действием ПДГ, что даёт ещё 2,5 АТФ благодаря образованию НАДН и его использованию в дыхательной цепи,
3) затем ацетилКоА вступает в ЦТК, что даёт ещё 10 АТФ.
В сумме окисление одной молекулы лактата даёт 15 молекул АТФ.
Реакции окисления лактата идут только в аэробных условиях.
75% лактата используются для синтеза глюкозы, который называется глюконеогенезом (ГНГ) и протекает в основном в печени.
Поэтому снижение ГНГ может стать причиной избытка лактата и ацидоза.
При превращении лактата в глюкозу:
1) сначала лактат превращается в пируват под действием ЛДГ, как и при окислении лактата,
2) затем пируват превращается в ОКСАЛОАЦЕТАТ (см. п.33 и п.21), что требует витамина БИОТИН – без биотина реакция не идёт (причиной дефицита биотина, нарушения этой реакции, ГНГ и избытка лактата бывает избыточное употребление сырых яиц, так как в них содержится вещество, препятствующее усвоению биотина),
3) оксалоацетат превращается в глюкозу в ходе реакций глюконеогенеза.
32.3.3. Избыток лактата.
Между процессами образования лактата и его использования должно быть равновесие, благодаря которому концентрация лактата в клетках и крови не превышала бы нормальную.
Повышение концентрации лактата в клетках приводит к снижению рН в клетках, нарушению работы белков клеток, повреждению клеток. Например, мышцы при избыточной концентрации лактата теряют способность сокращаться.
Повышение концентрации лактата в крови называется гиперЛАКТАТемией и приводит к ацидозу.
К избытку лактата приводит или повышение образования лактата, или снижение утилизация лактата.
Причины повышенного образования лактата (см. выше 32.2.3. и 32.3) –
1) дефицит кислорода (гипоксия при анемии, сердечной или лёгочной недостаточности и т.д.) или снижение его использования,
2) снижение превращения пирувата в ацетилКоА из-за нарушения работы ПДГ при дефиците В1 или других витаминов ПДГ,
3) повышенная концентрация НАДН, которая бывает не только при дефиците кислорода, но и при употреблении алкоголя.
Причины сниженного использования лактата и пирувата:
1) дефицит витаминов ПДГ и биотина, без которых не протекают реакции использования и пирувата,
2) нарушение работы печени, в котором должен протекать ГНГ.
Для коррекции ацидоза, обусловленного избытком лактата, нужно убрать причины избытка лактата – восстановить кровоток, принимать витамины, не употреблять алкоголь и т.д.
32.5. Достоинства и недостатки анаэробного гликолиза.
Таким образом, то, что анаэробный гликолиз даёт хоть немного АТФ без кислорода – это его ДОСТОИНСТВО.
А то, что этого количества АТФ МАЛО (хватает только эритроцитам, тканям конечностей при гипоксии и скелетным мышцам при короткой напряжённой работе, но не хватает другим тканям вообще, а тканям конечностей не хватает более чем на час, не хватает мышцам для длительной работы – это НЕДОСТАТОК анаэробного гликолиза).
Ещё один недостаток анаэробного гликолиза – то, что избыток его конечного метаболита (лактата) приводит к ацидозу.
Гликолиз является единственным процессом, за счёт которого может образоваться АТФ при отсутствии (или неиспользовании) кислорода. Именно гликолиз обеспечивает молекулами АТФ эритроциты, в которых нет других источников АТФ из-за отсутствия митохондрий и дыхательной цепи см. п.23.
32.7. РЕГУЛЯЦИЯ ГЛИКОЛИЗА. См. п.6 и 7, 33, 37, 102.
Смысл регуляции любого процесса в том, чтобы его скорость соответствовала потребностям организма и отдельных клеток.
Изменение скорости процесса осуществляется через изменение активности или концентрации ферментов процесса под влиянием метаболитов и гормонов.
Активность повышается активаторами и снижается ингибиторами.
Концентрация ферментов повышается в результате усиления синтеза ферментов под влиянием индукторов и снижается в результате снижения синтеза под влиянием репрессоров.
Регулировать все ферменты 10 реакций гликолиза не обязательно. Достаточно регулировать активность некоторых ферментов. Эти регулируемые ферменты называются КЛЮЧЕВЫМИ – см. п.7.
В гликолизе ключевыми ферментами являются три – ферменты первой реакции, последней и третьей – гексокиназа, фосфофруктокиназа и пируваткиназа.
Фермент первой реакции последовательности реакций часто является ключевым. Фермент третьей реакции гликолиза является самым медленным – это тоже типично (что фермент самой медленной реакции метаболического пути является ключевым).
32.7.1. Регуляция гликолиза гормонами.
Гликолиз стимулируется инсулином (за счёт индукции ключевых ферментов) и подавляется глюкокортикостероидами (ГКС; за счёт репрессии ключевых ферментов). См. п.85.
Катехоламины адреналин и норадреналин усиливают гликолиз в мышцах и подавляют в печени.
Благодаря стимуляции инсулином гликолиза (а также стимуляции инсулином транспорта глюкозы во многие клетки) расход глюкозы тканями увеличивается, что приводит
1) к удовлетворению потребностей клеток в глюкозе и
2) к снижению концентрации глюкозы в крови –
это снижение концентрации глюкозы в крови называется гипогликемическим эффектом,
а инсулин, который обусловливает снижение концентрации глюкозы в крови, является гипогликемическим гормоном.
32.7.2. Регуляция гликолиза метаболитами.
Обычно активаторами процесса являются его субстраты, а ингибиторами – продуктами. Механизм – аллостерический (см. п.6).
Активатором гликолиза является АДФ (в качестве субстрата – чтобы превратиться в АТФ), а ингибиторами – продукты гликолиза АТФ, ацетилКоА и цитрат (отдалённо это продукты гликолиза, так как цитрат образуется из ацетилКоА и оксалоацетата, образующихся из пирувата).
Эффекты АДФ противоположны эффектам АТФ, так как АДФ образуется при расщеплении АТФ, и [АДФ] становится тем больше, чем меньше [АТФ].
Ещё один активатор гликолиза – метаболит фруктозо-2,6- бисфосфат, который активирует фосфофруктокиназу.
Сравните регуляцию гликолиза с регуляцией ГНГ – эти два процесса регулируются одними и теми же регуляторами, но действие регуляторов на эти два противоположных процесса прямо противоположно. Это помогает рассказать регуляцию ГНГ, зная регуляцию гликолиза.
Р е г у л я ц и я г о р м о н а м и процессов углеводного обмена.
Усиление процесса
способствует (гипер/гликемии) или не способствует? Г о р м о н ы
Ин ГГ КА ГКС
; [Г] Гипергликемические
Гликолиз (глюкоза в пируват) Нет, ведёт к гипогликемии ; ; в печени см. *
Глюконеогенез (синтез глюкозы) Да, ведёт к гипергликемии ; ; в печени
* В мышцах КА (в т.ч. адреналин) не снижают гликолиз, а усиливают – так как КА должны обеспечить мышцы энергией за счет выработки в мышцах АТФ. Гликолиз – это единственный процесс, который может дать АТФ при дефиците кислорода (дефицит кислорода обязательно возникает в работающих скелетных мышцах, как бы ни старались легкие и сердце обеспечить мышцы кислородом; но чем лучше человек тренирован, тем лучше организм справляется с нагрузками).
Р е г у л я ц и я гликолиза и ГНГ метаболитами. (Аллостерическая).
(Действие регуляторов на гликолиз и на ГНГ противоположно.)
ацетилКоА Цитрат А Т Ф А Д Ф Фруктозо-2,6-бисфосфат
гликолиз Продукт продукт Продукт Субстрат
Ингибиторы гликолиза Активаторы гликолиза
ГНГ: синтез
глюкозы Субстрат продукт
Активаторы ГНГ Ингибиторы ГНГ
12
В чем д о с т о и н с т в а
(см. выше) 1) только он дает АТФ в ЭЦ,
2) только он дает АТФ без О2, хоть и мало (всего 2),
3) быстро включается, что ценно при короткой напряженной работе 1) дает много АТФ,
2) дает метаболиты ЦТК
(ОА и ацетилКоА из П,
а затем в ЦТК образуются остальные, в т.ч. для аминокислот и белков)
В чем н е д о с т а т к и
(см. выше) 1) образует лактат, что приводит к ацидозу,
2) мало АТФ (из-за чего жгут нельзя долго держать) не протекает
без кислорода
(при гипоксии)
14 Каким
тканям или клеткам
дает достаточное количество энергии Эритроцитам, скелетным мышцам при короткой работе, а при жгуте – час Всем, в которых протекает
(в эритроцитах не протекает)
О б м е н л а к т а т а . Лактат (Л) – это соль или анион молочной кислоты.
7.
Причины накопления лактата
(гипер/лактат/емии),
связанные
с увеличением образования лактата
(см. 3 и 4) У здоровых – работа скелетных мышц (усталость, тошнота).
У больных – разные формы Г И П О К С И И , в т.ч:
1. из-за ишемии (жгут, тромб, спазм, падение АД и т.д.),
2. из-за патологии дыхательной системы (эмфизема легких при курении и др.),
3. из-за патологии сердечно-сосудистой системы,
4. из-за анемии (нет Fe, витаминов, белка, есть гемолиз),
5. при шоке, коллапсе, терминальных состояниях.
8. Причины накопления лактата,
связанные с уменьшением его утилизации (использования) 1) патология печени из-за снижения ГНГ
(п/л печени – из-за алкоголизма и др.),
2) дефицит В1, т.к. без него не может работать ПДГ и ЦТК.
Как и при каких условиях гликолиз может преобладать над аэробным процессом?

Анаэробный процесс расщепления углеводов может начинаться с гликогена — гликогенолиз — или с глюкозы— гликолиз.
Конечным продуктом анаэробного распада углеводов является молочная кислота.

Где происходит аэробный этап гликолиза а) в митохондриях б) в легких с)в пищеварительном канале г) в цитоплазме?
Где происходит аэробный этап гликолиза а) в митохондриях б) в легких с)в пищеварительном канале г) в цитоплазме.

Где происходит процесс гликолиза?
Где происходит процесс гликолиза?

Как называется этап энергетического обмена, в котором участвует кислород?
Как называется этап энергетического обмена, в котором участвует кислород?
Гликолиз Подготовительный Аэробное дыхание.

Что происходит с глюкозой в процессе гликолиза?
Что происходит с глюкозой в процессе гликолиза.
Какую энергетическую ценность имеет подготовительный, аэробный и анаэробных гликолизы, энергетического обмена в клетке?
Какую энергетическую ценность имеет подготовительный, аэробный и анаэробных гликолизы, энергетического обмена в клетке?
Какие процессы характерны для аэробного гликолиза?
Какие процессы характерны для аэробного гликолиза?
А - продукты окисляются до диоксида углерода и воды.
Б - продукты гликолиза превращаются в этиловый спирт в - образуется молочная кислота.

В каких условиях возник аэробный тип обмена веществ у прокариот?
В каких условиях возник аэробный тип обмена веществ у прокариот.

Куда девается глюкоза в аэробном процессе клетки?
Куда девается глюкоза в аэробном процессе клетки.
Полное расщепление одной молекулы глюкозы в мышцах в процессе аэробного дыхания дает около 25 молекул АТФ?
Полное расщепление одной молекулы глюкозы в мышцах в процессе аэробного дыхания дает около 25 молекул АТФ.
Сравните эту величину с количеством молекул АТФ, образующихся при гликолизе.
Какой процент энергии теряет в этом случае мышца?
При каких условиях процесс гликолиза все же идет в мышцах?
Насколько это выгодно организму?
Сколько моль пировиноградной кислоты образуется при распаде 1 моль глюкозы при аэробном гликолизе ?
Сколько моль пировиноградной кислоты образуется при распаде 1 моль глюкозы при аэробном гликолизе ?
Сколько моль АТФ будет синтезироваться в этом процессе ?
На странице вопроса Как и при каких условиях гликолиз может преобладать над аэробным процессом? из категории Биология вы найдете ответ для уровня учащихся 10 - 11 классов. Если полученный ответ не устраивает и нужно расшить круг поиска, используйте удобную поисковую систему сайта. Можно также ознакомиться с похожими вопросами и ответами других пользователей в этой же категории или создать новый вопрос. Возможно, вам будет полезной информация, оставленная пользователями в комментариях, где можно обсудить тему с помощью обратной связи.
ЦВЕТОК — ГЕНЕРАТИВНЫЙ ОРГАН. СОЦВЕТИЯ Цветок — орган семенного размножения. В цветке различают следующие части от периферии к центру : околоцветник, тычинки, пестик (или пестики). Околоцветник состоит из чашечки и венчика. Чашечка окружает венчик..
Приспособлением плода к распространению водой это увеличение его плавучести. Этого достигается за счет того, что образуются воздушные полости внутри плода. Например , кокос , он не тонет . Или кувшинка.

В процессе фотосинтеза энергия света расходуется также на расщепление молекулы воды - фотолиз. При этом образуются протоны (Н + ), электроны (е) и свободный кислород : 2Н2О (свет) = > 4H( + ) + 4e( - ) + O2 Электроны, образующиеся при фотолизе, восп..
Аденин комплиментарен тимину и равен ему Тимин - 28 Аденин - 28 Остальное гуанин и цитозин 100 - 28 - 28 = 44 44÷2 = 22 Цитозин - 22 Гуанин - 22.

Начнем с самой примитивной и закончим наиболее развитой нервными системами 1. Диффузная - кишечнополостные 2. Лестничная - характерна для плоских червей 3. Нервная цепочка - кольчатые черви. Членистоногие 4. Разбросанно - узловая - моллюски 5. ..

Это зависит от среды обитания (и от надобности каких - либо органов чуаств). Например : крот обитает в почве, имеет плохое зрение на поверхности(ему оно не нужно) ; какой - нибудь хищник, например рысь имеет очень хорошее зрение для охоты ночью и т.
Органы чувств — специализированная периферическая анатомо - физиологическая система, обеспечивающая, благодаря своим рецепторам, получение и первичный анализ информации из окружающего мира и от других органов самого организма, то есть из внешней сред..
Ощущение растягивания в зоне костей ( и неприятное ощущение в той области ).
С помощью циркуляторной системы и кислорода осуществляется транспорт веществ в организме.
Читайте также: