
Сообщение на тему самоопыление
Обновлено: 05.07.2024
Опыление — это перенос пыльцы из пыльников на рыльце пестика или пыльцевход семяпочки. Различают два основных типа опыления: перекрёстное и самоопыление.
Перекрестное опыление
При перекрёстном опылении пыльца из пыльников попадает на рыльце пестика другого цветка. В результате этого соединяются наследственно разнородные гаметы, поэтому потомство получается более жизнеспособным, чем при самоопылении. Перекрёстное опыление биологически более совершенно и широко распространено в мире растений.
Приспособления к опылению
Для обеспечения перекрёстного опыления у растений в процессе эволюции выработался ряд приспособлений: автостерильность или самобесплодность (у вишни, груши, яблони); неодновременное созревание тычинок и пестиков (бобовые, зонтичные, сложноцветные и др.); разная длина тычинок и пестиков (губоцветные); образование однополых цветков (береза, ива, дуб).
Гибридизация
Перекрёстное опыление может происходить не только между особями одного вида, но и между разными видами и разновидностями. В этом случае происходит гибридизация, которая приводит к образованию половых гибридов со смешанной наследственностью.
Способы перекрестного опыления
Перенос пыльцы может осуществляться насекомыми, птицами, ветром, водой.
Опыление насекомыми. У растений, опыляемых насекомыми, цветки с ярким околоцветником, нередко имеют сильный запах. Мелкие цветки собраны в соцветия. Привлекают насекомых также нектар и пыльца, служащие для них пищей.
Возможность самоопыления у цветковых растений с обоеполыми цветками отнюдь не исключена, несмотря на наличие у них разнообразных устройств, противодействующих ему. Для подавляющего большинства покрытосеменных, за исключением двудомных и строго самонесовместимых видов, следует допустить, что некоторая, иногда довольно большая, часть семян ежегодно образуется в результате самоопыления.
Наряду со случайной автогамией нередко встречается более или менее регулярная автогамия, возникающая под давлением условий, затрудняющих перекрестное опыление или препятствующих ему. Это вынужденное самоопыление. Оно представляет резервный способ опыления и должно рассматриваться как адаптация к неблагоприятным условиям среды.
Самоопыление у однолетних растений встречается чаще, чем у многолетних. Однолетние самоопыляющиеся виды имеются во многих семействах (крестоцветные, бобовые, злаки и др.). Примерами их могут служить крошечные мелкоцветковые растения резушка Таля(Arabidopsis thialiana), пастушья сумка, веснянка весенняя, клевер скученный, клевер шершавый, клевер подземный, герань нежная и др. Они растут на сухих и бедных почвах, быстро завершают жизненный цикл, самоопыление у них очень обычно и связано с особым: образом жизни. Ему благоприятствуют, видимо, два обстоятельства. Прежде всего популяции однолетних растений подвержены сильным колебаниям численности в разные годы. Самоопыление позволяет каждой такой популяции легче и быстрее восстановиться после понесенного ею ущерба, когда она уменьшается до малого числа особей или даже до единственного индивидуума. Кроме того, однолетние растения обычно связаны с узко ограниченными и специфическими местообитаниями. Благодаря самоопылению адаптация этих однолетних видов к таким условиям оказывается вполне устойчивой, что дает им известные преимущества в борьбе за жизнь. Здесь напрашивается аналогия с однолетними самоопыляющимися культурными растениями (культурные виды пшениц, ячменя, овса, гороха посевного и др.), гомозиготные популяции которых в определенных условиях культуры вполне устойчивы и продуктивны.
Самоопыление возникает на базе ксеногамного опыления в результате нарушения (дегенерации) важнейших его механизмов — самонесовместимости и дихогамии. Вследствие подавления генов самонесовместимости происходит сдвиг в сторону более или менее полной самосовместимости. Дихогамия становится как бы стертой вследствие удлинения начальных фаз (рыльцевой и тычиночной) в развитии цветка, благодаря чему эти фазы совмещаются или налегают друг на друга.
Автогамия происходит в обоеполых цветках в разные периоды цветения — в самом начале его, .иногда еще в бутонах (бутонная автогамия), на протяжении всего цветения или в самом конце его.
Бутонная автогамия встречается редко. Более обычной является автогамия в конце периода цветения, когда перекрестное опыление с помощью ветра или насекомых по каким-либо причинам, чаще из-за дождливой и холодной погоды или из-за отсутствия опылителей, не осуществилось. В этом случае страхующая функция автогамии выступает очень наглядно.
Автогамия осуществляется разными способами: при прямом соприкосновении рыльца пыльников (контактная автогамия), при высыпании пыльцы из пыльников и оседании ее на рыльце под действием собственной тяжести (гравитационная автогамия), при помощи дождя, (омброавтогамия), ветра (анемоавтогамня), мельчайших насекомых, обитающих в цветке (трипсавтогамия).
Контактная автогамия наиболее обычна. В начале цветения, когда еще не утрачены шансы на перекрестное опыление, пыльники и рыльца в цветке созревают в разное время или расположены таким образом, что непосредственный контакт между ними невозможен. Позднее в цветке происходят изменения во взаимном расположении пыльников и рыльца. Они связаны с ростовыми движениями и проявляются в удлинении или изгибе тычинок или столбиков, благодаря чему вскрывшиеся пыльники и ставшие восприимчивыми рыльца располагаются на одном уровне и в непосредственной близости.
Контактную автогамию можно наблюдать у характерных растений темнохвойного леса (седмичника европейского, цирцеи альпийской, майника двулистного). У седмичника цветки слабо прототипичны. Рыльце восприимчиво к моменту распускания цветка. В это время пыльники находятся примерно на одном уровне с рыльцем и рядом с ним, что делает возможным их соприкосновение. Липкое рыльце прочно удерживает пыльник. Пыльник при покрывании высыпает на него пыльцу. Подобная же контактная автогамия встречается у цирцеи альпийской и майника двулистного. У последнего возможна также гейтоногамия между соседними цветками соцветия. У рассмотренных растений темнохвойного леса автогамия имеет более или менее случайный характер.Никаких структурных приспособлений к автогамии и связанных с ней редукций в цветке нет. Цветки этих растений вполне сохранили энтомофильный облик, но опыление насекомыми у них утрачено, вследствие чего они перешли к автогамин.
Копытень европейский — обычное растение в еловых лесах южной тайги и в смешанных и широколиственных лесах. Цветет в мае и первой половине июня. Цветки одиночные, невзрачные, располагаются у поверхности почвы, скрыты листьями. Венчик колокольчатый, спайнолепестный, снаружи буроватый, изнутри грязпо-пурпуровый. Долгое время способ опыления у копытня оставался неясным. Сейчас единодушно признается наличие у него обязательной контактной автогамии.
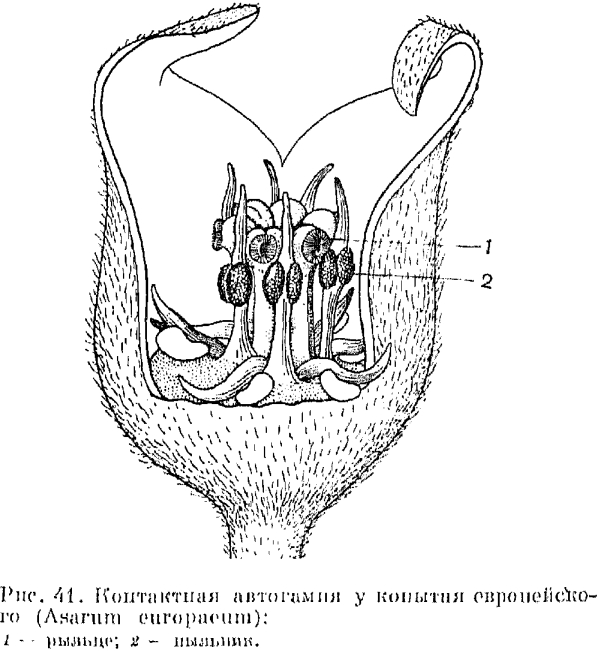
В развитии цветка копытня можно выделить три стадии —- начальную женскую, среднюю обоеполую и конечную мужскую, Для опыления имеет значение только средняя стадия. Цветки прототипичны: в бутоне и тотчас после распускания цветка рыльце восприимчиво, но пыльники 12 тычинок еще не вскрыты. В следующей, обоеполой стадии, при еще функционирующем рыльце, пыльники 6 длинных тычинок внутреннего круга вдвигаются между лопастями рыльца и, вскрываясь, оставляют на них пыльцу (рис. 41) на этой стадии легко можно обнаружить целые связки пыльцевых трубочек, возникших из пыльцевых зерен, находящихся еще во вскрытых пыльниках и врастающих в рыльце. Наконец, в последней, мужской стадии развития цветка рыльце уже утратило восприимчивость, а пыльники 6 наружных тычинок вскрываются, но, будучи более короткими, не достигают уровня рыльца. Поэтому эта конечная стадия, как и первая, не имеет сейчас значения для опыления, но могла иметь его в филогенетическом прошлом, когда копытень был еще энтомофильным растением. Облигатная (обязательная) автогамия копытня очень близка к клейстогамии. Иногда у него обнаруживаются закрытые цветки, в которых произошла автогамия.
С гравитационной автогамией лучше всего можно познакомиться на примере одноцветки (табл. 1.). Это очень красивое растение еловых лесов. Крупные белые венчики его как звездочки мерцают в сумраке леса. Они источают нежный аромат, но не посещаются насекомыми. Для осуществления автогамии у одноцветки необходима определенная ориентация цветоножки. Тогда, как это показано на рисунке 42, пыльца, высыпающаяся из пыльников, попадает на рыльце. То же самое, по-видимому, происходит в поникших цветках вересковых (брусинки, черники, голубики, подбела и грушанок круглолистной и средней, тогда как у грушанки малой имеется, видимо, контактная автогамия, поскольку пыльники у нее очень приближены к рыльцу). Ветер может содействовать гравитационной автогамии, поскольку при раскачивании и сотрясении растений происходит высыпание пыльцы через отверстия (поры), имеющиеся в пыльниках вересковых.
Своеобразный способ автогамии цветков при помощи дождя описан известным датским ботаником Олафом Хагерупом на Фарерских островах. Венчик цветка во время дождя наполняется до определенного уровня водой, причем пыльца плавает на ее поверхности. Пыльцевые зерна переносятся при помощи дождевых капель из пыльников на рыльце того же цветка. Насколько действен этот способ опыления, пока еще неизвестно, и по этому поводу высказываются сомнения.
Автогамии могут способствовать мельчайшио насекомые, обитающие в цветке (трипсы). это так называемая трипсоавтогамия. Она встречается у самых различных растений.
Клейстогамия представляет крайнюю форму автогамии. Она осуществляется в закрытых цветках, имеет экологический характер и вызывается неблагоприятными условиями внешней среды. Причины, вызывающие клейстогамию, разнообразны и у разных видов неодинаковы. Среди таких причин нужно назвать засуху, высокую атмосферную влажность, низкую или высокую температуру, условия освещения (глубокая тень, несоответствие длины фотопериода), затопление водой, скудное минеральное питание, неблагоприятные почвенные условия и т. д.
Клейстогамия выражена в разной степени. В соответствии с этим можно различать обязательную и факультативную клейстогамию. Под обязательной клейстогамией мы понимаем такие случаи, когда у вида постоянно образуются клейстогамные цветки, обнаруживающие черты более или менее далеко зашедшей редукции. Среди обязательных клейстогамов весьма обычны такие, у которых на одной и той же особи образуются нормальные хазмогамные и редуцированные клейстогамные цветки, причем именно последние являются плодущими. Развиваются эти разные формы цветков в разные сроки вегетационного периода, роже одновременно.
Клейстогамные цветки очень мелкие, никогда не раскрываются, напоминают бутоны. Чашечка уменьшена в размерах, лепестки рудиментарны или совсем отсутствуют, число тычинок часто уменьшено, пыльники мелкие, содержат мало пыльцы, которая не высыпается на рыльце, а прорастает в гнездах пыльника. При этом пыльцевые трубки прободают стенку пыльников и достигают рыльца. Последнее слабо развито. Эти цветки не выделяют нектар и не испускают аромат. В описанном случае развитие клейстогамии зашло далеко. Клейстогамия вызвала упрощение структуры цветка —- известную редукцию его органов и утрату ряда приспособлений к перекрестному опылению. Вместе с тем клейстогамия представляет важное приспособление к неблагоприятным условиям среды, обеспечивая постоянно высокую семенную продуктивность.
Хорошим примером такой постоянной, далеко зашедшей клейстогамии, сопровождающейся явлениями редукции в цветке, могут служить многие фиалки и кислица обыкновенная.
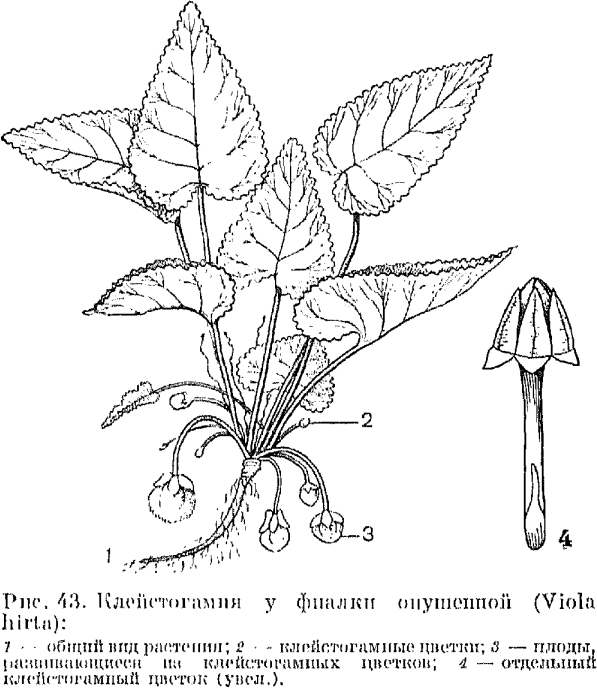
Фиалка удивительная — довольно обычное растение широколиственных и смешанных лесов. Она образует весной более или менее одновременно клейстогамные и хазмогамные открытоцветущие цветки. Последние в большинстве стерильны. У фиалки опушенной, произрастающей в широколиственных лесах, по сухим склонам,кустарниковым зарослям, ранней весной появляются на длинных цветоножках красивые цветки с крупными фиолетовыми венчиками. Они имеют нормально развитые тычинки и завязи, но, как правило, бесплодны. После их увядания на коротких цветоносах развиваются крохотные цветки; лепестки их прикрыты чашечкой. К моменту, когда начинает развиваться завязь, тычинки и лепестки в виде шапочки располагаются на рыльце. Из этих клейстогамных цветков образуются крупные многосемянные коробочки. Они сидят на удлинившихся к этому времени цветоносах, протягивающихся по поверхности земли (рис. 43). Клейстогамные цветки развиваются и у других фиалок (душистой, холмовой, собачьей).
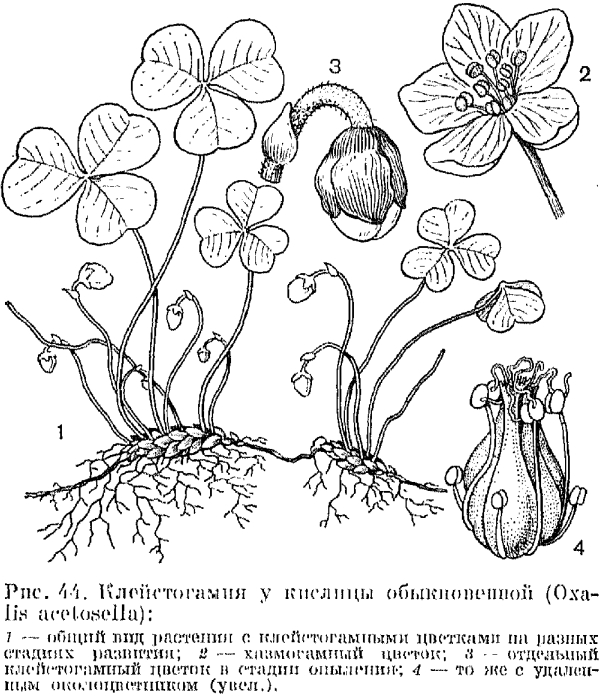
Кислица обыкновенная — характерное растение темнохвойной тайги. Она цветет в конце весны — начале лета (май — июнь). Крупные бело-розовые цветки ее многочисленны и являются украшением сумрачных еловых лесов. Из этих хазмогамных цветков образуется некоторое количество плодов-коробочек. Клейстогамные цветки кислицы мелкие, напоминают бутоны, не превышают 3 мм, сидят на коротких цветоножках, скрытых в моховом покрове (рис. 44). Появление их совпадает с началом созревания семян: в коробочках хазмогамных цветков (примерно спустя месяц после их цветения). В клейстогамных цветках кислицы лепестки сильно редуцированы и имеют вид крошечных чешуек, пыльники никогда не вскрываются, пыльца прорастает в пыльниках, пыльцевые трубки проникают через стенку пыльника и растут в сторону рыльца, нередко круто изгибаясь. По сравнению с хазмогамными цветками гинецей здесь отличается тем, что столбики у него короче и сосочки на рыльцах развиты слабее, а иногда столбиков совсем нет и рыльцевая поверхность располагается на вершине плодолистиков. Клейстогамия у кислицы обусловлена сильным затенениоми отчасти повышенным или избыточным увлажнением.
Особую группу составляют растения с подземными клейстогамными цветками. Лучше других известен из этой группы арахис подземный, называемый также китайскими орешками. Это культурное масличное растение родом из Южной Америки. У нас его возделывают на юге Украины, Северном Кавказе, в Закавказье и Средней Азии. У арахиса бобы образуются в почве. С самого начала они развиваются под землей из подземных клейстогамных цветков. Однако у арахиса имеются еще и надземные цветки с оранжево-желтыми венчиками, расположенные у самой поверхности почвы или несколько выше на стебле. После опыления этих цветков завязи их внедряются в почву на глубину 8 см вследствие удлинения ножки (гинофора), на которой сидит завязь.
Факультативная клейстогамия, будучи по своей природе также экологической, представляет как бы начальную фазу развития данного явления — она далеко не столь постоянна, проявляется лишь временами, при определенном сочетании условий среды и не сопровождается (это самое главное) признаками редукции в цветке. Факультативная клейстогамия довольно обычна. Примером ее могут служить ковыли, хлебные и другие злаки, отдельные виды пустынных маревых, некоторые водные и болотные растения — частуха подорожниковая, вахта трехлистная, виды рода росянка (Droseга), вербейник обыкновенный и др.
Превосходный пример факультативной клейстогамии представляют ковыли — характернейшие растения степей. У ковылей бывает открытое (хазмогамное) и закрытое (клейстогамное) цветение. Открытое цветение и перекрестное опыление при помощи ветра происходит у них ночью и на рассвете, но бывает далеко не всегда. Нередко ковыли полностью или частично цветут клейстогамно. Клейстогамия вызывается у них почвенной засухой (ксероклейстогамия) или пониженной температурой (термоклейстогамия). Клейстогамия у ковылей усиливается, и становится преобладающей в болое сухие сезоны и в более засушливых районах.
Экологический характер имеет клейстогамия у хлебных злаков. У овса она вызывается высокой или очень низкой температурой, а у ячменя — недостатком тепла и дождливой погодой на севере и засухой в степной золе. Что касается пшениц, то представления об условиях, способствующих или препятствующих открытому цветению их, очень противоречивы. Пo-видимому, теплая и влажная погода способствует открытому цветению, тогда как засуха и высокая температура — клейстогамному.
Итак, самоопыление у цветковых растений представляет явление вторичное, вызванное неблагоприятными для ксеногамного опыления условиями среды. Соотношение самоопыления и разных способов перекрестного опыления зависит от экологических условий. Эта зависимость отчетливо выступает в темнохвойной тайге, Арктике и пустынях, поскольку именно здесь крайние условия часто препятствуют осуществлению перекрестного опыления.
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров . 1974 .
Есть два виды опыления, с учетом источника пыльцы: самоопыление и перекрестное опыление. Кроме того, автоопыление можно подразделить на автогамию и геотогамию..
Опыление - это процесс, при котором пыльцевые зерна какой-либо пыльника - мужская часть цветка - переносятся в женскую часть цветка, известную как стигма..

Чтобы опыление было успешным, перенесенные пыльцевые зерна должны быть одного и того же вида..
Самоопыление - это вид опыления, при котором пыльца с пыльников цветка передается рыльцам того же цветка..
Перекрестное опыление включает перенос пыльцевых зерен с цветка одного растения на клеймо цветка другого растения. Это единственный тип опыления, который приводит к стигме различных генетических типов пыльцевых зерен во время опыления..
В зависимости от агента опыления перекрестное опыление можно классифицировать на абиотическое и биотическое опыление..
Типы опыления
1- Самоопыление
Это самый основной тип опыления, потому что он включает только один цветок. Этот тип опыления происходит, когда пыльцевые зерна пыльника падают прямо на рыльце того же цветка..
Хотя этот тип опыления является простым и быстрым, он приводит к сокращению генетического разнообразия, поскольку сперма и яйцеклетки одного и того же цветка имеют генетическую информацию.
Этот механизм самоопыления наблюдается у некоторых овощей, таких как арахис и соя. У большинства самоопыляющихся растений маленькие, незаметные цветки..
Эти цветы проливают пыльцу прямо на клеймо, еще до того, как распустится бутон..
Растения, которые следуют за процессами автоматического опыления, часто имеют одинаковое количество тычинок и ковров. Растения опыляются и могут давать потомство, которое самооплодотворяется.
Некоторые растения, которые проявляют этот тип опыления, включают персики, инжир, розы, помидоры, орхидеи и фиалки, среди других..
Самоопыление можно разделить на автогамию и геотогамию.
- автогамия
Это относится к слиянию двух гамет от одного человека. Автогамия преимущественно наблюдается в форме самоопыления.
Это происходит, когда сперма пыльцы из тычинки растения достигает ковров того же растения и оплодотворяет существующую яйцеклетку. При этом типе самоопыления сперма и яичники, собранные вместе, происходили из одного цветка..
- Geitogamia
У цветковых растений пыльца переносится с одного цветка на другой цветок на том же растении. В системах опылителей с животными это достигается, когда опылитель посещает несколько цветов одного растения.
Этот процесс также возможен у видов, которые опыляются воздухом, и может быть обычным источником семян, которые самооплодотворяются у самосовместимых видов.
Хотя функциональная геотогамия представляет собой перекрестное опыление с участием опыляющего агента, она генетически сходна с аутогамией, поскольку пыльцевые зерна происходят от одного растения..
Кукуруза - это растение, которое проявляет геотогамию.
Преимущества самоопыления
- Растения, которые опыляют себя, тратят меньше энергии на производство привлекательных опылителей..
- Они могут расти в областях, где организмы, которые могут помочь опылению, такие как насекомые и другие животные, отсутствуют или отсутствуют. Это включает в себя арктические районы и районы с очень высокими отметками.
- Этот процесс позволяет распространять растения за пределы диапазона доступных опылителей или производить потомство в регионах, где наблюдается сокращение популяции опылителей..
- Вероятность неудачного опыления меньше, и поэтому они помогают поддерживать чистоту своего вида..
Недостатки самоопыления
- Там нет возможности для производства новых видов.
- Потомки проявляют меньше энергии.
- Нежелательные характеристики не могут быть устранены.
- Способность противостоять болезням снижается.
- Это не помогает эволюции.
- Новые функции не введены.
2- Перекрестное опыление
Это происходит, когда пыльцевые зерна переносятся на цветок другого растения. Растения, которые проходят через этот процесс, часто имеют тычинки длиннее, чем их ковры.
Эти растения используют механизмы, обеспечивающие распространение пыльцевых зерен на другие цветы растения..
Процесс перекрестного опыления требует помощи биотических или абиотических агентов, таких как воздух, вода, насекомые, птицы и другие животные, которые действуют как опылители.
- Абиотическое опыление
Опыление проводится без вмешательства других организмов. Наиболее распространенная форма - опыление ветром; Опыление водой существует в водных растениях.
- Биотическое опыление
Это опыление требует опылителей для передачи пыльцевых зерен от пыльника к восприимчивой части или рыльца ковров или пестиков..
Существует много форм биотического опыления. Опыление насекомыми, опыление птицами или летучими мышами, а также человеком - наиболее распространенные варианты.
Растения, которые используют этот тип опыления, обычно имеют характеристики запаха, цвета и формы для привлечения опылителей..
Опыление насекомыми происходит у растений, у которых есть окрашенные лепестки и сильный запах, чтобы привлечь насекомых; У растений, которые опыляются воздушными позвоночными, обычно есть белые лепестки и эффектные запахи. Цветы, опыленные птицами, имеют трубчатые венчики ярких цветов.

Многие дачники при выборе посадочного материала в описании к тому или другому сорту растений встречают упоминание о различных способах опыления или самоопылении. Это понятия, которые мы все изучали еще в школе на уроках ботаники. Но не многие уже помнят, что они означают. Давайте освежим нашу память и вспомним типы опыления у растений и их биологическое значение. А заодно узнаем, почему не дают плодов некоторые наши растения, высаженные на даче или подоконнике.
Генеративный орган высших растений
Цветок – это видоизмененный побег, где формируются споры и гаметы. Высшие растения (покрытосеменные) имеют сложно устроенные цветки со множеством приспособлений к различным типам опыления. Разнообразный по деталям цветок совмещает в себе процессы как полового, так и бесполого размножения. Главными компонентами цветка являются его репродуктивные части – мужской андроцей (тычинки) и женский гинецей (пестик с завязью, столбик и рыльца). Цветки могут быть обоеполые (есть и пестик, и тычинки) и однополые (есть либо пестик, либо тычинки). Другие составные части цветка очень разнообразны и выполняют специфические функции.
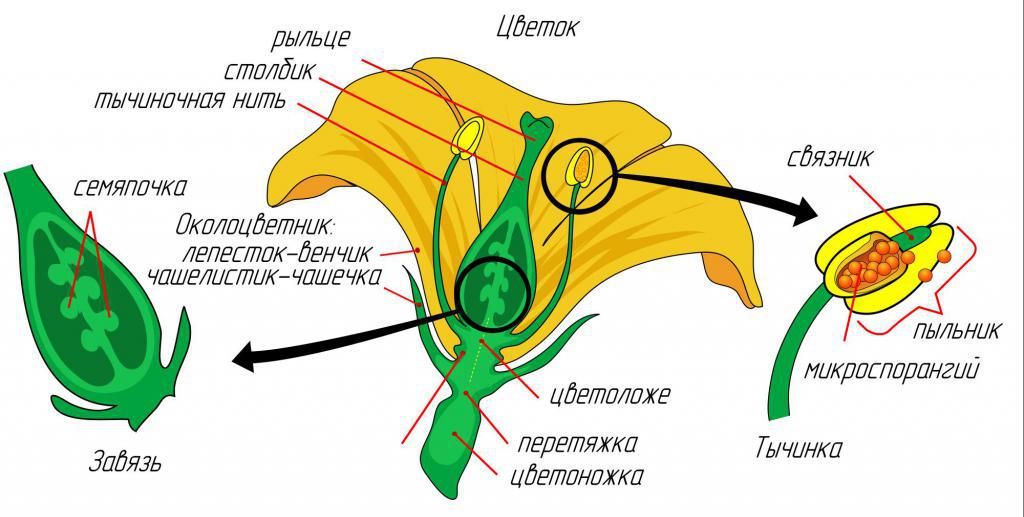
Встреча тычинок и пестика
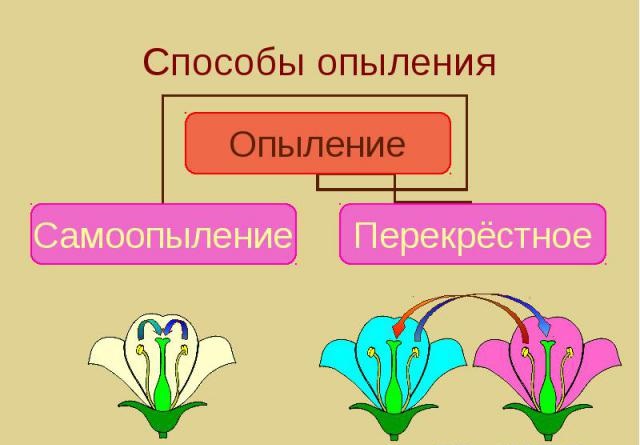
Опыление – это процесс переноса пыльцы с тычинок на рыльце пестика. Без этого невозможно размножение растений, формирование плодов и семян. В процессе эволюционного развития растения выработали несколько способов осуществления этого переноса при использовании биотических и абиотических факторов природы. В экологии выделяют два типа опыления:
- Перенос пыльцы с одного цветка на пестик другого. Этот процесс называется перекрестным опылением, или ксеногамией. Он осуществляется посредством биотических (насекомые, птицы, летучие мыши) и абиотических (ветер, вода) факторов.
- Автогамия (самоопыление). Это перенос пыльцы на рыльце с одного цветка. Автогамия не так часто встречается у дикорастущих форм.
Это типы опыления, которые могут чередоваться у некоторых растений.
Условия самоопыления
Но обязательным условием осуществления самоопыления является обоеполость цветка. Случайное самоопыление цветков – нередкое событие. Но оно может происходить только при физиологической совместимости пыльцевого зерна и пестика. У многих растений пыльца не прорастает в пыльцевую трубку, что является ограничителем для перекрестноопыляемых растений. Факторов, которые способствуют случайной автогамии, довольно много. Регулярное самоопыление растений (пример – горох, фасоль) может иметь гравитационный механизм. В этом случае пыльца попадает на рыльце под действием силы тяжести. В других случаях самоопыление происходит как контактная автогамия – тычинка соприкасается с рыльцем пестика. Переносчиками пыльцы в середине цветка могут выступать капли росы и мелкие насекомые (трипсы), которые живут в цветке. У некоторых растений процесс происходит в бутоне и полностью исключает возможность перекрестного опыления.
Факультативное самоопыление
Особенностью такого типа автогамии является наличие нестабильных условий, которые не благоприятствуют перекрестному опылению. Такой тип самоопыления встречается у злаковых, росянки, ковыля. У этих растений в засуху или при пониженных температурах формируются цветки однополые, а в теплую и влажную погоду – обоеполые. Перекрестное опыление этих растений осуществляется при помощи ветра, и в условиях затруднения осуществления такого опыления биологически целесообразно прибегнуть к самоопылению.

Эволюционное значение
Самоопыление в эволюционном плане имеет отрицательное значение. В соответствии с современными представлениями, для эволюции необходимо свободное скрещивание, которое обеспечивается перекрестным опылением. Именно оно повышает разнообразие аллелей (степени проявления гена) в популяциях. А самоопыление, наоборот, ведет к гомозиготности (однотипности) аллелей. Но при определенных обстоятельствах самоопыление может вести к изоляции новых форм, обособлению и фиксации в популяции аллелей, дающих благоприятные признаки растению. Именно в этом и заключается положительное эволюционное значение чередования автогамии и ксеногамии.

Самоопыляющиеся растения
У таких растений чаще перенос пыльцы осуществляется еще в нераскрытом бутоне (например, у фасоли и гороха) или в период еще не раскрытой листовой трубки (ячмень). Самоопылителями из сельскохозяйственных культур считают горох, фасоль, ячмень, пшеницу, овес, помидоры, баклажаны и многие другие. Почему считают? Потому что самоопыление не может быть абсолютным, всегда есть вероятность заноса пыльцы с других растений. Даже закрытые бутоны иногда прогрызают насекомые и заносят пыльцу других растений! Какие же признаки отличают самоопылителей? Это однозначно растения с обоеполыми цветками, большими перистыми рыльцами и большим количеством пыльцы. Кроме того, у их цветов нет ярких лепестков, нектарников и приятного запаха.
Самоопыление у фиалок
В природе фиалки опыляются и перекрестно, и автогамно. Наши комнатные фиалки – продукт кропотливой работы селекционеров. У них такое строение тычинок и пестика, что перекрестное опыление без участия человека практически невозможно. Опыление происходит еще в нераскрытом бутоне, и только терпеливый любитель с помощью специальных методик может осуществить опыление разных по цветовой гамме фиалок для выведения новых сортов. Спасибо энтузиастам за разнообразие этих цветов, украшающих наши подоконники!

Партенокарпические огурцы
Современная селекция предлагает множество сортов огурцов, как самоопыляемых (партенокарпических), так и опыляемых насекомыми. Эти растения выведены специально для раннего выращивания в теплицах, где нет естественных опылителей. Покупая семена, необходимо останавливаться на прочтении качеств сорта, так как свои преимущества и недостатки имеются и у самоопыляемых, и у перекрестноопыляемых сортов.

Опыление у злаковых
Овес, рожь, пшеница, просо, ячмень – представители сельскохозяйственных злаковых. Цветки имеют 2 цветковые чешуйки, 2 пленки, три тычинки и один пестик. Самоопыление у них происходит в нераскрытых цветках. Когда цветок раскрылся, перекрестное опыление практически исключено.

Самоопыление у плодовых деревьев
Хотя большинство плодовых сортов имеют цветки, в которых есть и пестики, и тычинки, самооплодотворение у большинства исключено. Причина – разделенное по времени созревание тычинок и пестика. Именно поэтому повысить урожай, например черешни, можно, посадив рядом несколько деревьев. Но у искусственно выведенных сортов самоопыление приветствуется. Пример – нектарины. Но не надейтесь вырастить из косточки урожайное растение. У таких гибридных форм последующие поколения испытывают гибридную депрессию – снижение жизнеспособности и урожайности.

Селекция и самоопыление
Это явление широко используется в селекции растений. Нам известно, что самооплодотворение и скрещивание близкородственных организмов ведет к переходу генов в гомозиготное состояние и приводит к снижению жизнестойкости и урожайности, а впоследствии и к вырождению. Непрерывный процесс мутаций, которые накапливаются, большинство из которых рецессивны и неблагоприятны, – причина этого угнетения. У растений с перекрестным опылением эти мутации находятся в гетерозиготном состоянии и никак не проявляются. При самоопылении вероятность их перехода в гомозиготу во много раз возрастает, но они не сохраняются в популяции из-за естественного устранения. Самоопыление в селекции используется как инструмент создания чистых (гомозиготных) линий с закрепленными признаками. Несмотря на снижение продуктивности, после гибридизации часто появляется явление гетерозиса – силы гибридов от сортов с самоопылением. Это явление называется межлинейной гибридизацией, и в магазинах мы можем увидеть именно такие гибридные семена (они помечены символом F1). В первом поколении гибриды превосходят чистые линии по урожайности, но в дальнейших поколениях эффект силы гибридов исчезает.
Читайте также: