
Закономерности биологической эволюции реферат
Обновлено: 02.07.2024
Палеонтологические. Ранее существовавшие организмы оставляют после себя различные формы ископаемых остатков: окаменелости, отпечатки, скелеты, следы деятельности. По этим остаткам можно проследить изменение групп организмов во времени. Реконструированы филогенетические ряды лошадиных, хоботных, некоторых моллюсков. Обнаружено множество переходных форм между современными группами организмов… Читать ещё >
Основные закономерности макроэволюции ( реферат , курсовая , диплом , контрольная )
Государственное Образовательное Учреждение Высшего Профессионального Образования Мордовский Государственный Университет Имени Н. П. Огарева Биологический факультет Кафедра генетики Реферат На тему: Основные закономерности макроэволюции Саранск
1. Доказательства эволюции органического мира
По современным представлениям, биологическая эволюция — это необратимое и, в известной мере, направленное историческое развитие живой природы, сопровождающееся изменением генетического состава популяций, формированием адаптаций, образованием и вымиранием видов, преобразованиями биогеоценозов и биосферы в целом.
Существуют многочисленные доказательства эволюции органического мира Земли, которые одновременно являются и методами изучения эволюции. К классическим доказательствам эволюции относятся палеонтологические, сравнительно-анатомические и сравнительно-эмбриологические.
1. Палеонтологические. Ранее существовавшие организмы оставляют после себя различные формы ископаемых остатков: окаменелости, отпечатки, скелеты, следы деятельности. По этим остаткам можно проследить изменение групп организмов во времени. Реконструированы филогенетические ряды лошадиных, хоботных, некоторых моллюсков. Обнаружено множество переходных форм между современными группами организмов. Однако из-за неполноты палеонтологической летописи не всегда удается реконструировать ход эволюции.
Историю Земли палеонтологи делят на длительные промежутки — эры, а эры — на периоды. Название эр и периодов, их продолжительность и характерный для них животный и растительный мир приведены в геохронологической таблице. В ней продолжительность эр и периодов указана в соответствии с геохронологической шкалой, принятой Международным геологическим конгрессом. Таблица отражает расположение земных пластов: наиболее древние — внизу, более поздние — вверху. Продолжительность эр и периодов определяют по количеству продуктов распада радиоактивных элементов в пробе породы. Установлено, что за каждые 100 млн. лет из 1 кг урана независимо от внешних условий остается 985 г, а образуется 13 г свинца и 2 г гелия. Определив количество свинца и гелия в пробе данной породы, можно подсчитать геологический возраст породы.
2. Сравнительно-морфологические. Системы органов современных организмов образуют ряд последовательных изменений. Например, на современных организмах можно проследить судьбу отдельных костей мозгового и висцерального черепа. Так, позвоночные животные имеют двустороннюю симметрию, общий план строения скелета, черепа, передних и задних конечностей, головного мозга и всех основных систем (нервной, пищеварительной, кровеносной и др.). Единство происхождения подтверждает строение гомологичных органов, рудиментов, атавизмов и переходных форм.
К сравнительно-морфологическим доказательствам близки сравнительно-биохимические. Например, на современных организмах можно проследить изменение структуры гемоглобина. Однако в этих рядах имеются и пробелы, поскольку далеко не все переходные формы дожили до нашего времени.
3. Сравнительно-эмбриологические. В ходе эмбрионального развития у зародышей часто наблюдаются черты сходства с зародышами предковых форм. Например, у всех позвоночных на ранних стадиях развития появляются внутренние жабры (или их зачатки — жаберные карманы).
В ходе эволюции наблюдаются эволюционные преобразования процессов онтогенеза, связанные с адаптациями взрослых (половозрелых) организмов. В ходе таких преобразований могут появляться новые органы, но могут и утрачиваться старые органы (полностью или превращаться в рудименты). При этом могут изменяться: начальная масса зачатка органа, место и время закладки органа. Эти преобразования могут происходить на разных стадиях онтогенеза: на самых ранних (закладка хорды, нервной трубки), средних (закладка чешуи у рыб, перьев у птиц, видоизменение побегов растений) и поздних (редукция хвоста у головастиков, формирование четырехкамерного сердца у птиц и млекопитающих, изменение формы листьев). При изменениях органов на поздних стадиях онтогенеза и может действовать филогенетический закон.
В настоящее время для изучения эволюционного развития той, или иной группы организмов используется целый комплекс методов: биогеографические, экологические, генетические, молекулярно-биологические, иммунологические, биохимические, а также методы палеоэкологии, сравнительной физиологии и этологии; широко используются методы компьютерного моделирования.
2. Макроэволюция. Связь макроэволюции с микроэволюцией
Микроэволюцией называется совокупность эволюционных процессов внутри видов. Сущность микроэволюционных преобразований составляет изменение генетической структуры популяций. В результате действия элементарных эволюционных факторов появляются новые аллели, а в результате действия отбора формируются новые адаптации. При этом происходит замещение одного аллеля другим аллелем, одного изотипа белка (фермента) другим изотипом.
Популяции являются открытыми генетическими системами. Поэтому на микроэволюционном уровне происходит латеральный перенос генов — обмен генетической информацией между популяциями. Это означает, что адаптивный признак, возникший в одной популяции, может перейти в другую популяцию. Следовательно, микроэволюцию можно рассматривать как эволюцию открытых генетических систем, способных обмениваться генетическим материалом.
Таким образом, учение о макроэволюции включает, с одной стороны, учение о родственных отношениях таксонов, а с другой стороны, учение об эволюционных (филогенетических) преобразованиях признаков этих таксонов.
Связь между микроэволюцией и макроэволюцией отражена в законе гомологических рядов. Н. И. Вавилов создал учение о виде как системе. В этой теории вида внутривидовая изменчивость полностью отделена от таксономических различий (впервые такую попытку предпринял Дж. Рэй).
1 чередование поколений у самых разнообразных организмов;
2 появление раковины моллюсков;
3 появление шерстного покрова млекопитающих и перьев у птиц;
4 появление сегментации у членистоногих и позвоночных;
5 преобразования дуг аорты у позвоночных (вместе с мышцами, нервами и жаберными щелями);
6 появление зубов позвоночных;
7 появление сложных глаз у членистоногих и позвоночных.
Появление этих признаков может быть обусловлено макромутациями в генах, отвечающих не за структуру ферментов, а за регуляцию развития. Тогда макроэволюция представляет собой самостоятельное явление, не связанное с микроэволюцией. Такой подход устраивает противников дарвинизма, которые признают естественнонаучную основу микроэволюции, но отрицают естественнонаучную основу макроэволюции.
3. Общие закономерности эволюции
Макроэволюция представляет собой обобщенную картину эволюционных преобразований. Только на уровне макроэволюции обнаруживаются общие тенденции, направления и закономерности эволюции органического мира.
В течение второй половины XIX — первой половины XX века на основании многочисленных исследований закономерностей эволюционного процесса были сформулированы основные правила (принципы) эволюции. (Эти правила носят ограниченный характер, не имеют универсального значения для всех групп организмов и не могут считаться законами.)
1. Правило необратимости эволюции, или принцип Долло (Луи Долло, бельгийский палеонтолог, 1893): исчезнувший признак не может вновь появиться в прежнем виде. Например, вторично-водные моллюски и водные млекопитающие не восстановили жаберного дыхания.
2. Правило происхождения от неспециализированных предков, или принцип Копа (Эдуард Коп, американский палеонтолог-зоолог, 1904): новая группа организмов возникает от неспециализированных предковых форм. Например, неспециализированные Насекомоядные (типа современных тенреков) дали начало всем современным плацентарным млекопитающим.
3. Правило прогрессирующей специализации, или принцип Депере (Ш. Депере, палеонтолог, 1876): группа, вступившая на путь специализации, в дальнейшем развитии будет идти по пути все более глубокой специализации. Современные специализированные млекопитающие (Рукокрылые, Ластоногие, Китообразные), скорее всего, будут эволюционировать поп пути дальнейшей специализации.
4. Правило адаптивной радиации, или принцип Ковалевского-Осборна (В.О. Ковалевский, Генри Осборн, американский палеонтолог): группа, у которой появляется безусловно прогрессивный признак или совокупность таких признаков, дает начало множеству новых групп, формирующих множество новых экологических ниш и даже выходящих в иные среды обитания. Например, примитивные плацентарные млекопитающие дали начало всем современным эволюционно-экологическим группам млекопитающих.
5. Правило интеграции биологических систем, или принцип Шмальгаузена (И.И. Шмальгаузен): новые, эволюционно молодые группы организмов вбирают в себя все эволюционные достижения предковых групп. Например, млекопитающие использовали все эволюционные достижения предковых форм: опорно-двигательный аппарат, челюсти, парные конечности, основные отделы центральной нервной системы, зародышевые оболочки, совершенные органы выделения (тазовые почки), разнообразные производные эпидермиса и т. д.
6. Правило смены фаз, или принцип Северцова-Шмальгаузена (А.Н. Северцов, И.И. Шмальгаузен): различные механизмы эволюции закономерно сменяют друг друга. Например, алломорфозы рано или поздно становятся ароморфозами, а на основе ароморфозов возникают новые алломорфозы.
В дополнение к правилу смены фаз Дж. Симпсон ввел правило чередования темпов эволюции; по скорости эволюционных преобразований он различал три типа эволюции: брадителлическую (медленные темпы), горотеллическую (средние темпы) и тахителлическую (быстрые темпы).
Беклемишев В. Н. Основы сравнительной анатомии беспозвоночных 3-е изд. М., 1964.
Воробьева Э. И. Морфология и особенности эволюции кистеперых рыб. — Труды Палеонтологического ин-та АН СССР, т. 163. М., 1977.
Голенкин М. И. Победители в борьбе за существование. 3-е изд. М., 1959.
Давиташвили Л. Ш. История эволюционной палеонтологии от Дарвина до наших дней. М., 1948.
Джоплин Д. А., Ричардс Д. Р. , Джоплин К. А. Определение возраста пород. М., 1976.
Целью работы является изучение закономерностей протекания эволюции, а также микроэволюции и образования новых видов.
Для достижения этой цели были поставлены следующие задачи:
• изучить закономерности протекания эволюции;
• изучить понятие микроэволюции и образования новых видов.
Оглавление
Введение…………………………………………………………………………. 3
1. Закономерности протекания эволюции……………………………………….4
1.1. Доказательства эволюции органического мира………………………. 4
1.2. Общие закономерности эволюции………………………………………7
2. Микроэволюция и видообразование………………………………………….9
2.1. Понятие о микроэволюции и популяции……………………………….9
2.2. Способы видообразования……………………………………………. 12
Заключение……………………………………………………………………….17
Список использованной литературы…………………………………………. 20
Файлы: 1 файл
КСЕ.docx
Министерство образования и науки РФ
Негосударственное образовательное учреждение
высшего профессионального образования
Дисциплина: Концепция современного естествознания
Преподаватель: Веселовская Е.Д.
Выполнил: студент гр. 12 БЮЗ 5 …
1. Закономерности протекания эволюции……………………………………….4
1.1. Доказательства эволюции органического мира………………………. 4
1.2. Общие закономерности эволюции………………………………………7
2. Микроэволюция и видообразование……………………………………… ….9
2.1. Понятие о микроэволюции и популяции……………………………….9
2.2. Способы видообразования……………………………………… ……. 12
Список использованной литературы…………………………………………. 20
Даже в XVIII веке этот взгляд разделял великий шведский натуралист, основатель современной систематики Карл Линней. Другая точка зрения - трансформизм: мир изменяется, исторически развивается. Применительно к биологии это означало историческое изменение организмов, постепенное усложнение их организации. В XVIII веке такие мысли выражали Бюффон во Франции, Эразм Дарвин (дед Чарльза Дарвина) в Англии, Гёте в Германии, Ломоносов в России. Но первые эволюционные догадки были умозрительные, они не имели достаточной фактической основы. Только в XIX веке сформировалось научное эволюционное учение.
Целью работы является изучение закономерностей протекания эволюции, а также микроэволюции и образования новых видов.
Для достижения этой цели были поставлены следующие задачи:
- изучить закономерности протекания эволюции;
- изучить понятие микроэволюции и образования новых видов.
1.Закономерности протекания эволюции
1.1. Доказательства эволюции органического мира
По современным представлениям, биологическая эволюция - это необратимое и, в известной мере, направленное историческое развитие живой природы, сопровождающееся изменением генетического состава популяций, формированием адаптаций, образованием и вымиранием видов, преобразованиями биогеоценозов и биосферы в целом.
Существуют многочисленные доказательства эволюции органического мира Земли, которые одновременно являются и методами изучения эволюции. К классическим доказательствам эволюции относятся палеонтологические, сравнительно-анатомические и сравнительно- эмбриологические.
1. Палеонтологические. Ранее существовавшие организмы оставляют после себя различные формы ископаемых остатков: окаменелости, отпечатки, скелеты, следы деятельности. По этим остаткам можно проследить изменение групп организмов во времени. Реконструированы филогенетические ряды лошадиных, хоботных, некоторых моллюсков. Обнаружено множество переходных форм между современными группами организмов. Однако из-за неполноты палеонтологической летописи не всегда удается реконструировать ход эволюции.
Историю Земли палеонтологи делят на длительные промежутки - эры, а эры - на периоды. Название эр и периодов, их продолжительность и характерный для них животный и растительный мир приведены в геохронологической таблице. В ней продолжительность эр и периодов указана в соответствии с геохронологической шкалой, принятой Международным геологическим конгрессом. Таблица отражает расположение земных пластов: наиболее древние - внизу, более поздние - вверху. Продолжительность эр и периодов определяют по количеству продуктов распада радиоактивных элементов в пробе породы. Установлено, что за каждые 100 млн. лет из 1 кг урана независимо от внешних условий остается 985 г, а образуется 13 г свинца и 2 г гелия. Определив количество свинца и гелия в пробе данной породы, можно подсчитать геологический возраст породы.
2. Сравнительно-морфологические. Системы органов современных организмов образуют ряд последовательных изменений. Например, на современных организмах можно проследить судьбу отдельных костей мозгового и висцерального черепа. Так, позвоночные животные имеют двустороннюю симметрию, общий план строения скелета, черепа, передних и задних конечностей, головного мозга и всех основных систем (нервной, пищеварительной, кровеносной и др.). Единство происхождения подтверждает строение гомологичных органов, рудиментов, атавизмов и переходных форм.
К сравнительно-морфологическим доказательствам близки сравнительно-биохимические. Например, на современных организмах можно проследить изменение структуры гемоглобина. Однако в этих рядах имеются и пробелы, поскольку далеко не все переходные формы дожили до нашего времени.
3. Сравнительно-эмбриологические. В ходе эмбрионального развития у зародышей часто наблюдаются черты сходства с зародышами предковых форм. Например, у всех позвоночных на ранних стадиях развития появляются внутренние жабры (или их зачатки - жаберные карманы).
В ходе эволюции наблюдаются эволюционные преобразования процессов онтогенеза, связанные с адаптациями взрослых (половозрелых) организмов. В ходе таких преобразований могут появляться новые органы, но могут и утрачиваться старые органы (полностью или превращаться в рудименты). При этом могут изменяться: начальная масса зачатка органа, место и время закладки органа. Эти преобразования могут происходить на разных стадиях онтогенеза: на самых ранних (закладка хорды, нервной трубки), средних (закладка чешуи у рыб, перьев у птиц, видоизменение побегов растений) и поздних (редукция хвоста у головастиков, формирование четырехкамерного сердца у птиц и млекопитающих, изменение формы листьев). При изменениях органов на поздних стадиях онтогенеза и может действовать филогенетический закон.
В настоящее время для изучения эволюционного развития той, или иной группы организмов используется целый комплекс методов: биогеографические, экологические, генетические, молекулярно-биологические, иммунологические, биохимические, а также методы палеоэкологии, сравнительной физиологии и этологии; широко используются методы компьютерного моделирования.
1.2. Общие закономерности эволюции
Макроэволюция представляет собой обобщенную картину эволюционных преобразований. Только на уровне макроэволюции обнаруживаются общие тенденции, направления и закономерности эволюции органического мира.
В течение второй половины XIX - первой половины XX века на основании многочисленных исследований закономерностей эволюционного процесса были сформулированы основные правила (принципы) эволюции. (Эти правила носят ограниченный характер, не имеют универсального значения для всех групп организмов и не могут считаться законами.)
1. Правило необратимости эволюции, или принцип Долло (Луи Долло, бельгийский палеонтолог, 1893): исчезнувший признак не может вновь появиться в прежнем виде. Например, вторично-водные моллюски и водные млекопитающие не восстановили жаберного дыхания.
2. Правило происхождения от неспециализированных предков, или принцип Копа (Эдуард Коп, американский палеонтолог- зоолог, 1904): новая группа организмов возникает от неспециализированных предковых форм. Например, неспециализированные Насекомоядные (типа современных тенреков) дали начало всем современным плацентарным млекопитающим.
3. Правило прогрессирующей специализации, или принцип Депере (Ш. Депере, палеонтолог, 1876): группа, вступившая на путь специализации, в дальнейшем развитии будет идти по пути все более глубокой специализации. Современные специализированные млекопитающие (Рукокрылые, Ластоногие, Китообразные), скорее всего, будут эволюционировать поп пути дальнейшей специализации.
4. Правило адаптивной радиации, или принцип Ковалевского- Осборна (В.О. Ковалевский, Генри Осборн, американский палеонтолог): группа, у которой появляется, безусловно, прогрессивный признак или совокупность таких признаков, дает начало множеству новых групп, формирующих множество новых экологических ниш и даже выходящих в иные среды обитания. Например, примитивные плацентарные млекопитающие дали начало всем современным эволюционно-экологическим группам млекопитающих.
5. Правило интеграции биологических систем, или принцип Шмальгаузена (И.И. Шмальгаузен): новые, эволюционно молодые группы организмов вбирают в себя все эволюционные достижения предковых групп. Например, млекопитающие использовали все эволюционные достижения предковых форм: опорно-двигательный аппарат, челюсти, парные конечности, основные отделы центральной нервной системы, зародышевые оболочки, совершенные органы выделения (тазовые почки), разнообразные производные эпидермиса и т.д.
6. Правило смены фаз, или принцип Северцова-Шмальгаузена (А.Н. Северцов, И.И. Шмальгаузен): различные механизмы эволюции закономерно сменяют друг друга. Например, алломорфозы рано или поздно становятся ароморфозами, а на основе ароморфозов возникают новые алломорфозы.
В дополнение к правилу смены фаз Дж. Симпсон ввел правило чередования темпов эволюции; по скорости эволюционных преобразований он различал три типа эволюции: брадителлическую (медленные темпы), горотеллическую (средние темпы) и тахителлическую (быстрые темпы).
2. Микроэволюция и видообразование
2.1. Понятие о микроэволюции и популяции
Возникновение новых видов Дарвин представлял как длительный процесс накопления полезных изменений, увеличивающихся из поколения в поколение. Мелкие индивидуальные изменения учений принимал за первые шаги видообразования. Их накопление через много поколений приводит к образованию разновидностей, которые он рассматривал как ступени на путь образования нового вида. Переход от одной ступени к другой происходит в результате накапливающего действия естественного отбора. Разновидность, по мнению Дарвина, - это зарождающийся вид, а вид – резко выраженная разновидность.
В процессе эволюции из одного родоначального вида может возникнуть несколько новых. Такой процесс Дарвин называл дивергенцией или расхождением признаков. Под этим термином он понимал возникновение разнообразных существ в потомстве от одного предка. Из измененных форм выживают и дают потомство только наиболее уклонившиеся разновидности, каждая из которых вновь дает веер изменившихся форм, и опять выживают наиболее уклонившиеся и лучше приспособленные. Так, шаг за шагом возникают все большие различия между крайними формами, перерастающими, наконец, в различия между видами, семействами и так далее. В природе не всегда сохранялись лишь наиболее расходящиеся, крайние формы, средние так же могли выжить и дать потомство. Из крайних форм иногда развиваются одна, но может развиваться и более.
Если условия среды не изменяются или мало изменяются в течение длительного времени, то вид остается почти неизмененным по сравнению с родоначальным.
С тридцатых годов прошлого столетия внимание ученых привлекает популяция как форма существования вида. Новые исследования проливают свет на самые начальные этапы эволюционного процесса, которые протекают внутри вида и приводят к образованию новых внутривидовых группировок – популяций и подвидов. Этот процесс называют микроэволюцией. Она доступна непосредственному наблюдению и изучению, так как происходит в исторически короткое время.
При построении филогенетических рядов биологи-эволюционисты, помимо палеонтологических данных, широко используют сравнительный метод, с помощью которого они устанавливают сходство в строении организмов, их биохимических реакциях, особенностях размножения или иных свойствах, по которым можно судить о путях развития группы от общего предка.
Содержимое работы - 1 файл
Документ Microsoft Office Word.docx
При построении филогенетических рядов биологи-эволюционисты, помимо палеонтологических данных, широко используют сравнительный метод, с помощью которого они устанавливают сходство в строении организмов, их биохимических реакциях, особенностях размножения или иных свойствах, по которым можно судить о путях развития группы от общего предка.
Изучая процесс эволюции, ученые выделяют следующие характерные типы эволюционных изменений: параллелизм, конвергенция и дивергенция. Иногда один филогенетический ряд может содержать в себе примеры изменений различного типа.
Параллелизм. Хороший пример параллельных изменений дает эволюция дикобраза. Два разных вида этого колючего млекопитающего эволюционируют независимо друг от друга в Африке и в Южной Америке. Более 70 млн лет назад, когда они обитали вместе, их общий предок был похож на большую, покрытую шерстью крысу. Когда два континента разошлись, популяция разделилась на две части, каждая из которых развивалась независимо от другой. Однако, поскольку условия существования обеих новых популяций были сходны, развитие дикобразов шло параллельными путями. Несмотря на то что они самостоятельно жили более 70 млн лет, и американский и африканский дикобразы очень близки по строению и образу жизни и занимают сходные ниши в сообществах.
Конвергенция. При конвергентной (от лат. convergo — приближаюсь, схожусь) эволюции два или более вида, не связанные близким родством, становятся все более и более похожими друг на друга. Такой тип эволюционных изменений является результатом приспособлений к сходным условиям внешней среды. Крупные водные хищники, показанные на рисунке 81, возникли в четырех совершенно разных группах: среди рыб, пресмыкающихся, птиц и млекопитающих. Их внешнее сходство возникло в процессе эволюционного развития под влиянием образа жизни и факторов внешней среды при совершенно разных исходных положениях организмов. Это сходство скрывает глубокие различия внутреннего строения и обмена веществ, которые свидетельствуют о столь же глубоких различиях эволюционной истории изображенных животных. Пути эволюционных преобразований их внешнего строения можно изобразить в виде сходящихся в общей точке векторов, начальное положение которых было совершенно различным.
Гомология и аналогия. При параллельной и конвергентной эволюции сходство внешнего строения может быть результатом гомологии — происхождения от общего предка (примером являются конечности разных групп позвоночных животных, рис. 82) или аналогии — независимой эволюции тех систем органов, которые выполняют сходные функции. Ясно, например, что крылья у птиц и насекомых имеют разное происхождение — это пример аналогии. Гомологичные структуры уже в эмбриональный период развиваются по одинаковым генетическим программам. Аналогичные структуры, наоборот, выполняют одинаковые функции, однако не имеют общего генетического базиса. Птицы и мухи летают в одной среде, но не имеют общего крылатого предка и путешествуют в эволюционной истории разными маршрутами.
Дивергенция. Одна из наиболее общих схем эволюционного процесса, реконструированная биологами на основе изучения палеонтологических данных, представлена обычно в виде эволюционного древа с расходящимися ветвями. Это образ дивергентной эволюции, или радиации: общий предок дал начало двум или большему количеству форм, которые, в свою очередь, стали родоначальниками многих видов и родов. Дивергенция (от лат. divergo — отклоняюсь) — расходящаяся эволюция — почти всегда отражает расширение адаптации к новым жизненным условиям. Так, класс млекопитающих распался на многочисленные отряды, представители которых различаются по внешнему строению, особенностям экологии, по характеру физиологических и поведенческих адаптации (насекомоядные, рукокрылые, хищные, китообразные и др.).
Главные линии эволюции. Изменения организмов в процессе эволюции могут иметь различный масштаб и характер. Для понимания исторического развития органического мира важно определить главные линии эволюции. Их выделяют три.
1. Ароморфоз (от греч. airomorphosis — поднимаю форму) — наиболее существенные эволюционные изменения. Такие изменения повышают общий уровень организации, вследствие чего жизнедеятельность организмов усиливается. Ароморфозы дают значительные преимущества в борьбе за существование, делают возможным переход в новую среду обитания. К ароморфозам у животных можно отнести появление живорождения, способности к поддержанию постоянной температуры тела, возникновение замкнутой системы кровообращения, а у растений — появление цветка, сосудистой системы, способности к поддержанию и регулированию газообмена в листьях.
2. Идиоадаптация (от греч. idios — своеобразный и лат. adaptatio — приспособление) — это прогрессивные, но мелкие эволюционные изменения, которые повышают приспособленность организмов к условиям среды обитания. Идиоадаптация не сопровождается изменением основных черт организации, общим подъемом ее уровня и повышением интенсивности жизнедеятельности организма. Примеры идио- адаптаций дает защитная окраска животных или приспособления некоторых рыб (камбала, сом) к жизни у дна — уплощение тела, окраска под цвет грунта, развитие усиков и пр. Другой пример — приспособления к полету у некоторых видов млекопитающих (летучие мыши, белки-летяги).
Примеры идиоадаптации у растений — многообразные приспособления к перекрестному опылению цветка насекомыми или ветром, приспособления к рассеиванию семян. Идиоадаптации приводят к возникновению низших таксономических групп (виды, роды, семейства).
3. Дегенерация (от лат. degenero — вырождение) ведет к упрощению организации, утрате ряда систем и органов и часто связана с переходом к паразитическому образу жизни. Упрощение организации паразита затрагивает прежде всего системы, необходимые для жизни в открытой среде, но лишние внутри хозяина — органы ориентации, пищеварения, движения и т. п.
При общем упрощении организации у паразитов возникают специфические приспособления (часто весьма изощренные) к условиям жизни внутри хозяина. У паразитических червей появляются присоски, крючки, получают значительное развитие органы размножения.
Пути эволюции крупных систематических групп (например, типов и классов) очень сложны. Нередко в развитии этих групп происходит смена эволюционных линий.
Ароморфозы случаются гораздо реже по сравнению с идиоадаптациями и знаменуют, как правило, новый этап развития органического мира. За каждым ароморфозом следует множество идиоадаптации, которые обеспечивают более полное использование всех ресурсов среды и освоение новых местообитаний. У животных, например, крупным ароморфозом при переходе на сушу явилось развитие внутреннего оплодотворения, а также ряд приспособлений к развитию зародыша в яйце на суше (вспомните особенности размножения земноводных, пресмыкающихся).
Птицы и млекопитающие заняли господствующее положение среди наземных животных. Постоянная температура тела позволила им выжить в условиях оледенения и проникнуть далеко в холодные страны. Успешному развитию этих групп способствовали и ароморфозы, и идиоадаптации, которые позволили млекопитающим освоить наземную, а птицам — воздушную среду.
Параллелизм. Конвергенция. Дивергенция. Гомология. Аналогия. Ароморфоз. Идиоадаптация. Дегенерация.
Сайт учителей биологии МБОУ Лицей № 2 г. Воронежа, РФ
Site biology teachers lyceum № 2 Voronezh city, Russian Federation
Эволюция протекает неравномерно, т. е. с разной скоростью в различные периоды истории Земли, но имеет тенденцию к ускорению.
Например, первые живые существа появились около 3,8 млрд лет назад, многоклеточные — 1,3 млрд лет назад, млекопитающие и птицы — 200 млн лет назад, приматы — 60 — 65 млн лет назад, род Человек — около 4 млн лет назад, Человек разумный — около 80 тыс. лет назад.

Эволюция различных групп происходит с разной скоростью. Принято оценивать скорость видообразования в числе поколений. Так, быстрое образование новых видов, связанное с крупными хромосомными перестройками, занимает до нескольких десятков тысяч поколений. Медленное накопление адаптаций даёт новый вид через несколько сотен тысяч поколений.

Эволюция не всегда идёт от простого к сложному, существует и направление, которое сопровождается упрощением строения. Примером такого пути может служить общая дегенерация.
Малая подвижность и пассивный тип питания двустворчатых моллюсков привели к исчезновению головы.
Эволюция является необратимым процессом (правило необратимости эволюции), следовательно, организмы не могут вернуться к прежнему состоянию.

Вернувшиеся в воду ихтиозавры не стали рыбами, а сохранили черты строения рептилий.
У некоторых организмов, в том числе у человека, в онтогенезе, в результате нарушения развития возможно появление отдельных признаков, которые существовали у предков, но были утрачены в процессе эволюции. Эти признаки носят название атавизмы.
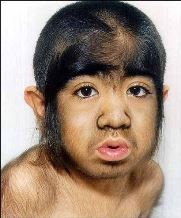
Атавизм (лат. атавус — отдалённый предок) — появление у организмов признаков, отсутствовавших у их ближайших предков, но существовавших у очень далёких предков. Примером атавизма может служить развитие хвостовидного придатка у человека; появление у лошади двух дополнительных пальцев по бокам развитого третьего пальца.
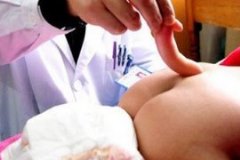
В процессе развития особи до некоторой степени повторяются черты организации предков, но нарушения нормального развития могут привести к тому, что у взрослого организма сохранятся на всю жизнь признаки предков, появляющиеся у зародыша и обычно исчезающие в ходе дальнейшего развития.
Примером такого атавизма может служить появление у человека шейной фистулы, напоминающей жаберную щель предков млекопитающих — рыб и амфибий. Сюда же относятся полимастия у человека (образование большего, чем в норме, числа пар грудных желёз), трёхпалость лошадей и т. д.
К атавизмам относится также возникновение признаков отдалённых предков при регенерации органов. В этом случае последние воссоздаются с признаками, характерными для более древних форм. Например, при регенерации хвоста у ящериц кольца его чешуй иногда образуются в более примитивной форме.
Правило прогрессивной специализации - это явление, в результате которого любая группа живых организмов, эволюционирующая по пути приспособления к конкретным условиям, и в дальнейшем будет продвигаться по пути углубления специализации.
Генетические основы этого правила заключены в том, что в процессе естественного отбора в условиях данной адаптивной зоны из популяции отсеиваются те гены, которые ей не соответствуют.

Примером служит адаптация гиббонов, приспособленных к древесному образу жизни. У них отсутствует хватательный хвост, имеющийся у прочих древесных обезьян, поэтому они передвигаются, прыгая с ветки на ветку при помощи удлиннёных передних конечностей. Большой палец у них значительно редуцирован и рука практически неспособна к манипуляциям мелкими предметами. При передвижении по земле руки гиббонов уже не участвуют в локомоции.
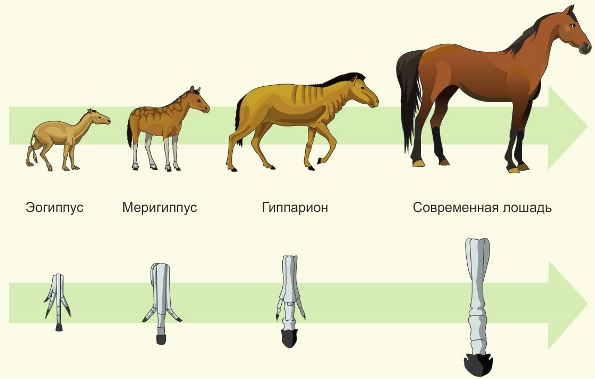
Примером прогрессивной специализации служат морфологические преобразования конечностей в эволюционной ветви лошадей.
При переходе к жизни на открытых пространствах с плотной почвой у предков лошади постепенно уменьшалось количество пальцев, пока не остался один. Эта черта строения не позволяет современным лошадям населять другие биотопы.
Принцип интеграции — объединение отдельных структур в целостный организм. Интеграция — это целесообразное объединение и координация действий разных частей целостной живой системы. Она проявляется в объединении органов в функционально единые системы, обеспечивающие одну из сторон жизнедеятельности организма.
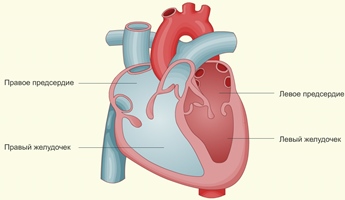
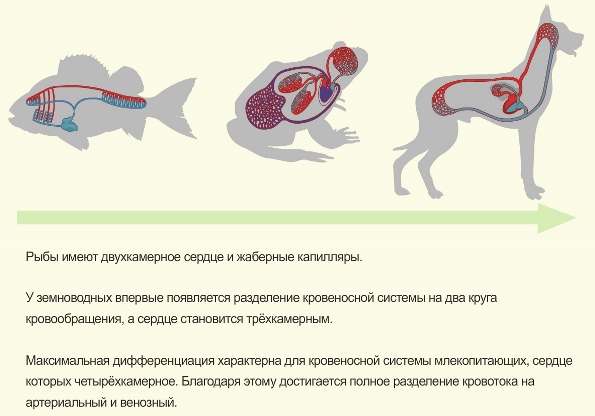
Четырёхкамерное сердце млекопитающих является примером интегрированной структуры: каждый его отдел выполняет определённую функцию, не имеющую смысла в отрыве от функций других отделов.
Принцип дифференциации представляет собой разделение однородной структуры на обособленные части, приобретающие специфическое строение. Таким образом, усложнение структуры всегда связано с усложнением функций и специализацией отдельных частей.
Примером филогенетической дифференциации может являться эволюция кровеносной системы в типе хордовых.

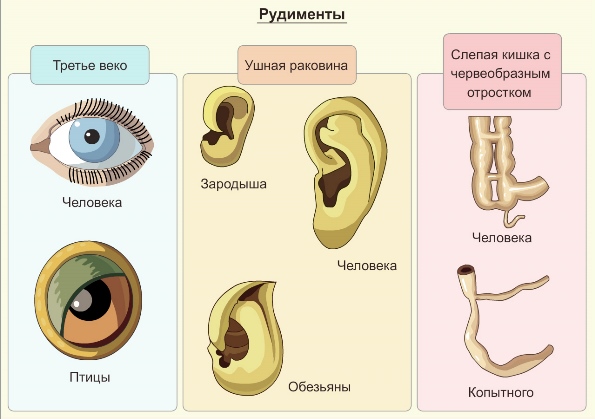
Рудименты (лат. рудиментум — зачаток, первооснова), или рудиментарные органы — сравнительно упрощённые, недоразвитые структуры, утратившие своё основное значение в организме в процессе филогенеза. Рудименты закладываются во время зародышевого развития, но полностью не развиваются.
Примерами рудиментов служат: малая берцовая кость у птиц, глаза у некоторых пещерных и роющих животных, остатки волосяного покрова и тазовых костей у ряда китообразных.
У человека к рудиментам относятся хвостовые позвонки, волосяной покров туловища, ушные мышцы, аппендикс. В отличие от атавизмов рудименты встречаются у всех особей вида.
Таким образом, биологическая эволюция (лат. evolutio – "развертывание") – это процесс постоянного и направленного естественным отбором изменения форм организмов на Земле, обеспечивающий их приспособленность к условиям окружающей среды. Достигается такая приспособленность путем отбора из множества случайных изменений таких, которые облегчают выживание организмов в конкретных условиях среды.
Закономерности эволюции

- Материалом для эволюции служат наследственные изменения — мутации (как правило, генные) и их комбинации.
- Основным движущим фактором эволюции является естественный отбор.
- Единицей эволюции является популяция.
- Эволюция носит в большинстве случаев дивергентный характер, т. е. один таксон может стать предком нескольких дочерних таксонов.
- Эволюция носит постепенный и длительный характер. Видообразование как этап эволюционного процесса представляет собой последовательную смену одной временной популяции чередой последующих временных популяций.
- Вид состоит из подвидов и популяций.
- Вид существует как целостное образование.
- Макроэволюция идёт путем микроэволюции. Для эволюции групп видов живых организмов характерны те же предпосылки и движущие силы, что и для микроэволюции.
- Любой таксон обычно имеет монофилетическое происхождение.
- Эволюция имеет ненаправленный характер, т. е. не идёт в направлении какой-либо конечной цели.
Эволюция – это исторический процесс развития живой природы, который зависит от взаимодействия многих внешних и внутренних факторов при ведущей роли отбора.
Читайте также: