
Реферат на тему генетика популяций
Обновлено: 05.07.2024
Популяция (франц. population – население) – совокупность особей одного вида, занимающих определенный ареал, свободно скрещивающихся друг с другом, имеющих общее происхождение, генетическую основу и в той или иной степени изолированных от других популяций данного вида.
Генетически популяция характеризуется ее генофондом (аллелофондом).
Генофонд представлен совокупностью аллелей, образующих генотипы организмов данной популяции. В каждой данной популяции состав генофонда из поколения в поколение может постоянно изменяться. Новые сочетания генов образуют уникальные генотипы, которые в своем физическом выражении, т. е. в форме фенотипов, подвергаются давлению факторов среды, производящих непрерывный отбор и определяющих, какие гены будут переданы следующему поколению.
Популяция, генофонд которой непрерывно изменяется из поколения в поколение, претерпевает эволюционное изменение. Статичный генофонд отражает отсутствие генетической изменчивости среди особей данного вида и отсутствие эволюционного изменения.
Генофонды природных популяций отличает наследственное разнообразие (генетическая гетерогенность, или полиморфизм), генетическое единство, динамическое равновесие доли особей с разными генотипами.
Наследственное разнообразие заключается в присутствии в генофонде одновременно различных аллелей отдельных генов. Первично оно создается мутационным процессом. Мутации, будучи обычно рецессивными и не влияя на фенотипы гетерозиготных организмов, сохраняются в генофондах популяций в скрытом от естественного отбора состоянии. Накапливаясь, они образуют резерв наследственной изменчивости. Благодаря комбинативной изменчивости этот резерв используется для создания в каждом поколении новых комбинаций аллелей. Объем такого резерва огромен. Так, при скрещивании организмов, различающихся по 1000 локусов, каждый из которых представлен десятью аллелями, количество вариантов генотипов достигает 10 1000 .
Генетическое единство популяции обуславливается достаточным уровнем панмиксии. В условиях случайного подбора скрещивающихся особей источником аллелей для генотипов организмов последовательных поколений является весь генофонд популяций. Генетическое единство проявляется также в общей генетической реализации популяции при изменении условий существования, что обуславливает как выживание вида, так и образование новых видов.
ЗАКОН ХАРДИ – ВАЙНБЕРГА
Любой физический признак определяется одним или несколькими генами. Каждый ген может существовать в нескольких различных формах – аллелях. Число организмов в данной популяции, несущих определенный аллель, определяет частоту данного аллеля (которую иногда называют частотой гена, что менее точно). Например, у человека частота доминантного аллеля, определяющего нормальную пигментацию кожи, волос и глаз, равна 99%. Рецессивный аллель, детерминирующий отсутствие пигментации – так называемый альбинизм, - встречается с частотой 1%. В популяционной генетике частоту аллелей или генов выражают не в процентах, а в десятичных дробях. Таким образом, в данном случае частота доминантного аллеля равна 0,99 , а частота рецессивного аллеля альбинизма – 0,01. Общая частота аллелей в популяции составляет 100%, или 1,0.
Как это принято в классической генетике, аллели обозначаются буквами, например A – доминантный аллель, a – рецессивный и рецессивный) в генофонде популяции, используются символы p и q. Таким образом,

где p – частота доминантного, q – частота рецессивного аллеля. (В примере с пигментацией у человека p = 0,99 , а q =0,01). Значение этого уравнения состоит в том, что, зная частоту одного из аллелей, можно определить частоту другого.
Частоты отдельных аллелей в генофонде позволяют вычислять генетические изменения в данной популяции и определять частоту генотипов.
Поэтому любые изменения частоты аллелей должны быть обусловлены нарушением одного или нескольких перечисленных выше условий. Все эти нарушения способны вызвать эволюционное изменение. Эти изменения и их скорость можно изучить и измерить с помощью уравнения Харди – Вайнберга.
Если имеется два организма, один гомозиготный по доминантному аллелю A, а другой – по рецессивному аллелю a, то все их потомки будут гетерозиготными

Если наличие доминантного аллеля A обозначить символом p, а рецессивного аллеля a – символом q, то картину скрещивания между особями F1, возникающие при этом генотипы и их частоты можно представить следующим образом:

Используя символы p и q результаты приведенного выше скрещивания можно представить следующим образом:
p 2 – доминантные гомозиготы
q 2 – рецессивные гомозиготы.
Такое распределение возможных генотипов носит статистический характер и основано на вероятностях. Три возможных генотипа, образующихся при таком скрещивании, представлены со следующими частотами:
Сумма частот трех генотипов, представленных в рассматриваемой популяции, равна единице; пользуясь символами p и q, можно сказать, что вероятности генотипов следующие:

На математическом языке представляет собой уравнение вероятности, тогда как является квадратом этого уравнения (т.е. ).
p – частота доминантного аллеля;
q – частота рецессивного аллеля;
p 2 – гомозиготный доминантный тип;
2pq – гетерозиготный генотип;
q 2 – гомозиготный рецессивный генотип,
можно вычислить частоты всех аллелей и генотипов, пользуясь выражениями
для частот аллелей:

для частот генотипов:
Однако, для большинства популяций частоту обоих аллелей можно вычислить только по доле особей, гомозиготных по рецессивному аллелю, так как это единственный генотип, который можно распознать по его фенотипическому выражению.
Например, один человек из 10000 – альбинос, т.е. частота альбинотического генотипа составляет 1 на 10000. Поскольку аллель альбинизма рецессивен, альбинос должен быть гомозиготным по рецессивному гену, т.е. на языке теории вероятности

Зная, что q 2 =0,0001, можно определить частоты аллеля альбинизма (q), доминантного аллеля нормальной пигментации (p), гомозиготного доминантного генотипа (p 2 ) и гетерозиготного генотипа (2pq). Так как q 2 =0,0001,

,
т.е. частота аллеля альбинизма в популяции равна 0,01 или 1%. Поскольку

частота доминантного аллеля в популяции равна 0,99 или 99%. А если
и , то

,
т.е. частота гетерозиготного генотипа составляет 0,0198; иными словами, примерно 2% индивидуумов в данной популяции несут аллель альбинизма либо в гетерозиготном либо в гомозиготном состоянии.
Гетерозиготных индивидуумов, нормальных по фенотипу, но обладающих рецессивным геном, который в гомозиготном состоянии может вызвать нарушение метаболизма, называют носителями. Существование в популяции неблагоприятных аллелей в составе гетерозиготных генотипов называют генетическим грузом. Как показывают вычисления с использованием уравнения Харди – Вайнберга, частота носителей в популяции всегда выше, чем можно было бы ожидать на основании фенотипического проявления данного дефекта.
СЛЕДСТВИЯ УРАВНЕНИЯ ХАРДИ – ВАЙНБЕРГА
Однако не все рецессивные аллели неблагоприятны для популяции. Например, у человека из всех групп крови чаще всего встречается группа 0, соответствующая гомозиготности по рецессивному аллелю.
Другим примером служит серновидноклеточная анемия. наследственное заболевание, широко распространенное в ряде областей Африки, индии, у негритянского населения Северной Америки. Индивидуумы, гомозиготные по соответствующему рецессивному аллелю, обычно умирают не достигнув половой зрелости и элиминируя таким образом из популяции по два рецессивных аллеля. Что касается гетерозигот, то они не гибнут. Установлено, что во многих частях земного шара частота аллеля серновидноклеточности остается относительно стабильной. У некоторых африканских племен частота гетерозиготного фенотипа достигает 40%. Раньше думали, что этот уровень поддерживается за счет появления новых мутантов. Однако в результате дальнейших исследований выяснилось, что во многих частях Африки, где среди факторов, угрожающих здоровью и жизни, важное место занимает малярия, люди, несущие аллель серновидноклеточности, обладают повышенной резистентностью к этой болезни. В малярийных районах Центральной Африки это селективное преимущество гетерозиготного генотипа поддерживает частоту аллеля серновидноклеточности на уровне 10 – 20%. У североамериканских негров, уже 200 – 300 лет не испытывающих на себе селективного эффекта малярии, частота аллеля серновидноклеточности упала до 5%.
Этот пример эволюции в действии ясно демонстрирует селективное влияние среды на частоту аллелей – механизм нарушающий равновесие Харди – Вайнберга. Именно такого рода механизмы вызывают в популяциях сдвиги, ведущие к эволюционному изменению.
ФАКТОРЫ, ВЫЗЫВАЮЩИЕ ИЗМЕНЕНИЯ
Принцип Харди – Вайнберга носит чисто теоретический характер. Очень немногие популяции находятся в условиях, при которых сохраняется равновесие.
Половая рекомбинация (кроссинговер во время мейоза, независимое распределение хромосом при мейозе, случайное оплодотворение) обуславливает перетасовку генов, лежащую в основе происходящих непрерывных изменений. Но хотя эти процессы и приводят к образованию новых генотипов и изменяют их частоты, они не вызывают никакого изменения имеющихся аллелей, так что частоты аллелей остаются постоянными.
А эволюционные изменения происходят вслед за появлением новых аллелей, а главным источником последних служат мутации.
Большой вклад в популяционную генетику внес русский ученый С. С. Четвериков, обративший внимание на насыщенность природных популяций рецессивными мутациями, а также на колебания частоты генов в популяциях в зависимости от действия факторов внешней среды.
Мутационный процесс, изменяя частоту одного аллеля по отношению к другому, оказывает на генофонд популяции прямое действие. За счет мутантных аллелей происходит формирование резерва наследственной изменчивости. Благодаря мутационному процессу поддерживается высокий уровень наследственного разнообразия природных популяций. Совокупность аллелей, возникающих в результате мутаций, составляет элементарный эволюционный материал.
Большинство мутаций первоначально оказывает на фенотип особей неблагоприятное действие. В силу рецессивности мутантные аллели обычно присутствуют в генофондах популяций в гетерозиготных по соответствующему локусу генотипах.
Благодаря этому достигается тройственный положительный результат: 1) исключается непосредственное отрицательное влияние мутантного аллеля на фенотипическое выражение признака, контролируемого данным геном; 2) сохраняются нейтральные мутации, не имеющие приспособительной ценности в настоящих условиях существования, но которые смогут приобрести такую ценность в будущем; 3) накапливаются некоторые неблагоприятные мутации, которые в гетерозиготном состоянии нередко повышают относительную жизнеспособность организмов (эффект гетерозиса). Таким образом, создается резерв наследственной изменчивости.
В большинстве природных популяций спаривание происходит неслучайным образом. Во всех тех случаях, когда наличие одного или нескольких наследуемых признаков повышает вероятность успешного оплодотворения гамет, имеет место половой отбор. У растений и животных существует много структурных и поведенческих механизмов, исключающих чисто случайный подбор родительских особей. Например, цветки, у которых лепестки крупнее и нектара больше, чем обычно, вероятно будут привлекать больше насекомых, что повысит вероятность опыления и оплодотворения. Характер окраски насекомых, рыб и птиц и особенности их поведения, связанные с постройкой гнезда, охраной территории и брачными церемониями, повышают избирательность при скрещивании.
Таким образом, половой отбор как механизм избирательного скрещивания обеспечивает некоторым особям более высокий репродуктивный потенциал, в результате чего вероятность передачи генов этих особей следующему поколению повышается. Репродуктивный потенциал особей с менее благоприятными признаками понижен, и передача их аллелей последующим поколениям происходит реже.
О дрейфе генов говорят в тех случаях, когда изменение частоты генов в популяциях бывают случайными и не зависят от естественного отбора. Случайный дрейф генов или генетико-автоматические процессы, или эффект Сьюэлла Райта, может служить важным механизмом эволюционных изменений в небольших и изолированных популяциях. В небольшой популяции могут быть представлены не все аллели, типичные для данного вида.
Генетический дрейф может возникнуть при отделении новой популяции от родительской из-за того, что процент определенных аллелей среди отделившихся особей будет иным, чем в исходной популяции. Например, некий аллель встречается в популяции в 1% случаев. Допустим, что 10 животных из этой популяции были занесены на изолированный остров и там образовали новую популяцию. Если среди этих десяти одно было носителем данного аллеля, то его частота в новой популяции будет составлять уже 10%. Если же не было ни одного, то в образовавшейся популяции этот аллель будет отсутствовать.
Или же, например, если данный аллель встречался в популяции из 1000000 особей с частотой, допустим 1% (т.е. q = 0,01), то им будет обладать 10000 особей; а в популяции состоящей из 100 особей, этот аллель будет иметься только у одной особи, так что вероятность его случайной утраты в малой популяции (например, в результате преждевременной гибели особи – единственного носителя данного аллеля в популяции) будет гораздо выше.
С дрейфом генов связано явление, известное под названием принципа основателя. Оно состоит в том, что при отделении от родительской популяции небольшой ее части последняя может случайно оказаться не вполне типичной по своему аллельному составу. Некоторые аллели в ней могут отсутствовать, а другие будут представлены с непропорционально высокой частотой. Постоянное скрещивание внутри такой пионерной популяции приведет к созданию генофонда, отличающегося по частотам аллелей от генофонда исходной родительской популяции. Дрейф генов обычно снижает генетическую изменчивость в популяции, главным образом в результате утраты тех аллелей, которые встречаются редко. Суть дрейфа генов в том, что из-за случайных колебаний один из аллелей может вообще выпасть из популяции и она окажется по данному гену гомозиготной. В результате происходит гомозиготизация особей и затухание изменчивости. Это открывает путь случайной дифференцировке популяций вплоть до видового обособления.
Поток генов – это перемещение аллелей из одной популяции в другую в результате скрещивания между членами этих двух популяций. Случайное внесение новых аллелей в популяцию – реципиента и удаление их из популяции – донора изменяет частоту аллелей в обеих популяциях и ведет к повышению генетической изменчивости. Несмотря на то, что поток генов вносит в популяции генетическую изменчивость, в смысле эволюционного изменения его действие оказывается консервативным. Распространяя мутантные аллели по всем популяциям, поток генов приводит к тому, что все популяции данного вида приобретают общий генофонд, т.е. различия между популяциями уменьшаются.
Таким образом, изменение частоты генов, вызванные теми или иными факторами внешней среды, служат основой возникновения различий между популяциями и в дальнейшем обуславливают преобразование их в новые виды. Поэтому изменения популяций в ходе естественного отбора называют микроэволюцией.
Видообразование – это качественный этап эволюционного процесса .
Это означает, что образованием видов завершается микроэволюция и
Микроэволюцию можно рассматривать как эволюцию популяций
–;открытых генетических систем, способных обмениваться генетическим
материалом, а макроэволюцию – как эволюцию совершенных таксонов
–;закрытых генетических систем, которые не способны обмениваться генами
в естественных условиях. Вид занимает промежуточное положение между
открытыми и закрытыми генетическими системами, поэтому биологич еский
вид можно рассматривать как ;устойчивую генетическую систему,
эволюционирующую относительно независимо от других подобных систем.
В то же время биологический вид представляет собой основную
единицу сообщества, поскольку именно виду соответствует собственная
Таким образом, для понимания сущности видообразования необходимо
рассмотреть его популяционно-генетические и экологические аспекты.
Видообразование – это сложный, не изученный до конца процесс.
Известно множество механизмов образования новых видов. Но в любом
случае новый вид должен отличаться от материнского или сестринского вида
хотя бы некоторыми наследуемыми признаками и, следовательно, хоты бы
некоторыми аллелями. Таким образ ом, в ходе видообразования исходный
набор аллелей должен быть замещен новым набором аллелей.
Согласно общепринятой точке зрения, в основе видообразования лежит
принцип дивергенции. В результате дивергенции увеличивается число видов.
Дивергенция ( от лат. ;divergo;– отклоняюсь, отхожу) – это расхождение
признаков организмов в ходе эволюции разных групп (филетических линий),
возникших от одного предка. Дивергенцией называют также разделение в
процессе эволюции единого таксона на два или несколько. Термин
видообразование затруднено. Например, в средней полосе европейской части
России м ногие роды высших растений и крупных животных представлены
или одним видом (ель, сосна, дуб, лещина; липа, медведь, волк, лиса,
белка…), или немногими видами (березы пушистая и бород авчатая; заяц-
беляк и заяц- русак…). Это связано с тем, что при большой и постоянной
численности популяций элементарные эволюционные факторы практически
Таким образом, для начала видообразования исходная большая
популяция (мегапопуляция) должна быть разделена на множество
малочисленных изолированных популяций (дробление крупных популяций
называется;инсуляризацией). Такая изоляция является первичной и носит

Связано с расхождением групп особей одного вида и обитающих на
одном ареале по экологическим признакам. При этом особи с
промежуточными характеристиками оказываются менее приспособленным и.
Расходящиеся группы формиру ют новые виды. Си м патрическое
видообразование м ожет протекать несколькими способами. Один из них —
возникновение новых видов при быстром изменении кариотипа путём
полиплоидизации. Известны группы близких видов, обычно растений, с
кратным числом хромосом. Другой способ симпатрического
видообразования — гибридизация с последующим удвоением числа
хромосом. Сейчас известно немало видов, гибридогенное происхождение и
характер г енома которых м ожет считаться экспериментально доказанным.
Третий способ симпатрического видообразования — возникновение
репродуктивной изоляции особей внутри первоначально единой популяции в
результате фрагментации или слияния хромосом и других хромосомных
перестроек. Особенностью симпатрического пути видообразования является
то, что он приводит к возникновению новых видов, всегда морфологически
близких к исходному виду. Лишь в случае гибридогенного возникновения
видов появляется новая видовая форма, отличная от каждой из родительских.
Рефераты и конспекты лекций по географии, физике, химии, истории, биологии. Универсальная подготовка к ЕГЭ, ГИА, ЗНО и ДПА!
Популяция - это совокупность свободно скрещивающихся особей одного вида, которая длительно существует на обособленной территории относительно изоолированно от других совокупностей особей того же вида. Совокупность генов популяции в данный период времени называется генофондом популяции.
В отличие от генома индивидуального диплоидного организма, в котором может быть только два аллельных гена, в генотипе популяции может быть больше чем два аллельных генов. Количество каждого вида генов в популяции описывается величиной, аналогичной концентрации для молекул, называемой частотой гена, которая определяется как отношение числа генов данного вида к общему числу всех аллельных для этого вида генов в генофонде популяции.
Частоты генов в генофонде постоянно меняются под действием мутаций, естественного отбора, миграции организмов из одной популяции в другую и из-за случайных причин. Однако в достаточно большой популяции (когда случайными изменениями можно пренебречь), при отсутствии мутаций, естественного отбора и миграций, а также при свободном скрещивании особей и отсутствие предпочтений в спаривании особей при половом размножении частоты генов, а следовательно, и генотипов остаються постоянными из поколения в поколение. Это явление называется законом генетического равновесия в популяциях Харди-Вайнберга.
Предположим, что в популяции есть только два вида аллельных генов А и а, и что в данном поколении популяции частоты этих генов в генофонде составляют p и q соответственно (p + q = 1). Тогда в соответствии с законом Харди-Вайнберга эти частоты и частоты генотипов АА, Аа и аа, которые равны p2, 2pq и q2 соответственно, будут оставатися постоянными во всех последующих поколениях. Популяция, подчиняющаяся закону Харди- Вайнберга, является идеализированной подобно модели идеального газа. Однако есть случаи, когда этот закон достаточно хорошо выполняется на практике. Например, при наследовании групп крови системы АВ0, о которой подробно будет рассказано дальше. Популяция, подчиняющаяся закону Харди-Вайнберга, - это статическая популяция.
Рассмотрим теперь динамику популяции, на которую постоянно действуют факторы, изменяющие частоты генов в генофонде. Практически всегда природные популяции испытывают давление естественного отбора, который действует через приспособленность организмов. Приспособленность данного генотипа (WG) можно определить как число зигот, продуцируемых в данном поколении к числу зигот предыдущего поколения. WG слагается из жизнеспособности генотипа (vG, вероятность завершения онтогенеза зиготой с генотипом G), половой активности (rG, верояность скрещивания особей, имеющих генотип G) и плодовитости (fG, среднее число зигот, продуцируемое особями с генотипом G): WG = vG7rG7fG. Например, если в данном поколении было 1000 зигот генотипа G, из них 45% прошли ранние эмбриональные фазы, 1/3 от оставшихся достигли половой зрелости, 2/3 от оставшихся вступили в скрещивания и произвели в среднем по 15 зигот, то WG = 1.5. Поскольку реальные значения приспособленности генотипов найти трудно, то обачно используют относительные величины приспособленности, принимая приспособленность одного из генотипов за 1.
Рассмотрим популяцию с двумя аллельными генами А и а. Примем за 1 приспособленность гетерозигот: WAa= 1. Тогда WAA= 1-s и Waa= 1-t, где s и t - относительные коэффициенты отбора (могут быть как положительными, так и отрицательными). Частоты генотипов АА, Аа и аа, которые в данном поколении были p2, 2pq и q2 соответственно, в следующем поколении (после отбора) будут составлять p2WAA, 2pqWAa и q2Waa. Величина WM = p2WAA+ 2pqWAa+ q2Waa = 1-(sp2+ tq2) называется средняя приспособленность популяции. Новые частоты генов А и а вы- разятся как p'=(p2WAA+ pqWAa)/WM и q'=(pqWAa+ q2Waa)/WM, откуда p'=p(1-sp)/(1-(sp2+ tq2)). Реккурентное соотношение p'=F(p), связывающее частоту гена в данном поколении с частотой этого гена в предыдущем уравнении, называется уравнением динамики популяции.
Изменяясь из поколения в поколение, частота p в конце концов может стабилизироваться и дальше оставаться постоянной. Такая частота (p*) называется стационарной частотой и ее можно найти из уравнения p*=F(p*). Однако найденная таким образом стационарная частота может быть устойчивой и неустойчивой. Устойчивая стационарная частота характеризуется тем, что после небольшого отклонения ее от величины p* она возвращается к этой же величине в последующих поколениях. Напротив, в случае неустойчивой стационарной частоты даже незначительное отклонение от p* в последующих поколениях продолжает увеличиваться, и частота p достигает нового стационарного значения. Естественно, что в природе реализуются только устойчивые стационарные частоты. Условие устойчивости находим разложив F(p) в ряд Тейлора в окрестности точки p*: F(p) ~ F(p*) + F'(p*)(p-p*) = p* + F'(p*)(p-p*). Частота гена в следующем поколении будет: p' = F(p) ~ p* + F'(p*)(p-p*), откуда p'- p* ~ F'(p*)(p-p*). Точка будет стационарной, когда отклонение p'-p* будет меньше, чем отклонение p-p*, т.е. при условии 2F'(p*)2 0 и t>0, т.е. когда приспособленность гетерозигот выше, чем обоих гомозигот. Если в популяции происходит мутация, например, А 6 а с частотой m, то если частота гена А в данном поколении равна p, через n поколений она будет составлять pn = p(1-m)n.
Частоты мутаций, как правило, лежат в пределах 10-6- 10-4. Мутации служат материалом для отбора, поэтому представляет интерес рассмотреть уравнение динамики популяции с мутацией и отбором по этой мутации: p'=p(1-m)(1-sp)/(1-(sp2+ tq2)).
Если мутация летальная: WAA= WAa=1 и Waa= 0, то s=0 и t=1. В этом случае уравнение динамики популяции упрощается: p = p(1-m)/(1-q2) = (1-m)/(1+q) и имеет единственную устойчивую стационарную частоту 1-p* = q* = m1/2. Это показывает, что даже такая вредная мутация, как летальная, не может быть полностью вытеснена отбором и продолжает сохраняться в популяции в гетерозиготном состоянии. Если же мутация не летальная, а просто вредная - понижающая приспособленность генотипа аа: WAA= WAa=1, Waa= 1-t, 0

Что такое популяция
Популяция - это термин, который происходит от латинского — populatio, т.е. население. Он обозначает совокупность организмов относящихся к одному виду, которые длительное время обитают на определённой территории (имеют общий ареал обитания) и полностью либо частично изолированны от особей других аналогичных групп того же вида.

Для чего нужна популяционная генетика
Интерес изучения генетики на популяционном уровне заключается в том, что через популяционный уровень осуществляется генетическая преемственность поколений, а также регуляция таких биологически важных свойств, как численность, плодовитость, устойчивость к заболеваниям и пр.
Таким образом, популяционная генетика — это раздел генетической науки, который изучает распределение частот аллелей (различных форм одного и того же гена, расположенных в одинаковых участках хромосом), а также их изменение под влиянием движущих сил эволюции, определяющих генетическую структуру популяции.
Она является одним из главных элементов синтетической теории эволюции. Механизмы, которые определяют генетическую структуру популяций, можно разделить на две группы:
- поддерживающие равновесие,
- те, которые, его нарушают.
К первым относятся естественный отбор и дрейф генов, а ко вторым мутагенез и поток генов. При этом естественный отбор и мутагенез оказывают систематическое воздействие, а дрейф и поток генов – случайное. Тем самым, популяционная генетика объясняет процессы адаптации и видообразования .
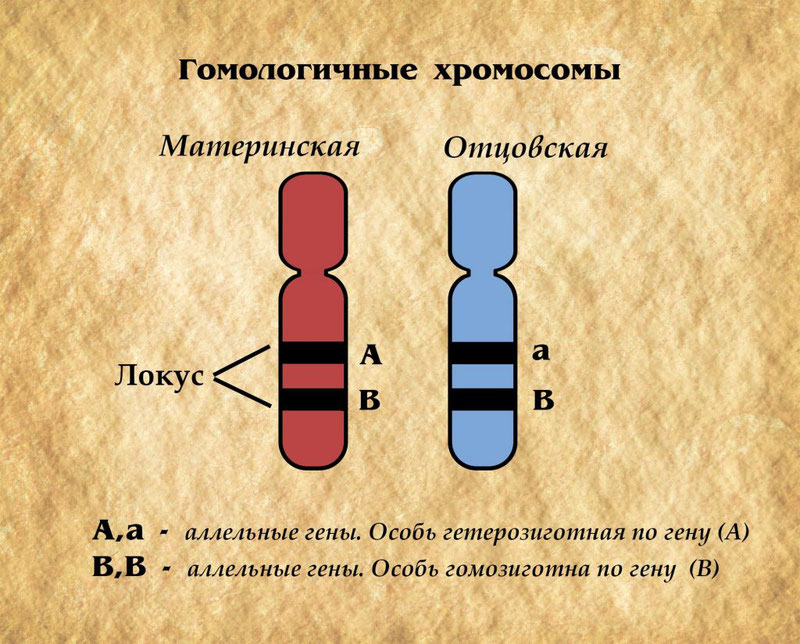
В общем исследования в области генетики популяций можно условно разделить на два направления:
- описание генетического состава популяций
- анализ причин изменения генофонда популяции.
Закон Харди-Вайнберга
Рассмотрение каждого из механизмов определяющих генетическую структуру популяции, заслуживает отдельной статьи, но понимание их невозможно без рассмотрения ключевого закона популяционной генетики – закона Харди-Вайнберга или закона равновесного состояния. На нем и остановим своё внимание.
Итак, механизмы этого закона был открыты в 1908 г. английским математиком Харди (Hardy) и немецким врачом (Weinberg) независимо друг от друга.
Для его понимания рассмотрим такое понятие как случайное скрещивание.
Случайное скрещивание – это скрещивание при котором генетическая структура особей не влияет на вероятность формирования брачных пар между ними. Таким образом, в случайно скрещивающихся популяциях частота спариваний носителей различных генотипов пропорциональна доле, частотности данных генотипов в популяции.
Итак, согласно закону Харди-Вайнберга при отсутствии элементарных эволюционных процессов (естественного отбора, мутаций, дрейфа генов и миграции) частоты генов из поколения в поколение остаются неизменными. Соответственно если скрещивания случайны, частота генотипа связана простым квадратичным соотношением с частотой генов (аллелей). Если речь идет об аутосомном локусе, то равновесие генов достигается за одно поколение, и если дополнительные вмешательства, нарушающие структуру популяции, отсутствуют, сохраняется во всех последующих поколениях. Если же мы говорим о сцепленных с полом локусах, равновесные частоты генотипов устанавливаются постепенно.

Годфри Харолд Харди (1877-1947)
Таким образом равновесные частоты генотипов определяются произведениями частот соответствующих аллелей. В случае наличия только двух аллелей А (с частотой встречаемости p) и а (с частотой - q), частоты каждого их трех возможных генотипов будут выражены уравнением:
(p + q) 2 = p 2 + 2pq + q 2 = 1.
Если мы говорим о трех аллелях А, а, А’ с частотами p, q, r, формула будет иметь вид:
(p + q + r) 2 = p 2 + q 2 + r 2 + 2pq + 2pr + 2qr = 1.
Этот прием возведения в квадрат многочлена может быть использован для определения равновесных частот генотипов при любом числе аллелей. При этом сумма частот генотипов (как и аллелей) всегда должна равняться единице.

Задачи на закон Харди-Вайнберга не всегда просто даются начинающим генетикам. Помните, что Вы всегда можете обратиться к нашим авторам за квалифицированной и своевременной помощью! Сделайте процесс своего обучения приятным и легким вместе с Zaochnik!
Иван Колобков, известный также как Джони. Маркетолог, аналитик и копирайтер компании Zaochnik. Подающий надежды молодой писатель. Питает любовь к физике, раритетным вещам и творчеству Ч. Буковски.

ГОСТ
Генетика популяций
Одной из важных отраслей генетики является генетика популяций. Эта отрасль науки посвящена изучению генетической структуры природных популяций, а также генетических процессов, происходящих в ней. Она играет исключительно важную роль для развития эволюционной теории. Благодаря достижениям генетики популяций установлено, что популяция это не просто единица вида, но и единица эволюции.
Основоположником работ в этом направлении первой половине $XX$ века ($20$-е годы) стал С.С. Четвериков, русский генетик. Также, весомый вклад в развитие этой науки внесли:
- С.М. Гершинзон,
- Н.П. Дубинин,
- Д.Д.Ромашов,
- Р.Райт,
- Р.Фишер,
- Ф.Г.Добржанский.
Что же такое популяция? Современное определение этого понятия выглядит так:
Закон Харди-Вайнберга
Так как популяция состоит из особей одного вида, то все они обладают одинаковым набором генов. Однако любой ген может быть представлен разными аллелями, количество которых может быть значительным. Значит, популяция – это совокупность неодинаковых в генетическом отношении особей, которые отличаются разными состояниями свойственных им признаков. В природе разные популяции одного и того же вида могут отличаться частотами встречаемости определенных аллельных генов.
Исследование закономерностей, которые распределяют аллели в популяциях, является важной задачей популяционной генетики. Над этой проблемой работали немецкий ученый В. Вайнберг и английский исследователь Г. Харди. Они сделали вывод, что у достаточно многочисленной популяции, особи которой свободно скрещиваются между собой свободно, и без влияния какого-либо внешнего фактора на определенные сочетания аллелей, новые мутации не возникают , и обмен генетической информацией с другими популяциями в результате миграции особей в другие популяции не происходит, соотношение аллелей стабилизируется на протяжении нескольких поколений, затем на длительное время остается постоянным.
Готовые работы на аналогичную тему
Закономерность, установленная Г. Харди и В. Вайнбергом, говорит о том, что при условии постоянства внешних и внутренних факторов частоты встречаемости аллелей у достаточно многочисленной популяции, которая изолированна от других, остается достаточно устойчивым на протяжении длительного периода.
Ученые предложили формулу, с помощью которой можно описать распределение частот в панмиксической популяции по одной паре аллельных генов ($A – a$). Эту формулу используют в качестве простейшей модели, служащей основой для проведения более сложных популяционно-генетических исследований.
$p^2AA + 2pqAa + q^2aa = 1$
Уравнение Харди-Вайнберга
- $q$ - частота встречаемости рецессивного гена;
- $p$ - частота встречаемости доминантного гена;
- $q^2$ - частоста встречаемости генотипа $aa$;
- $p^2$ - частоста встречаемости генотипа $AA$;
- $2pq$ - частота встречаемости генотипа $Aa$.
Исследования показали, что только при отсутствии свободного скрещивания, можно существенно повлиять на распределение частот встречаемости генотипов потомков.
Кроме того на изменения частот могут повлиять интенсивные миграции особей из между популяциями. Это приводит к активному обмену разными аллелями.
Мутации, происходящие в популяциях, также дают отклонение в показателях частот встречаемости аллелей.
Дрейф генов
Среди главных причин изменений генетической структуры популяций можно выделить дрейф генов.
Дрейф генов – это случайное ненаправленное изменение частоты встречаемости в популяциях аллелей.
Очень четко это явление проявляется в популяциях с немногочисленными особями, благодаря существующему ограничению свободного спаривания во время размножения. Но чем больше численность популяции, тем меньшее влияние на изменение частот встречаемости аллелей имеет дрейф генов.
Генетическая структура популяций
Популяции между собой отличаются частотой различных сочетаний аллельных генов.
Совокупность всех генов и их аллелей особей определенной популяции именуется генофондом.
Особое значение в генетике имеют рецессивные мутации. Рецессивные признаки распространяются в популяции в гетерозиготном состоянии, в фенотипе не проявляются. Но со временем увеличивается вероятность образования гомозигот по рецессивному признаку. Тогда эти признаки проявляются в фенотипе. Если признак окажется полезным или нейтральным, он сохранится, если вредным – его носители просто погибнут. Поэтому рецессивные мутации называют резервом наследственной изменчивости популяций.
Читайте также: