
Реферат митохондрии как энергетические станции клеток
Обновлено: 04.07.2024
Человеческие клетки содержат в среднем 1500 митохондрий.Их особенно много в клетках с интенсивным метаболизмом (например, в мускулах или печени).
Митохондрии подвижны и перемещаются в цитоплазме в зависимости от потребностей клетки. Благодаря наличию собственной ДНК они размножаются и самоуничтожаются независимо от деления клетки.
Клетки не могут функционировать без митохондрий, без них жизнь не возможна.
Различный типы клеток отличаются друг от друга как по количеству и форме митохондрий, так и по количеству крист. Особенно много крист имеют митохондрии в тканях с активными окислительными процессами, например в сердечной мышце. Вариации митохондрий по форме, что зависит от их функционального состояния, могут наблюдаться и в тканях одного типа. Митохондрии — изменчивые и пластичные органеллы.
Специфическая ДНК:Самая примечательная особенность митохондрий - это наличие у них своей собственной ДНК: митохондриальной ДНК. Независимо от ядерной ДНК, каждая митохондрия имеет свой собственный генетический аппарат.Как и следует ожидать исходя из ее названия, митохондриальная ДНК (мтДНК) находится внутри митохондрий, небольших образований, располагающихся в цитоплазме клетки, в отличие от ядерной ДНК, упакованной в хромосомы внутри ядра. Митохондрии имеются у большинства эукариот и имеют единое происхождение, как считается, от одной древней бактерии, когда-то на заре эволюции поглощенной клеткой и превратившейся в ее составную часть, которой были "поручены" очень важные функции. Митохондрии часто называют "энергетическими станциями" клеток по той причине, что внутри них вырабатывается аденозинтрифосфорная кислота (АТФ), химическую энергию которой клетка может расходовать практически везде, подобно тому как человек использует в своих целях энергию топлива или электричества. И точно так же, производство топлива и электричества требует немалых затрат человеческого труда и слаженной работы большого количества специалистов, получение АТФ внутри митохондрии (или "клеточное дыхание", как его называют) использует огромную массу клеточных ресурсов, включая "топливо" в виде кислорода и некоторых органических веществ, и конечно предполагает участие в этом процессе сотен белков, каждый из которых выполняет свои специфические функции.
Назвать этот процесс просто "сложным" будет наверное мало, потому что прямо или косвенно он связан с большинством других процессов обмена веществ в клетке, за счет того, что эволюция наделила каждый "винтик" данного механизма множеством дополнительных функций. Основной принцип заключается в создании условий, когда внутри митохондриальной мембраны становится возможным присоединение еще одного фосфата к молекуле АДФ, что "энергетически" нереально в обычных условиях. И наоборот, последующее использование АТФ заключается в возможности разрыва этой связи с освобождением энергии, которую клетка может использовать в своих многочисленных целях. Строение митохондриальной мембраны очень сложное, включает большое количество белков различного вида, которые объединяются в комплексы, или как говорят, "молекулярные машины", выполняющие строго определенные функции. Биохимические процессы, протекающие внутри митохондриальной мембраны (трикарбоновый цикл и др.), на входе принимают глюкозу, в числе продуктов выхода дают углекислый газ и молекулы НАДН, способные отщеплять атом водорода, передавая его белкам мембраны. При этом протон переносятся на наружную сторону мембраны, а электрон в конечном счете забирает молекула кислорода на внутренней стороне. Когда разница потенциалов достигает определенной величины, протоны начинают двигаться внутрь клетки через специальные белковые комплексы, и соединяясь с молекулами кислорода (уже получившими электрон), образуют воду, а энергия движущихся протонов используется при образовании АТФ. Таким образом, на вход всего процесса поступают углеводы (глюкоза) и кислород, а выходом являются углекислота, вода и запас "клеточного топлива" - АТФ, которое может транспортироваться в другие части клетки.
Как уже упоминалось выше, все эти функции митохондрия унаследовала от своего предка - аэробной бактерии. Поскольку бактерия является самостоятельным одноклеточным организмом, внутри ее имеется молекула ДНК, в которой записаны последовательности, определяющие строение всех белков данного организма, то есть, прямо или косвенно - все функции, им выполняемые. Когда бактерия-протомитохондрия и древняя эукариотическая клетка (по происхождению - тоже бактерия) слились, новый организм получил две различающиеся молекулы ДНК - ядерную и митохондриальную, которые, по-видимому, поначалу кодировали два совершенно самостоятельных жизненных цикла. Однако внутри новой единой клетки такое обилие обменных процессов оказалось ненужным, так как они во многом дублировали друг друга. Постепенное взаимное приспособление двух систем привело к замещению большинства белков митохондрии собственными белками эукариотической клетки, способными выполнять аналогичные функции. В результате участки кода митохондриальной ДНК, прежде выполнявшие определенные функции, переходили в разряд некодирующих и со временем терялись, приводя к сокращению молекулы. Благодаря тому, что некоторые формы жизни, например грибы, имеют весьма длинные (и полностью функционирующие!) цепочки митохондриальной ДНК, мы можем судить об истории упрощения этой молекулы достаточно достоверно, наблюдая, как в течение миллионов лет в разных ответвлениях Древа Жизни терялись те или иные ее функции. Современные хордовые, в том числе млекопитающие, имеют мтДНК длиной от 15000 до 20000 нуклеотидов, оставшиеся гены которой расположены очень плотно друг к другу. Лишь немногим более 10 белков и всего два типа структурной РНК кодируется в самой митохондрии, все остальное, что требуется для клеточного дыхания (это более 500 белков) предоставляется ядром. Единственная, пожалуй, сохранившаяся целиком подсистема - это транспортные РНК, гены которых до сих пор лежат в митохондриальной ДНК. Транспортные РНК, в состав каждой из которых входит трехнуклеотидная последовательность, служат для синтеза белков, одной стороной "читая" трехбуквенный кодон, задающий будущий белок, а другой присоединяя строго определенную аминокислоту; само соответствие между трехнуклеотидными последовательностями и аминокислотами называется "таблицей трансляции" или "генетическим кодом". Транспортные РНК митохондрий участвуют только в синтезе митохондриальных белков и не могут использоваться ядром, потому что между ядерным и митохондриальным кодами за миллионы лет эволюции накопились небольшие различия.
Также упомянем, что существенно упростилась сама структура митохондриальной ДНК, так как многие составные части процесса транскрипции (чтения) ДНК были утеряны, вследствие чего исчезла необходимость в особом структурировании митохондриального кода. Белки-полимеразы, осуществляющие транскрипцию (чтение) и репликацию (удвоение) митохондриальной ДНК, кодируются не в ней самой, а в ядре.
Главной и непосредственной причиной разнообразия форм жизни являются мутации кода ДНК, то есть замены одного нуклеотида другим, вставки нуклеотидов и их удаления. Как и мутации ядерной ДНК, мутации мтДНК в основном происходят при размножении молекулы - репликации. Однако циклы деления митохондрий независимы от делений клетки, в связи с чем мутации в мтДНК могут возникать независимо от клеточных делений. В частности, между мтДНК, находящимися в разных митохондриях внутри одной клетки могут быть некоторые минорные различия, равно как и между митохондриями в разных клетках и тканях одного организма. Это явление называется гетероплазмией. В ядерной ДНК точного аналога гетероплазмии нет: организм развивается из единственной клетки, содержащей единственное ядро, где весь геном представлен одной копией. В дальнейшем в течение жизни индивида различные ткани могут накапливать т.н. соматические мутации, но при этом все копии генома в конечном счете происходят от одной. Ситуация с митохондриальным геномом несколько иная: зрелая яйцеклетка содержит сотни тысяч митохондрий, которые, делясь, могут быстро накапливать небольшие различия, причем весь набор вариантов передается по наследству новому организму после оплодотворения. Таким образом, если несовпадения между вариантами ядерной ДНК различных тканей вызваны только соматическими (прижизненными) мутациями, то различия митохондриальных ДНК - как соматическими, так и герминальными (зародышевыми) мутациями.
Еще одним отличием является то, что молекула митохондриальной ДНК - кольцевая, в то время ядерная ДНК упакована в хромосомы, которые можно (с некоторой степенью условности) рассматривать как линейные последовательности нуклеотидов.
Наконец, последней особенностью митохондриальной ДНК, которую мы упомянем в данном вводном разделе, является ее неспособность к рекомбинации. Иными словами, между различными эволюционными вариантами митохондриальной ДНК одного вида невозможен обмен гомологичными (т.е. сходными) участками, и поэтому вся молекула изменяется только путем медленного мутирования в течение тысячелетий. У всех хордовых животных митохондрии наследуются только от матери, поэтому эволюционное древо митохондриальной ДНК соответствует генеалогии по прямой женской линии. Однако данная особенность не является уникальной, в различных эволюционных семействах те или иные ядерные хромосомы также не подвержены рекомбинации (не имея пар) и наследуются только от одного из родителей. Так. например, Y-хромосома у млекопитающих может быть передана только от отца к сыну. Митохондриальная ДНК наследуется только по материнской линии и передается из поколения в поколение исключительно женщинами Эта особая форма наследственности митохондриального генома позволила создать родословное древо разных человеческих этносов, определив местонахождение наших общих предков в Эфиопии около 200 000 лет назад.Обладая необыкновенными способностями к адаптации, при увеличении потребности в энергии митохондрии также способны размножаться независимо от клеточного деления. Это явление возможно благодаря митохондриальной ДНК .Митохондриальная ДНК передается исключительно женщинами Митохондриальная ДНК не наследуется по законам Менделя, а по законам цитоплазматического наследования. Во время оплодотворения проникающий в яйцеклетку сперматозоид теряет жгутик, в котором находятся все митохондрии. Зародышу передаются только митохондрии, содержащиеся в яйцеклетке матери. Таким образом, клетки наследуют их единственный источник энергии из материнских митохондрий.Митохондрия: энергетическая станция клеткиУникальный источник энергии.В обычной жизни существуют различные способы извлечения энергии и использования ее для бытовых нужд: солнечные батареи, атомные электростанции, ветроэлектростанции. У клетки есть только одно решение для извлечения, преобразования и хранения энергии: митохондрия. Только митохондрия может преобразовать различные виды энергии в АТФ, энергию, используемую клеткой.
Процесс преобразования клеточной энергииМитохондрии используют 80% кислорода, который мы вдыхаем, чтобы преобразовывать потенциальную энергию в энергию, используемую клеткой. В процессе окисления освобождается большое количество энергии, которая сохраняется митохондриями в виде молекул АТФ.
В 2000 году было доказано, что митохондрии ускоряют процесс фотостарения. На участкахкожи, регулярно подвергающихся воздействию солнечных лучей, уровень мутаций ДНК значительно выше, чем на защищенных участках.Сравнение результатов биопсии (взятие образцов кожи для анализа) участка кожи, подвергшегося воздействию ультрафиолетовых лучей , и защищенного участка показывает, что мутации митохондрий под воздействием УФ-излучения вызывают хронический окислительный стресс.Клетки и митохондрии навсегда связаны между собой: энергия, поставляемая митохондриями, необходима для деятельности клеток. Поддерживать деятельность митохондрий необходимо для лучшей клеточной деятельности и для улучшения качества кожи, особенно кожи лица, слишком часто подвергающейся воздействию УФ-лучей.
Поврежденная митохондриальная ДНК за несколько месяцев порождает более 30 подобных себе митохондрий, т.е. с теми же повреждениями.
Восстановление функций метахондрия и ограничение процессов приводящих к старению возможно при применении коэнзима Q10. В результате проведенных экспериментов было установлено замедление процессов старения и увеличение продолжительности жизни у некоторых многоклеточных организмов в результате введения добавок CoQ10.
Хотя наш организм может вырабатывать CoQ10, он не всегда производит его в достаточном количестве. Поскольку мозг и сердце входят в число самых активных тканей организма, дефицит CoQ10 негативно влияет на них в наибольшей степени и может привести к серьезным проблемам с этими органами. Дефицит CoQ10 могут вызвать различные причины, в том числе плохое питание, генетические или приобретенные дефекты и, к примеру, повышенная тканевая потребность. Сердечно-сосудистые заболевания, включая высокие уровни холестерина и повышенное артериальное давление, также требуют увеличения уровня CoQ10 в тканях. Кроме того, поскольку уровни CoQ10 снижаются с возрастом, людям старше 50 лет может потребоваться больше этого вещества. Многие исследования показали, что ряд лекарственных препаратов (прежде всего гиполипидемические лекарственные средства, такие как статины) снижают уровень CoQ10.
Учитывая ключевую роль CoQ10 в митохондриальной функции и защите клеток, этот кофермент может быть полезен при наличии целого ряда проблем со здоровьем. CoQ10 может принести пользу при наличии такого широкого перечня болезней, что нет никаких сомнений в его важности как питательного вещества. CoQ10 является не только антиоксидантом общего действия, но и может помочь при наличии следующих заболеваний:
Сердечно-сосудистые заболевания: высокое кровяное давление, застойная сердечная недостаточность, кардиомиопатия, защита во время хирургических операций на сердце, высокий уровень холестерина, который лечат с помощью медикаментов, особенно статинов
Рак (для повышения иммунной функции и/или компенсации побочных эффектов химиотерапии)
Сахарный диабет
Мужское бесплодие
Болезнь Альцгеймера (профилактика)
Болезнь Паркинсона (профилактика и лечение)
Парадонтоз
Дегенерация желтого пятна
Исследования на животных и людях подтвердили полезность CoQ10 при всех вышеуказанных заболеваниях, особенно сердечно-сосудистых. В самом деле, исследования показали, что 50–75 процентов людей с различными заболеваниями сердечно-сосудистой системы страдают от дефицита CoQ10 в сердечных тканях. Устранение этого недостатка часто может приводить к драматическим результатам у пациентов с какой-либо болезнью сердца. Например, было доказано, что дефицит CoQ10 имеет место у 39 процентов пациентов с высоким артериальным давлением. Только этот вывод обуславливает необходимость приема добавок с CoQ10. Однако, как представляется, преимущества CoQ10 не ограничиваются устранением сердечно-сосудистой недостаточности.
Проведенное в 2009 году исследование, результаты которого были опубликованы в журнале Pharmacology & Therapeutics, дает основания предположить, что воздействие CoQ10 на артериальное давление становится заметным только через 4–12 недель после лечения и типичное снижение систолического и диастолического артериального давления у больных с высоким давлением является довольно скромным — в пределах 10 процентов.
Статиновые препараты, такие как Crestor, Lipitor, и Zocor, действуют путем ингибирования фермента, который необходим печени для выработки холестерина. К сожалению, они также блокируют выработку других веществ, необходимых для функционирования организма, в том числе CoQ10. Это может объяснить наиболее часто встречающиеся побочные эффекты данных препаратов, особенно усталость и мышечные боли. В одном крупном исследовании ENDOTACT, итоги которого были опубликованы в International Journal of Cardiology в 2005 году, было продемонстрировано, что статиновая терапия значительно снижает уровень CoQ10 в плазме крови, но это снижение можно предотвратить с помощью приема добавки со 150 мг CoQ10. Кроме того, добавки с CoQ10 значительно улучшают функцию выстилки кровеносных сосудов, что является одной из ключевых целей при лечении и профилактике атеросклероза.
В двойных слепых исследованиях было продемонстрировано, что прием добавок с CoQ10 был весьма полезным для некоторых пациентов с болезнью Паркинсона. У всех пациентов в этих исследованиях были три основных симптома болезни Паркинсона — тремор, ригидность и замедление движения, и диагноз заболевания был поставлен им в течение последних пяти лет.
В 2005 году исследование, опубликованное в Archives of Neurology, также показало замедление снижения функциональности у пациентов с болезнью Паркинсона, которые принимали CoQ10. После первоначальной проверки и исходных анализов крови пациенты были разделены случайным образом на четыре группы. Три группы получали CoQ10 в разных дозах (300 мг, 600 мг и 1200 мг в день) в течение 16 месяцев, в то время как четвертая группа получала плацебо. Группа, которая принимала дозу 1200 мг, показала меньшее ухудшение психических и двигательные функций и способности осуществлять повседневные действия, такие как кормление или одевание себя. Наибольший эффект был отмечен в повседневной жизни. У групп, которые получали 300 мг и 600 мг в день, инвалидность развилась в меньшей степени, чем в группе плацебо, но и результаты у членов данных групп были менее кардинальными, чем у тех, кто получал самую высокую дозировку препарата. Эти результаты показывают, что благотворного влияния CoQ10 при болезни Паркинсона можно достичь при наиболее высоких дозах приема препарата. Ни у кого из пациентов не были отмечены какие-либо существенные побочные эффекты.
Кофермент Q10 является очень безопасным. Никогда не сообщалось о серьезных побочных эффектах даже при его длительном применении. Поскольку безопасность во время беременности и лактации не была подтверждена, CoQ10 не следует использоваться в эти периоды, если только врач не установит, что клинические результаты перевешивают риски. Я обычно рекомендую принимать от 100 до 200 мг CoQ10 в день. Для наилучшего усваивания мягких желатиновых капсул их следует принимать с едой. При более высоких уровнях дозировки лучше принимать препарат разделенными дозами, а не одной дозой (лучше по 200 мг три раза в день, чем сразу все 600 мг).
Митохондрия имеет форму цилиндра d = 0,2 - 0,1 мкм и длиной до 7 мкм. Она образована двумя мембранами - наружной и внутренней. Внутренняя митохондриальная мембрана образует выросты - кристы, на которых находятся элементарные (грибовидные) частицы - оксисомы или F 1 частицы, на которых происходят процессы окислительного фосфорилирования.
Между мембранами митохондрий находится межмембранное пространство, содержащее небольшое количество ферментов: ферменты дыхательной цепи и сукцинатдегидрогеназу (СДГ), АТФ-синтетазы.
рис. 1. Электронограмма митохондрии.

Между кристами располагается митохондриальный матрикс - гомогенное мелкозернистое вещество, содержащее несколько сотен окислительно-восстановительных ферментов (ферменты цикла Кребса (кроме СДГ), ферменты окисления жирных кислот, белкового синтеза).
В матриксе содержится собственный геном - кольцевая митохондриальная ДНК (что может свидетельствовать о происхождении этих органелл от аэробных симбионтов), митохондриальные рибосомы и м-, т- и р-РНК. Эти компоненты составляют автономную систему синтеза белка митохондрии. Синтезируемые белки используются для роста самих митохондрий. Их деление происходит путем перешнуровки существующих митохондрий; жизненный цикл митохондрий составляет около 10 суток. Однако, несмотря на автономию, большинство белков митохондрии кодируется ядерной ДНК и лишь 5-6% митохондриальных белков кодируется собственной ДНК.
На электронограмме в матриксе митохондрии видны электроноплотные осмиофильные гранулы, представляющие собой скопления липидов, витаминов, катехоламинов. Mg 2+ , Са 2+ .
Существуют два вида митохондрий:
1. С пластинчатыми кристами.
2. С трубчато-везикулярными кристами.
Последние характерны для клеток, продуцирующих стероидные гормоны (клетки коркового вещества надпочечников, фолликулярные клетки и клетки желтого тела яичника, клетки Лейдига семенника).
В цитоплазме клеток митохондрии скапливаются в местах с максимальным потреблением энергии: около ядра, между миофибриллами, вблизи органелл движения.
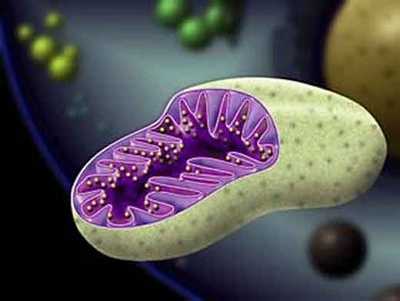
Определение и роль
Митохондрии представляют собой двумембранный органоид эукариотической клетки, основное задание которого – окисление органических соединений, синтез молекул АТФ, с последующим применением энергии, образованной после их распада. То есть по сути митохондрии это энергетическая база клеток, говоря образным языком, именно митохондрии являются своего рода станциями, которые вырабатывают необходимую для клеток энергию.
Количество митохондрий в клетках может меняться от нескольких штук, до тысяч единиц. И больше их естественно именно в тех клетках, где интенсивно идут процессы синтеза молекул АТФ.
Сами митохондрии также имеют разную форму и размеры, среди них встречаются округлые, вытянутые, спиральные и чашевидные представители. Чаще всего их форма округлая и вытянутая, с диаметром от одного микрометра и до 10 микрометров длинны.
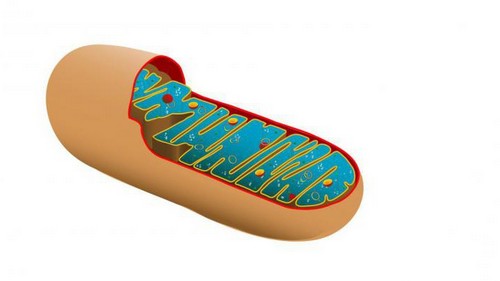
Примерно так выглядит митохондрия.
Также митохондрии могут, как перемещаться по клетке (делают они это благодаря току цитоплазмы), так и неподвижно оставаться на месте. Перемещаются они всегда в те места, где наиболее требуется выработка энергии.
Происхождение
Еще в начале прошлого ХХ века была сформирована так званая гипотеза симбиогенеза, согласно которой митохондрии произошли от аэробных бактерий, внедренных в другую прокариотическую клетку. Бактерии эти стали снабжать клетку молекулами АТФ взамен получая необходимые им питательные вещества. И в процессе эволюции они постепенно потеряли свою автономность, передав часть своей генетической информации в ядро клетки, превратившись в клеточную органеллу.
Строение
Митохондрии состоят из:
- двух мембран, одна из них внутренняя, другая внешняя,
- межмембранного пространства,
- матрикса – внутреннего содержимого митохондрии,
- криста – это часть мембраны, которая выросла в матриксе,
- белок синтезирующей системы: ДНК, рибосом, РНК,
- других белков и их комплексов, среди которых большое число всевозможных ферментов,
- других молекул
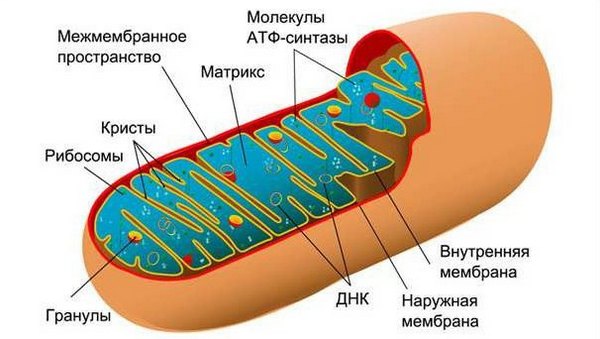
Так выглядит строение митохондрии.
Внешняя и внутренняя мембраны митохондрии имеют разные функции, и по этой причине различается их состав. Внешняя мембрана своим строением схожа с мембраной плазменной, которая окружает саму клетку и выполняет в основном защитную барьерную роль. Тем не менее, мелкие молекулы могут проникать через нее, а вот проникновение молекул покрупнее уже избирательно.
На внутренней мембране митохондрии, в том числе на ее выростах – кристах, располагаются ферменты, образуя мультиферментативные системы. По химическому составу тут преобладают белки. Количество крист зависит от интенсивности синтезирующих процессов, к примеру, в митохондриях клеток мышц их очень много.
У митохондрий, как впрочем, и у хлоропластов, имеется своя белоксинтезирующая система – ДНК, РНК и рибосомы. Генетический аппарат имеет вид кольцевой молекулы – нуклеотида, точь в точь как у бактерий. Часть необходимых белков митохондрии синтезируют сами, а часть получают извне, из цитоплазмы, поскольку эти белки кодируются ядерными генами.
Функции
Как мы уже написали выше, основная функция митохондрий – снабжение клетки энергией, которая путем многочисленных ферментативных реакций извлекается из органических соединений. Некоторые подобные реакции идут с участием кислорода, а после других выделяется углекислый газ. И реакции эти происходят, как внутри самой митохондрии, то есть в ее матриксе, так и на кристах.
Ферменты
Ферменты транслоказы внутренней мембраны митохондрий осуществляют транспортировку АДФ в АТФ. На головках, что состоят из ферментов АТФазы идет синтез АТФ. АТФаза обеспечивает сопряжение фосфорилирования АДФ с реакциями дыхательной цепи. В матриксе находится большая часть ферментов цикла Кребса и окисления жирных кислот
Биологическая роль, структура и выделение митохондрий из печени крыс.
1.1. История вопроса
1.2. Перспективы исследований
2. Биологическая роль митохондрий
3. Ультраструктура митохондрий
3.1. Общие принципы организации
3.2. Особенности строения мембраны митохондрий
4. Описание общих принципов различных методов выделения митохондрий
4.1. Гомогенизация материала
4.1.2. На основе кавитации газов
4.2. Разделение субклеточных компонентов
4.2.2. Разделение в двухфазной системе, содержащей два полимера
4.2.4. Ферменты - маркеры митохондрий
7. Список литературы
Кёлликер одним из первых описал характерным образом ориентированные гранулы в саркоплазме поперечно-полосатой мышцы. Ему принадлежит так же честь первого выделения митохондрий из клеточных структур. В 1888 году он выделил эти гранулы из мышцы насекомых и показал, что они обладают мембраной и набухают в воде. Началом новой эры в цитологическом изучении митохондрий явилась работа Бенсли и Хэрра, которые предприняли попытку выделить митохондрий из взвеси разрушенных клеток печени, применив метод дифференциального центрифугирования. Из-за отсутствия подходящей среды для суспендирования и несовершенства метода центрифугирования Бенсли не удалось получить интактные митохондрий, но именно новаторская работа Бенсли определила слияние цитологических исследований митохондрий с исследованиями дыхания.
В настоящее время основная задача изучения митохондрий на молекулярном уровне сводится к выделению ферментативных компонентов окислительных циклов, определения их молекулярной структуры и механизма действия, анализу их участия в полиферментных системах в митохондриях и картированию их локализации на структурах митохондрий. То обстоятельство, что ферменты дыхательной цепи составляют более 25 % белка митохондриальных мембран заставляют рассматривать эти ферменты не только как функциональные, но и как структурные элементы митохондрии.
Биологическая роль митохондрий.
На протяжении ста с лишним лет, т. е. со времени первых работ Кёлликера (1850), наблюдавшего гранулы в саркоплазме поперечнополосатых мышц, велись кропотливые морфологические исследования, которые постепенно подготавливали почву для всестороннего изучения природы митохондрий. Но только в 1949 г. Кеннеди и Ленинджер установили, что в митохондриях протекает цикл окислительного фосфорилирования, т.е. что митохондрий служат местом генерирования энергии. С этого момента начинается новая эра в изучении митохондрий — эра, в которой блистательные открытия следуют одно за другим. В короткий срок были открыты осмотическая, сократительная, регуляторная и генетическая функции митохондрий и найдены многие из тех молекулярных структур, которые служат первичным субстратом 'этих функций. Было показано также, что митохондрий обеспечивают интеграцию многочисленных процессов клеточного обмена. Эти исследования еще не, завершены, но они могут служить примером плодотворности нового подхода к изучению живого, того подхода, который отличает молекулярную биологию.
В развитии молекулярной биологии за последнее время наметился новый этап. До сих пор это были исследования главным образом, на уровне однородных молекул белков и нуклеиновых кислот, исследования, посвященные их структуре, функции и биосинтезу. Теперь же исследователь не довольствуется этим и переходит к изучению специфически организованных надмолекулярных комплексов, каковыми являются клеточные органеллы. Некоторые функции этих органелл также могут быть истолкованы на уровне индивидуальных молекул, например молекул отдельных ферментов. Но главная особенность клеточных органелл — это интеграция ферментативных процессов. Так, благодаря наличию в составе митохондрий различных белков, липидов, нуклеиновых кислот и углеводов, соединению их между собой и упорядоченному размещению в пространстве в виде трехслойных мембран эти образования приобретают свойства, которые исчезают при их расчленении на отдельные молекулы. Свойства эти: векторный характер действия митохондриальных ферментов (в отличие от скалярного, т. е. не зависящего от направления, действия растворимых ферментов), способность к непосредственному преобразованию энергии окисления в осмотическую и механическую энергию, способность к автономному синтезу белков и т. д. Каждое из этих свойств определяется не простой суммой реакций, катализируемых отдельными ферментами, а обусловлено взаимодействием точно ориентированных ферментных и неферментных макромолекул. Само собой разумеется, что глубокое исследование и познание природы клеточных органелл возможно лишь путем расчленения их на отдельные молекулы с последующей реконструкцией.
Общие принципы организации.
Внутренне пространство митохондрии окружено двумя непрерывными системами мембран, каждая из которых представляет собой замкнутый мешок; эти мешки расположены так, что всю митохондрию можно представить себе, как мешок внутри мешка. Просвет внутреннего мешка не сообщается с пространством между двумя мембранами. Наружная мембрана гладкая, а внутренняя образует многочисленные впячивания, которые в самом простом случае имеют форму перегородок, но могут принимать крайне сложные очертания. Палад назвал эти впячивания митохондриальными кристами. Другой характерный компонент структуры митохондрии - это матрикс, который заполняет просвет, окруженный внутренней мембраной. Известно, что он содержит много белка и некоторое количество липидов; по-видимому, он обладает определенной организацией и более или менее жесткой структурой. Наконец, митохондрии, фиксированные осмием часто содержат в матриксе ряд мелких гранул. Число, диаметр и плотность этих внутримитохондриальных гранул изменяются в зависимости от состояния обмена веществ в тканях.
Особенности строения мембраны митохондрии.
Так как наибольшее практическое значение представляют внутренние мембраны митохондрии, содержащие дыхательные ансамбли, имеет смысл более детально познакомиться с ультраструктурой митохондриальной мембраны. При детальном анализе было выявлено, что митохондриальные мембраны содержат 35 -40 % липидов, преимущественно фосфатидов, и 60 - 65 % белка. Небольшие различия, которые иногда наблюдаются обусловлены различными условиями получения при использовании различных физических и химических способов разрушения структуры митохондрии.
Митохондрии печени крысы содержат значительные количества фосфатидилэтаноламина, фосфатидилхолина, инозитфосфатидов, кардиолипина и фосфатидилсерина; содержание плазмалогена и сфингомиелина невелико, иногда они вовсе отсутствуют. Характерное содержание и количественное содержание липидов в митохондриальной мембране, вероятно обусловлены необходимостью поддержания термодинамически устойчивого двойного слоя липидов, образующего остов мембраны, который служит опорой для дыхательных ансамблей. По-видимому, большое значение имеет тот факт, что практически все липиды митохондриальной мембраны экстрагируются смесью хлороформ - метанол. Это указывает на наличие лишь незначительного числа ковалентных связей между липидами и белковыми элементами или даже на полное их отсутствие; этот факт свидетельствует о высокой степени стабилизации липидов и белков мембранных структурах. Крейн показал, что цитохром с соединяется с фосфатидилэтаноламином, образуя устойчивый комплекс. Возможно, что именно такое взаимодействие липид - белок совместно с гидрофобными связями и обеспечивает такую стабилизацию мембранной структуры. Криддл и сотрудники выделили мономерную форму, которую они назвали структурным белком митохондриальной мембраны. При нейтральном рН структурный белок находится в полимерной форме и не растворим в воде. Мономерная форма имеет молекулярный вес около 22000, но тенденция к полимеризации нарушает точность седиментационных и электрофоретических исследований. Структурный белок способен соединяться с чистыми цитохромами а, Ь, и ее образованием растворимых в воде комплексов в молярном отношении 1:1, причем условия этого взаимодействия для каждого случая различны. Предполагается, что в таких комплексах образуются преимущественно гидрофобные связи. Далее, оказалось, что структурный белок соединяется с фосфолипидами. Таким образом, структурный белок способен к взаимодействию с двумя другими основными молекулярными элементами мембраны - с переносчиками электронов и с фосфолипидами. Склонность цитохромов, флавопротеидов и структурного белка к существованию в мономерной и полимерной формах указывает на выраженную тенденцию этих молекул к образованию очень
устойчивых макромолекулярных ансамблей, имеющих пластинчатую структуру.
Так как для будущих исследований наибольший интерес представляет цитохром с, то следует уделить особое внимание именно этому ферменту.
Этот цитохром отличается от остальных тем, что он легко экстрагируется из митохондрий в растворимой форме с помощью кислот и нейтральных растворов солей. Молекулярный вес кристаллического фермента 12000, изоэлектрическая точка при высоком рН; в молекулу входит одна железопорфириновая группа, которая представлена производным протопорфирина и соединена (ковалентно) с двумя цистеиновыми остатками пептидной цепи, посредством двух тиоэфирных связей.
При рН 7,0 атомы железа в положениях 5 и 6, очевидно, координированы с остатками гистидина; при нейтральных значениях рН цитохром с не имеет тенденции реагировать с кислородом. Известно, что
третичная структура цитохрома с резко изменяется, как функция состояния окисления - восстановления.
Цитохром с восстанавливается тиолами, аскорбатом, хинолами, и восстановленными цитохромами b и с1, а восстановленный цитохром с окисляется феррицианидом, некоторыми красителями и цитохромом а.
Было показано, что в водных растворах цитохром с способен к полимеризации; удалось получить его димер и очистить так же тример и тетрамер. Вторичная и третичная структура цитохрома с изучается методом
рентгеноструктурного анализа. Цитохром с легко соединяется с липидами, в частности с фосфатидилэтаноламином он образует комплекс, названный липоцитохромом с.
Описание общих принципов различных методов выделения митохондрий.
Митохондрии и после выделения сохраняют свой вид, не смотря на то, что мембраны их несколько повреждаются и контуры сглаживаются. В сущности, в наше время их выделяют тем же самым способом, которым пользовался еще Варбург. Прежде всего ткани гомогенизируют, используя для этого изотонический или гипертонический раствор сахарозы (сахароза способствует сохранности митохондрий, стабилизируя митохондриальную мембрану). Затем методом дифференциального центрифугирования отделяют ядра и неразрушенные клетки, а полученную надосадочную жидкость вновь центрифугируют при более высоких скоростях. Митохондрий оседают при этом на дно, образуя плотный буроватый осадок. Промыв этот осадок несколько раз можно получить довольно чистую митохондриальную фракцию. Таким путем очищенные фракции митохондрий были выделены из самых различных клеток животного и растительного происхождения и даже из некоторых простейших.
В более ранних исследованиях митохондрий, как правило, получали из печени животных, так как её клетки очень легко разрушить, а так же потому, что она богата митохондриальной фракцией. После того, как было обнаружено, что бактериальная протеиназа разрушает клетку, не повреждая митохондрий, был разработан простой и быстрый метод получения мышечных митохондрий. Сахароза способствует сохранности митохондрий, стабилизируя митохондриальную мембрану.
Гомогенизация материала. Методы гомогенизации:
(гомогенизатор Даунса 5-12 мл + плотно прилегающий пестик). Гипотонический шок + гомогенизатор.
• Метод основанный на кавитации газов
( клеточная суспензия обогащенная газообразным азотом на 15-30 минут выдерживается при давлении до 65 атм., при быстром понижении давления до атмосферного, вследствие выделения азота происходит разрушение клеток. Органеллы остаются интактными).
Состав среды в которой разрушают клетки определяется применяемым методом гомогенизации. Для разрушения тканей используют изоосмотический раствор сахарозы из расчета 100 мл на 10 г сырого веса ткани, при этом нестрашно если раствора сахарозы будет немного больше, особенно при центрифугировании в роторе с большим объемом. Гипотоническая среда способствует разрушению клеток, но, если обработка длится слишком долго, при этом может происходить разрушение клеточных органелл. Остальные параметры среды трудно обобщить и каждый мембранный препарат требует индивидуального подхода.
Разделение субклеточных компонентов.
Основан на различиях по скорости седиментации (определяется весом, размером и формой частиц). Вещество, используемое для приготовления градиента должно быть легко растворимо в воде, физиологически безвредно и химически инертно; его раствор должен быть невязким, прозрачным в видимой и УФ-областях спектра, должен создавать низкое осмотическое давление. Обычно разделение субклеточных компонентов проводят в градиенте плотности сахарозы. Он удовлетворяет большинству указанных требований, но при высоких концентрациях он весьма вязок. Как правило разделение осуществляется за 3-4 часа или в течение ночи при 80.000 д или выше в роторе с подвесными стаканчиками.
Обработка гомогената из печени крысы 10 мМ фосфатом калия увеличивает скорость седиментации митохондрий, не влияя на осаждение лизосом, что позволяет очищать последние в градиенте плотности перколла.
• Разделение в двухфазной системе, содержащей два полимера.
Полезным и быстрым способом выделения мембран является разделение их в водной двухфазной системе декстран - полиэтиленгликоль (ПЭГ), особенно если отсутствуют необходимые ультрацентрифуги и роторы. Метод, продуктивность которого может быть легко повышена, основан на использовании целого ряда свойств мембран, включая электрический заряд, плотность, массу и гидрофобность. Различие в этих свойствах обуславливает разное распределение компонентов смеси между верхней и нижней фазами и поверхностью раздела. По сродству к верхней фазе компоненты животной клетки располагаются в следующем порядке: эндоплазматический ретикулум / митохондрий / лизосомы / аппарат Гольджи / плазматические мембраны Успех в применении фазовых систем зависит от адекватности выбора их состава. Качество разделения зависит не только от конкретного полимера (пока число их ограничено - в основном используют декстран и ПЭГ), но и от других параметров: молекулярной массы полимера, концентрации солей, рН, химической модификации полимера.
С помощью высоковольтного электрофореза в свободном потоке можно получить препараты субклеточных мембран и органелл высокой степени чистоты. Подлежащую разделению смесь компонентов подают тонкой струйкой в разделяющий буфер, который движется в электрическом поле; мембраны, несущие разный электрический заряд перемещаются в разделяющей ячейке со скоростью, определяемой их зарядом, при этом достигается 100 %-ное разрешение. Это мягкий способ выделения мембран с высоким препаративным выходом. Методику электрофореза в свободном потоке впервые применили для выделения лизосом и митохондрий из печени крысы.
Читайте также: