
Реферат механизм репликации концов линейных хромосом с участием теломеразы
Обновлено: 04.07.2024
• Для стабилизации концов хромосомы необходимы теломеры
• Теломера состоит из простых повторов, Ц+А-обогащенная цепь которых характеризуется последовательностью
При идентификации теломерной последовательности необходимо принимать во внимание два критерия: • Последовательность должна быть расположена на конце хромосомы.
• Она должна придавать линейной молекуле ДНК устойчивость.
Методический подход, который позволил отбирать последовательности ДНК по функциональному критерию, был разработан опять-таки с использованием дрожжей. Все плазмиды, которые способны существовать в клетках дрожжей (и обладающие ARS- и CEN-элементами), представляют молекулы циркулярной ДНК. Линейные плазмиды нестабильны и деградируют.
Теломеры можно отбирать как последовательности, способные обеспечивать стабильность плазмид. Так можно идентифицировать фрагменты дрожжевой ДНК, локализованные на концах хромосом. Аналогичным образом, стабильность линейных форм дрожжевых плазмид обеспечивается концевыми участками природных линейных молекул ДНК — экстрахромосомной рДНК Tetrahymena.
Типичная теломера характеризуется простой повторяющейся структурой,
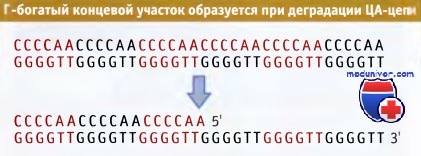
ГТ-богатая цепь которой выходит за пределы ЦА-богатой цепи.
Г-богатый концевой участок образуется при ограниченной деградации ЦА-богатой цепи.
Теломерные последовательности были обнаружены в хромосомах самых разных представителей высших и низших эукариот. Для хромосом клеток растений и человека обнаружен одинаковый тип последовательности. Это свидетельствует о том, что в основе структурной организации теломеры лежит общий принцип. Каждая теломера состоит из протяженного участка коротких последовательностей, расположенных тандемно. В зависимости от организма он может содержать 100-1000 повторов.
Все теломерные последовательности могут быть представлены в виде общей формулы Сn(А/Т)m, где n > 1 и m = 1 - 4. На рисунке ниже представлен типичный пример.
Необычным свойством теломерной последовательности является продолжение ГТ-цепи в виде одиночной, длиной 14—16 оснований. По-видимому, Г-конец образуется за счет специфической ограниченной деградации ЦА-богатой цепи.
Теломерный участок реплицируется по специфическому механизму. Фермент теломераза представляет собой рибонуклеопротеид, который содержит матричную РНК, с той же последовательностью, что и в ЦА-богатой цепи. РНК присоединяется к теломере, образуя праймер, с которого начинается рост цепи с участием обратной транскриптазы, входящей в теломеразный комплекс. Процессивность фермента и число добавляемых повторов контролируются другими белками комплекса.
Поскольку репликация ДНК не может начаться с самого конца линейной молекулы, при репликации хромосомы количество повторов в теломере снижается. Это можно продемонстрировать, заблокировав активность теломеразы. Как следует из рисунка ниже, мутация по гену теломеразы приводит к укорачиванию теломерных участков с каждым делением клетки. Рисунок ниже показывает, что длина теломеры на протяжении 120 клеточных поколений снижается с 400 пн до нуля.
Способность теломеразы добавлять повторы к теломере при синтезе de novo, компенсирует их потерю за счет невозможности протекания репликации до конца хромосомы. Удлинение и укорачивание теломер находятся в динамическом равновесии. Если эти участки постоянно удлиняются (и укорачиваются), то их первичная последовательность не играет роли. Необходимо всего лишь, чтобы конец последовательности признавался подходящим субстратом для удлинения теломеры.
Теломеразная активность присутствует во всех делящихся клетках, и фермент обычно не обнаруживается в терминально дифференцированных, т. е. неделящихся клетках.
Мутации в теломеразе вызывают укорочение теломер после каждого цикла деления.
Полная потеря теломеры вызывает разрывы и перегруппировки хромосом. В диком штамме дрожжей длина теломеры составляет около 350 пн,
однако у мутанта по гену trt1, кодирующего компонент РНК теломеразы, длина ее быстро сокращается до нуля.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Изучение тонкой структуры теломер и механизма действия теломераз находится еще только в начальной стадии. Однако они привлекают к себе огромный интерес исследователей, работающих в самых разных областях биологии и медицины, и здесь уже в ближайшее время можно ждать новых интересных открытий.
Оглавление
Файлы: 1 файл
генетика Баяшов Ердос.pptx
Карагандинский Государственный Медицинский Университет.
Кафедра: Молекулярной биологии
Подготовил: Баяшов Е.Н.
Караганда 2011г.
- Изучение тонкой структуры теломер и механизма действия теломераз находится еще только в начальной стадии. Однако они привлекают к себе огромный интерес исследователей, работающих в самых разных областях биологии и медицины, и здесь уже в ближайшее время можно ждать новых интересных открытий.
- Существование структур на концах хромосом было открыто в 1938 году классиками генетики, лауреатами Нобелевской премии Барбарой Мак-Клинток и Германом Мёллером. Независимо друг от друга они обнаружили, что фрагментация хромосом (под действием рентгеновского облучения) и появление у них дополнительных концов ведут к хромосомным перестройкам и деградации хромосом.
- В сохранности оставались лишь области хромосом, прилегающие к их естественным концам. Лишенные концевых теломер, хромосомы начинают сливаться с большой частотой, что ведет к тяжелым генетическим аномалиям. Следовательно, заключили они, естественные концы линейных хромосом защищены специальными структурами.
- В 1932 Г. Мёллер предложил называть их теломерами (от греч. телос - конец и мерос - часть).
- Хромосома имеет две теломеры.
- У человека теломеры содержат единственный повтор GGGTTA.
- В клетках человека теломеры обычно представлены одноцепочечной ДНК и состоят из несколько тысяч повторяющихся единиц последовательности ТТАГГГ. Эти последовательности с высоким содержанием гуанина стабилизируют концы хромосом, формируя очень необычные структуры, называемые
- G-квадруплексами и состоящие из четырёх, а не двух взаимодействующих оснований. Четыре гуаниновых основания, все атомы которых находятся в одной плоскости, образуют пластинку, стабилизированную водородными связями между основаниями и хелатированием в центре неё иона металла (чаще всего калия). Эти пластинки располагаются стопкой друг над другом.
- Ответственны за прикрепление хромосом к ядерным матриксом;
- Основная функция этих участков — поддержание целостности концов хромосом;
- Играют важную роль в создании специфической архитектуры и внутренней упорядоченности клеточного ядра;
- Теломеры также защищают концы ДНК от деградации экзонуклеазами и предотвращают активацию системы репарации.
- Теломера содержит специальные последовательности ДНК, обеспечивающие точную репликацию хромосом;
- Первым на проблему "концевой недорепликации ДНК" обратил внимание А.М. Оловников в 1971 году.
- Он высказал гипотезу о том, что потеря концевых последовательностей ДНК вследствие их недорепликации ведет к старению клетки.
- Иными словами, предполагалось, что процесс укорочения теломер и есть тот часовой механизм, который определяет репликативный потенциал "смертной" клетки, и когда длина теломер становится угрожающе короткой, этот механизм предотвращает дальнейшее деление клетки.
- А.М. Оловников предположил также, что в нестареющих клетках (а к ним кроме раковых относятся зародышевые, стволовые и другие генеративные клетки) должна существовать специализированная ферментативная система, которая контролирует и поддерживает длину теломерной ДНК.
- Во-первых, было установлено, что теломеры нормальных (то есть обреченных на старение) клеток действительно укорачиваются на 50-60 нуклеотидных звеньев при каждом клеточном делении.
- Во-вторых, в 1984 году Э. Блэкберн и Э. Грайдер выделили фермент, который с помощью механизма, отличного от механизма реакций, лежащих в основе репликации ДНК, синтезирует теломерную ДНК. Этот фермент был назван теломеразой
- Повторное копирование матрицы, включающее этап элонгации, когда дезоксирибонуклеотиды последовательно добавляются к 3'-концу G-богатой цепи теломеры.
- и этап транслокации фермента на конец новообразованной цепи.
- В результате действия теломеразы образуется достаточно длинный 3'-конец, по которому затем достраивается комплементарная цепь. В итоге теломера становится длиннее.
- Теломераза добавляет особые повторяющиеся последовательности ДНК к 3'-концу цепи ДНК на участках теломер.
- Длина теломерных участков хромосом увеличивается или сохраняется на постоянном уровне, компенсируя таким образом концевую недорепликацию и позволяя клетке делиться неограниченно долго.
- РНК-компонент экспрессируется на постоянном уровне практически во всех клетках, и для индуцирования теломеразной активности необходима экспрессия белкового компонента, названного поэтому каталитическим компонентом теломеразы.
- Искусственно индуцированная экспрессия гена каталитического компонента теломеразы, делает клеточную культуру иммортальной (бессмертной), т.е. способной делиться неограниченно долго, отменяя тем самым для культуры предел Хейфлика.
- Теломераза экспрессируется в стволовых, половых и некоторых других типах клеток организма, которым необходимо постоянно делиться для функционирования определённых тканей.
- Клетки 85 % раковых опухолей обладают теломеразной активностью.
- В подавляющем большинстве соматических клеток человека на стадии раннего эмбриогенеза происходит выключение гена ее каталитической субъединицы (обратной транскриптазы),кодирующего теломеразу.
- Тем самым инициируется процесс прогрессивного укорочения теломер, или так называемого "репликативного" старения.
- Другие же составляющие теломеразы, включая теломеразную РНК, образуются в этих клетках, хотя и в меньших количествах, чем в их "бессмертных" прародителях, но постоянно, конститутивно .
- Клетки большинства раковых опухолей характеризуются достаточно высокой активностью теломеразы, которая поддерживает длину теломер на постоянном уровне.
- Этот уровень заметно ниже, чем, например, у эмбриональных клеток, но он достаточен, чтобы обеспечить безграничное деление раковых клеток в культуре.
- Сравнительно небольшая длина теломер у большинства раковых клеток наводит на мысль о том, что они происходят из нормальных клеток, достигших предкризисного состояния.
- Это состояние характеризуется нарушением регуляции многих биохимических реакций. В таких клетках происходят многочисленные хромосомные перестройки, которые в том числе ведут и к злокачественной трансформации.
- Большинство этих клеток погибают, но в части из них в результате случайных мутаций может активироваться постоянная экспрессия генов теломеразы, которая будет поддерживать длину теломер на уровне, необходимом и достаточном для их функционирования.
- Некоторое время вызывал недоумение тот факт, что примерно пятая часть проанализированных раковых опухолей и клеток вообще не содержала активной теломеразы.
- Оказалось, что длина теломер в них поддерживается на должном уровне.
- Таким образом, в этих клетках действует другой, не теломеразный, а скорее рекомбинационный механизм образования теломерной ДНК
Структура теломер. Зелёным цветом показан ион металла, хелатированный в центре структуры
Гипотеза А.М. Оловникова
"КОНЦЕВАЯ НЕДОРЕПЛИКАЦИЯ ДНК"
ДНК-полимеразы, синтезируя дочернюю цепь ДНК, прочитывают родительскую цепь в направлении от ее 3'-конца к 5'-концу. Соответственно дочерняя цепь синтезируется в направлении 5' 3'. В противоположном направлении синтез цепи ДНК фермент катализировать не может.
ДНК-полимераза начинает синтез только со специального РНК-праймера - короткой РНК-затравки, комплементарной ДНК.
После окончания синтеза ДНК РНК-праймеры удаляются, а пропуски в одной из дочерних цепей ДНК заполняются ДНК- полимеразой. Однако на 3'-конце ДНК такой пропуск заполнен быть не может, и поэтому 3'-концевые участки ДНК остаются однотяжевыми, а их 5'-концевые участки - недореплицированными.
Отсюда ясно, что каждый раунд репликации хромосом будет приводить к их укорочению.
Понятно, что прежде всего должна сокращаться длина теломерной ДНК.
Гипотеза А.М. Оловникова нашла убедительное подтверждение в последующие годы.
Теломеразы являются рибонуклеиновыми ферментами. РНК-компонент теломераз содержит короткий район, комплементарный одному повтору G-богатой цепи теломерной ДНК.
Механизм действия теломеразы
Необходимо отметить, что теломераза синтезирует лишь небольшой участок теломеры, утрачиваемый вследствие концевой репликации.
Основная же часть теломерной ДНК реплицируется путем обычного синтеза ведущей и отстающей цепей с помощью ДНК-полимеразы.
Розовым цветом показаны теломеры.
Теломеразная активность
Теломеразная активность соматических клеток
Теломеразная активность раковых клеток
Рак без теломераз?
Стволовые клетки
одна из которых останется стволовой ("бессмертной"),
а другая вступит в процесс дифференцировки
У стволовой клетки всегда есть возможность дать две дочерние клетки,
Понятие и функциональные особенности теломер как районов хромосомы, локализованных на ее конце. Процесс укорочения теломер как часовой механизм, который определяет репликативный потенциал "смертной" клетки. Гипотеза А.М. Оловникова, ее подтверждение.
| Рубрика | Биология и естествознание |
| Вид | реферат |
| Язык | русский |
| Дата добавления | 02.03.2016 |
| Размер файла | 18,7 K |
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
Карагандинский государственный медицинский университет
Кафедра Молекулярной биологии и медицинской генетики
На тему: Теломеры. Теломеразная активность
Выполнила: Рабиева Н.
На концах линейных хромосом эукариот находятся специализированные ДНК-белковые структуры - теломеры. У большинства организмов теломерная ДНК представлена многочисленными короткими повторами. Их синтез осуществляется необычным РНК-содержащим ферментом теломеразой.
ДНК-полимеразы, синтезируя дочернюю цепь ДНК, прочитывают родительскую цепь в направлении от ее 3'-конца к 5'-концу. Соответственно дочерняя цепь синтезируется в направлении 5' 3'. В противоположном направлении синтез цепи ДНК фермент катализировать не может.
Кроме того, ДНК-полимераза начинает синтез только со специального РНК-праймера - короткой РНК-затравки, комплементарной ДНК. После окончания синтеза ДНК РНК-праймеры удаляются, а пропуски в одной из дочерних цепей ДНК заполняются ДНК-полимеразой. Однако на 3'-конце ДНК такой пропуск заполнен быть не может, и поэтому 3'-концевые участки ДНК остаются однотяжевыми, а их 5'-концевые участки - недореплицированными. Отсюда ясно, что каждый раунд репликации хромосом будет приводить к их укорочению. Понятно, что прежде всего должна сокращаться длина теломерной ДНК.
Первым на проблему "концевой недорепликации ДНК" обратил внимание А.М. Оловников в 1971 году. Он высказал гипотезу о том, что потеря концевых последовательностей ДНК вследствие их недорепликации ведет к старению клетки. Иными словами, предполагалось, что процесс укорочения теломер и есть тот часовой механизм, который определяет репликативный потенциал "смертной" клетки, и когда длина теломер становится угрожающе короткой, этот механизм предотвращает дальнейшее деление клетки. А.М. Оловников предположил также, что в нестареющих клетках (а к ним кроме раковых относятся зародышевые, стволовые и другие генеративные клетки) должна существовать специализированная ферментативная система, которая контролирует и поддерживает длину теломерной ДНК.
Гипотеза А.М. Оловникова нашла убедительное подтверждение в последующие годы. Во-первых, было установлено, что теломеры нормальных (то есть обреченных на старение) клеток действительно укорачиваются на 50-60 нуклеотидных звеньев при каждом клеточном делении. Во-вторых, в 1984 году Э. Блэкберн и Э. Грайдер выделили фермент, который с помощью механизма, отличного от механизма реакций, лежащих в основе репликации ДНК, синтезирует теломерную ДНК. Этот фермент был назван теломеразой. теломера хромосома репликативный
Итак, основное назначение теломеразы - синтезировать тандемно повторяющиеся сегменты ДНК, из которых состоит G-цепь теломерной ДНК. Таким образом, она относится к классу ДНК-полимераз, причем оказалось, что теломераза - это РНК-зависимая ДНК-полимераза или обратная транскриптаза. Ферменты этого класса, синтезирующие ДНК на РНК-матрицах, очень хорошо известны молекулярным биологам. Они закодированы и содержатся в ретровирусах (например, в вирусе иммунодефицита человека, вызывающем заболевание СПИДом) и служат для синтеза ДНК-копий их геномов, который в ретровирусе представлен РНК. В клеточном геноме обратные транскриптазы закодированы в ретротранспозонах.
РНК, используемая теломеразой для синтеза теломерной ДНК в качестве матрицы, входит в состав этого фермента. В этом уникальность теломеразы: на сегодня это единственная известная РНК-содержащая обратная транскриптаза. Теломеразные РНК у разных организмов сильно различаются по длине и структуре. Теломеразы простейших содержат РНК длиной в 150-200 нуклеотидных остатков (н.о.), длина теломеразной РНК человека - 450 (н.о.), в то время как теломераза дрожжей содержит аномально длинную РНК (около 1300 н.о.). Как и любая другая РНК клетки, теломеразная РНК обладает специфической вторичной и третичной структурой. Вторичная структура изолированной теломеразной РНК достоверно установлена только для теломераз простейших. Пространственная структура теломеразной РНК в составе ферментативного комплекса пока еще неизвестна.
Матричный участок представлен в теломеразной РНК только один раз. Его длина не превышает длину двух повторов в теломерной ДНК, которые он кодирует и которым он, разумеется, комплементарен.
Так как теломераза синтезирует сегменты ДНК, повторяющиеся много раз, используя только один сегмент своей РНК, она должна обладать способностью периодически (после завершения синтеза каждого повтора) перемещать (транслоцировать) матричный участок в район 3'-конца синтезируемой теломерной ДНК. Источником энергии для такого перемещения, по-видимому, служит сама реакция синтеза цепи теломерной ДНК, поскольку дезоксинуклеозидтрифосфаты - субстраты этой реакции - высокоэнергетические вещества.
На первой стадии теломераза находит 3'-конец теломерной ДНК, с которым часть матричного участка теломеразной РНК образует комплементарный комплекс. При этом теломераза использует 3'-конец хромосомной ДНК в качестве праймера. Далее наступает очередь РНК-зависимой ДНК-полимеразной активности теломеразы. Она обеспечивается специальной субъединицей теломеразы, которая по устройству своего каталитического центра во многом сходна с обратными транскриптазами ретровирусов и ретротранспозонов. Когда синтез ДНК-повтора заканчивается, происходит транслокация, то есть перемещение матрицы и белковых субъединиц фермента на заново синтезированный конец теломерной ДНК, и весь цикл повторяется вновь.
Знакомство даже с весьма схематичным описанием механизма теломеразной реакции приводит к заключению, что двумя компонентами - обратной транскриптазой и теломеразной РНК - для ее осуществления обойтись нельзя. Нет сомнений в том, что в его составе должны быть субъединица, отвечающая за поиск и связывание 3'-конца хромосомы (и выполняющая таким образом своеобразную якорную функцию); субъединица, ответственная за транслокацию; субъединицы, связывающие продукт реакции (однотяжевую ДНК). В составе теломеразы обычно обнаруживается и белковая субъединица с нуклеазной активностью, которая, по-видимому, отщепляет от 3'-конца теломерной ДНК один за другим несколько нуклеотидов до тех пор, пока на этом конце не окажется последовательность, комплементарная нужному участку матричного сегмента теломеразной РНК. Эти субъединицы теломеразы, выполняющие разнообразные функции в ходе синтеза G-цепи теломерной ДНК. Нужно еще раз подчеркнуть, что полный белковый состав фермента не известен до сих пор ни в одном случае.
Широкое распространение теломераз среди эукариот говорит о том, что механизм синтеза теломерной ДНК, который мы наблюдаем у современных организмов, возник очень давно. Более того, эволюционно-генетический сравнительный анализ нуклеотидных последовательностей генов каталитических субъединиц теломераз и других обратных транскриптаз показывает, что этот механизм мог существовать еще до появления первых эукариотических клеток.
С-цепь теломерной ДНК синтезируется с помощью обычной ДНК-полимеразы. Поэтому 3'-концевой участок G-цепи, на котором, по-видимому, первоначально была РНК-затравка, в конечном итоге остается в однотяжевом состоянии (то есть в принципе он готов к тому, чтобы теломераза нарастила на нем новый повтор).
Высокая теломеразная активность наблюдается в половых клетках человека в течение всей его жизни. Соответственно их теломеры состоят из наибольшего числа ДНК-повторов и содержат все необходимые белки для нормальной пролиферации клеток. Аналогичная ситуация наблюдается и для стволовых клеток. Напомним, что стволовые клетки делятся неограниченно долго. Однако у стволовой клетки всегда есть возможность дать две дочерние клетки, одна из которых останется стволовой ("бессмертной"), а другая вступит в процесс дифференцировки. Благодаря этому стволовые клетки служат постоянным источником разнообразных клеток организма. Например, стволовые клетки костного мозга дают начало гемопоэзу - процессу образования клеток крови, а из базальных клеток эпидермиса происходят разнообразные клетки кожного покрова. Как только потомки половых или стволовых клеток начинают дифференцироваться, активность теломеразы падает и их теломеры начинают укорачиваться. В клетках, дифференцировка которых завершена, активность теломеразы падает до нуля, и, как мы уже отмечали, с каждым клеточным делением они с неизбежностью приближаются к состоянию сенесенса (перестают делиться). Вслед за этим наступает кризис, и большинство клеток погибают. Эта картина характерна для подавляющего большинства известных культур клеток эукариот. Однако и здесь есть редкие, но важные исключения: теломеразная активность обнаруживается в таких "смертных" клетках, как макрофаги и лейкоциты.
Недавно было установлено, что нормальные соматические клетки потому лишены теломеразной активности, что в них полностью подавлена экспрессия гена ее каталитической субъединицы (обратной транскриптазы). Другие же составляющие теломеразы, включая теломеразную РНК, образуются в этих клетках, хотя и в меньших количествах, чем в их "бессмертных" прародителях, но постоянно (или, как говорят, конститутивно). Открытие этого важного факта Дж. Шеем, В. Райтом и их сотрудниками и стало основой для той сенсационной работы по преодолению "лимита Хейфлика". Действительно, все остальное было уже делом техники (хотя и очень непростой).
В нормальные соматические клетки были внесены гены теломеразной обратной транскриптазы с помощью специальных векторов, сконструированных из вирусных ДНК. Уровень экспрессии гена в эукариотической клетке зависит от многих факторов, в том числе от белков - факторов транскрипции, связывающихся со специализированными участками ДНК, расположенными в хромосоме по соседству с этим геном. Геномы вирусов, которым нужно быстро размножиться в клетке-хозяине, несут в себе участки ДНК, способные во много раз усилить экспрессию того или иного гена. Исследователи позаботились о том, чтобы в их конструкциях ген теломеразной обратной транскриптазы человека оказался в окружении именно таких участков вирусной ДНК. Результаты их экспериментов можно суммировать кратко: клетки, в которых теломераза поддерживала длину теломер на уровне, характерном для молодых клеток, продолжали делиться и тогда, когда контрольные клетки (без теломеразы) дряхлели и умирали.
В этой и аналогичной ей работах особенно тщательно контролируется отсутствие в культуре клеток раковых клеток. Известно, что клетки большинства исследованных на сегодня раковых опухолей характеризуются достаточно высокой активностью теломеразы, которая поддерживает длину теломер на постоянном уровне. Этот уровень заметно ниже, чем, например, у эмбриональных клеток, но он достаточен, чтобы обеспечить безграничное деление раковых клеток в культуре. Существует гипотеза, у которой немало сторонников, предполагающая, что потеря теломеразной активности соматическими клетками современных организмов есть благоприобретенное в процессе эволюции свойство, уберегающее их от злокачественного перерождения.
Сравнительно небольшая длина теломер у большинства раковых клеток наводит на мысль о том, что они происходят из нормальных клеток, достигших предкризисного состояния. Как мы уже отмечали, это состояние характеризуется нарушением регуляции многих биохимических реакций. В таких клетках происходят многочисленные хромосомные перестройки, которые в том числе ведут и к злокачественной трансформации. Большинство этих клеток погибают, но в части из них в результате случайных мутаций может активироваться постоянная экспрессия генов теломеразы, которая будет поддерживать длину теломер на уровне, необходимом и достаточном для их функционирования.
Некоторое время вызывал недоумение тот факт, что примерно пятая часть проанализированных раковых опухолей и клеток вообще не содержала активной теломеразы. Оказалось, однако, что длина теломер в них поддерживается на должном уровне. Таким образом, в этих клетках действует другой (не теломеразный, а скорее рекомбинационный) механизм образования теломерной ДНК. Иными словами, такие клетки находятся в том же ряду исключений из правила, что и дрозофила.
Какие же практические выводы следуют из того, что на сегодняшний день удалось узнать о связи между активностью теломеразы, раковым ростом и старением клеток. Казалось бы, они лежат на поверхности: не хочешь стареть - активируй теломеразу; хочешь убить раковую опухоль - убей в ней сначала теломеразу.
Легковесность первого вывода (а именно его подхватили средства массовой информации) очевидна: между культурой клеток и клеточной тканью, а тем более организмом дистанция огромного размера. Еще не пришло время всерьез обсуждать проблему получения трансгенных органов человека для пересадки их больным людям (хотя теоретически это, конечно, возможно). А главное, процесс старения не только организма, но и клетки - это исключительно сложный комплекс изменений во множестве биохимических реакций, и его вряд ли можно повернуть вспять, воздействуя только на какую-то одну из них. В то же время существуют вполне реальные планы активировать теломеразу в клетках кожи, которую пересаживают пациентам с сильными ожогами, и тем самым активировать их рост. Или попытаться тем же путем "омолодить" клетки сетчатки глаза, взяв их у пациента, страдающего помутнением сетчатки (а это широко распространенное заболевание у пожилых людей, ведущее к слепоте), и затем вернуть назад.
Что же касается разработки методов избирательного подавления теломеразной активности в раковых опухолях, то сейчас это важное направление в поиске новых средств борьбы со злокачественными заболеваниями. Пока большинство работ связано с испытанием ингибиторов обратных транскриптаз (каталитических субъединиц теломераз). Опыт борьбы со СПИДом, где пытаются решить аналогичную задачу, говорит о том, что определенные надежды найти такое лекарство есть. Главная трудность заключается в том, что каталитическая субъединица теломеразы - это одна из ДНК-полимераз и искомый ингибитор должен быть направлен именно на теломеразную ДНК-синтезирующую активность. В противном случае он будет токсичен для нормальных клеток.
Более перспективными кажутся недавно появившиеся работы, в которых описано избирательное подавление теломеразной РНК, вызывающее гибель раковых клеток в культуре. В нормальных клетках, как это мы отмечали выше, теломеразная РНК синтезируется, но эти клетки лишены теломеразной активности и, скорее всего, теломеразная РНК им не нужна.
Изучение тонкой структуры теломер и механизма действия теломераз находится еще только в начальной стадии. Однако они привлекают к себе огромный интерес исследователей, работающих в самых разных областях биологии и медицины, и здесь уже в ближайшее время можно ждать новых интересных открытий.
Подобные документы
Понятие и функциональные особенности теломер как районов хромосомы, локализованных на ее конце. История их открытия и исследования, современные достижения в данной области. Теломеразная активность и определение основных факторов, влияющих на нее.
презентация [641,8 K], добавлен 25.03.2016
Строение теломер - концевых участков хромосом, характеризующихся отсутствием способности к соединению с другими хромосомами или их фрагментами и выполняющим защитную функцию. Гипотеза концевой недорепликации ДНК А. Оловникова. Механизм действия теломераз.
презентация [838,8 K], добавлен 04.11.2014
Открытие и исследование теломер - специализированных ДНК-белковых структур, их функции. Связь длины теломерных районов хромосом и активности теломеразы с процессами старения и злокачественного роста. Основное назначение теломеразы, принцип работы.
презентация [649,0 K], добавлен 08.12.2014
Изучение тонкой структуры теломер и механизма действия теломераз. Образование теломерной ДНК. Разработка методов избирательного подавления теломеразной активности в раковых опухолях. Поиск новых средств борьбы со злокачественными заболеваниями.
презентация [741,6 K], добавлен 29.05.2013
Изучение строения теломер; их основная функция: участие в фиксации хромосом к ядерному матриксу. Описание принципа маргинотомии в матричном синтезе полинуклеотидов. Концевая недорепликация ДНК. Анализ теломеразной активности соматических и раковых клеток.
ВВЕДЕНИЕ 2
ГЛАВА 1 ОСНОВНАЯ ЧАСТЬ 3
1.1СЛОЖНОСТЬ ПРОЦЕССА РЕПЛИКАЦИИ КОНЦЕВЫХ УЧАСТКОВ ХРОМОСОМ ЭУКАРИОТ 3
1.2 ХАРАКТЕРИСТИКА СТРОЕНИЯ ТЕЛОМЕР ХРОМОСОМ ЭУКАРИОТ 3
1.3 ФЕРМЕНТ ТЕЛОМЕРАЗА КАК ФАКТОР РАСПОЗНАВАНИЯ НУКЛЕОТИДНЫХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ 4
ГЛАВА 2 СПОСОБЫ РЕГУЛИРОВКИ ДЛИНЫ ТЕЛОМЕР НА УРОВНЕ КЛЕТОК 6
ЗАКЛЮЧЕНИЕ 8
СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ 9
ПРИЛОЖЕНИЕ 10
Теломераза обладает способностью распознавать имеющийся конец повторяющихся последовательностей при каждом делении клеток. Ферментативная часть теломеразы напоминает обратные транскриптазы, данные ферменты участвуют в синтезе ДНК, используя для этого матрицу РНК [рис. 1].
После каждой репликации дочерние цепи оказываются короче материнских на размер первого РНК-праймера, при этом образуются выступающие однонитевые 3’-концы материнских цепей, которые будут узнаваться теломеразой и дальнейшем происходит наращивание материнской цепи с помощью 3’-OH-концы в качестве затравок, а РНК в качестве матрицы. Выступающий 3’-конец загибается петлей назад, просовывает однонитевой конец в спираль теломерной повторяющейся последовательности ДНК и в дальнейшем образуется t-петля [рис. 2].
T-петли обеспечивают строение нормальных концов хромосом с уникальной структурой, которая служит защитой их от действия ферментов, а значит и дальнейшей ферментативной деградации, а так же четко ограничивает их от концов других разорванных молекул ДНК, которые клетки быстро репарируют [5]. В свою очередь, образованные одноцепочечные концы служат матрицами для синтеза дочерних цепей по традиционному репликационному механизму [рис. 3].
1. Альбертс, Б. Молекулярная биология клетки / Б.Альбертс. – Ижевск: Институт компьютерных исследований, 2012. – 584-562 с.
2. Богданов, А.А. Теломеры и теломераза / А.А. Богданов // Соросовский Образовательный Журнал. – 1998. №4. – С. 12 – 18.
3. Бокуть, С.Б. Молекулярная биология / C.Б. Бокуть. – Минск : Вышейшая школа, 2005. – 127-129 с.
4. Докудовская, С.С. Теломераза – необычный РНК-содержащий фермент / С.С. Докудовская // Биохимия. – 1997. №11. – С. 1411-1420.
5. Скулачев, В.П. Старение организма – особая функция, а не результат поломки сложной живой системы / В.П. Скулачев // Биохимия. – 1997. №11. – С. 1394-1398.
6. Фаворова, О.О. Сохранение ДНК в ряду поколений: репликация ДНК / О.О. Фаворова // Соросовский Образовательный Журнал. – 1996. №4. – С. 11 – 17.
7. Хейфлик, Л. Смертность и бессмертие на клеточном уровне / Л.Хейфлик // Биохимия. – 1997. №11. – С. 1380-1383.

Вся информация о строении и функционировании любого организма содержится в закодированном виде в его генетическом материале, основу которого у подавляющего числа организмов составляет ДНК. Роль ДНК заключается в хранении и передаче генетической (наследственной) информации в живых организмах. Чтобы эта информация могла передаваться от одного поколения клеток (и организмов) к другому, необходимо её точное копирование и последующее распределение её копий между потомками. Процесс, с помощью которого создаются копии молекулы ДНК, называется репликацией. Реплика́ция (от лат. replicatio — возобновление) — процесс синтеза дочерней молекулы дезоксирибонуклеиновой кислоты на матрице родительской молекулы ДНК. В ходе последующего деления материнской клетки каждая дочерняя клетка получает по одной копии молекулы ДНК, которая является идентичной ДНК исходной материнской клетки. Этот процесс обеспечивает точную передачу генетической информации из поколения в поколение. Репликацию ДНК осуществляет сложный ферментный комплекс, состоящий из 15—20 различных белков, называемый реплисомой.
Клетки, растущие в культуре, могут делиться только определённое число раз, после чего переходят в стадию старения. Такие клетки характеризуются снижением интенсивности энергообмена, замедлением синтеза РНК и белков, понижением эффективности репарации ДНК и накоплением мутаций. Часто наблюдается разбалансировка клеточной регуляции. Признаками клеточного старения считаются накопление специфического гликолипопротеида липофусцина и активация бета-галактозидазы. Стареющие клетки могут долгое время оставаться жизнеспособными. Нередко после остановки деления и торможения клеточного цикла у них не наступает программируемой клеточной гибели. Обычно они уничтожаются клетками иммунной системы. С возрастом в организме происходит накопление старых клеток, вероятно, вследствие ухудшения выполнения иммунной системой своих функций. Клеточное старение — явление, которое обычно связывают с потерей способности клетки к делению. Этот процесс также называется репликативным старением (снижение функциональной активности клеток по мере увеличения их возраста).
Стареющие клетки могут оказывать влияние как на соседние клетки, так и на весь организм, выделяя определённые сигнальные молекулы. Влияние это разнообразно, изучено недостаточно и, в общем, скорее отрицательно. Похоже, что клеточное старение является одним из механизмов старения организма.
ДНК-полимераза не может начинать цепочку ДНК, для этого существует фермент праймаза, который на матрице ДНК синтезирует РНК-фрагмент (праймер, 10-20 нуклеотидов), от 3'-конца которого начинает работать ДНК-полимераза. Праймер затем удаляется, а это место достраивается ДНК-полимеразой следующего по счету фрагмента Оказаки.
На концах хромосом имеются участки, не несущие наследственной информации – теломеры. Их укорочение не приносит вреда; у человека они рассчитаны примерно на 60 репликаций. Больше 60 раз (число Хейфлика) клетки человека поделиться не могут, поскольку концевая недорепликация начинает затрагивать гены.
Стволовые клетки (в коже, красном костном мозге, семенниках) должны делиться гораздо больше, чем 60 раз. Поэтому в них функционирует фермент теломераза, который после каждой репликации удлиняет теломеры.
Теломераза удлиняет выступающий 3'-конец ДНК, так что он увеличивается до размера фрагмента Оказаки. После этого праймаза синтезирует на нем праймер, и ДНК-полимераза удлиняет недореплицированный 5'-конец ДНК.
Теломеры — концевые участки хромосом. Теломерные участки хромосом характеризуются отсутствием способности к соединению с другими хромосомами или их фрагментами и выполняют защитную функцию. У большинства организмов теломерная ДНК представлена многочисленными короткими повторами. Их синтез осуществляется необычным РНК-содержащим ферментом теломеразой.
В клетках человека теломеры обычно представлены одноцепочечной ДНК и состоят из несколько тысяч повторяющихся единиц последовательности ТТАГГГ. Эти последовательности с высоким содержанием гуанина стабилизируют концы хромосом, формируя очень необычные структуры, называемые
G-квадруплексами и состоящие из четырёх, а не двух взаимодействующих оснований. Четыре гуаниновых основания, все атомы которых находятся в одной плоскости, образуют пластинку, стабилизированную водородными связями между основаниями и хелатированием в центре неё иона металла (чаще всего калия). Эти пластинки располагаются стопкой друг над другом.
Теломерная ДНК имеет определенный состав, а для поддержания ее длины, как правило, используется специальный фермент — теломераза.
Нуклеотидная структура теломерных последовательностей ДНК нам уже известна. Более высокие уровни организации образуются за счет специфических белков. Благодаря этим белкам, теломеры имеют плотную упаковку, т. е. относятся к фракции гетерохроматина. С помощью теломерных белков теломеры крепятся к компонентам ядерного матрикса, в т. ч., возможно, к ядерной ламине.
Функции теломер:
механические: теломеры участвуют в фиксации хромосом к ядерному матриксу. Это важно для правильной ориентации хромосом в ядре, и данное обстоятельство особенно проявляется в мейозе;
стабилизационные: в присутствии теломеразы к местам разрыва присоединяется теломерная ДНК. Это стабилизирует хромосомные фрагменты и позволяет им функционировать;
отсчет количества клеточных делений: отсчитывает количество делений клетки после исчезновения теломеразной активности.
У человека длина теломерной ДНК обычно убывает за год на 15-65 пар нуклеотидов. Значительное сокращение длины теломер (достижение критического уровня, при котором клетка неспособна делиться) запускает процессы клеточного старения и резко снижает способность тканей к регенерации. Такое положение дел типично также для для пациентов группы риска в отношении атеросклероза, гипертонии, сердечно-сосудистых и инфекционных заболеваний, диабета, метаболического синдрома, болезни Альцгеймера и некоторых других патологий. Длина теломер (у человека ее часто определяют в лейкоцитах) может служить полезным маркером общего статуса старения организма, а удлинение теломер может коррелировать, согласно первым клиническим наблюдениям, с процессами клеточного омоложения в организме. В одном из исследований Тима Спектора было показано, что курение, избыточный вес и социо-экономический статус также оказывают влияние на длину теломер и продолжительность жизни.
Результаты другого исследования Спектора показали, что длина теломер у физически активных участников исследования была на 200 п.н. длиннее, чем у малоподвижных. Можно сделать вывод, что для предотвращения старения клеток важно поддерживать организм в тонусе с помощью физических упражнений и ведения здорового образа жизни. Исследования, проводимые компанией THINC , показали, что условия, связанные с психологическими нагрузками, в том числе, стрессы, депрессии, тревоги, могут влиять на сокращение длины теломер. До сих пор до конца не ясно, как именно стресс укорачивает теломеры.
Читайте также: