
Координация рефлексов доминанта реферат
Обновлено: 04.07.2024
А.А. Ухтомский [45] первый обратил внимание на то, что доминанта есть общий принцип работы центральной нервной системы и что она определяет освобождение организма от побочной деятельности во
Нейрофизиологические механизмы образования условного рефлекса связаны с суммацией; в начальной стадии (генерализации) образования условный рефлекс аналогичен по физиологическим механизмам доминанте, но затем становится существенно иным. Доминанта образуется на всех и любых этажах центральной нервной системы (например, спинальная доминанта), и для ее образования не обязательно участие коры головного мозга. Для выработки условного рефлекса необходима кора больших полушарий (у высших животных). Они разные по степени своей сложности, по структурам центральной нервной системы, вовлеченным в процесс, по системам связи и значениям этих связей для поведения [8; 20].
Исходной предпосылкой замыкания временной связи является общефизиологический феномен проторения пути. Он заключается в усилении рефлекторного ответа организма при одновременном или последовательном раздражении одного и того же рецептивного поля. Одно раздражение своими следовыми эффектами облегчает появление ответа на действие
следующего. Явление проторения состоит в постепенном повышении проводимости нервного возбуждения по первично стимулируемому пути и нервного центра. Благодаря явлению проторения нередко далее под-пороговое раздражение постепенно оказывается действенной предпосылкой для возникновения очага повышенной возбудимости. Такой процесс непосредственно связан с явлением суммации возбуждения.
Явление суммации впервые было описано И.М. Сеченовым. Главное, на что обращал внимание И.М. Сеченов, это то, что иррадиация возбуждения в центральной нервной системе происходит в направлении очагов повышенной возбудимости. Явление суммации заключается в возникновении действенного возбуждения в нервных центрах в условиях применения ряда последовательных раздражений, каждое из которых само по себе недостаточно, чтобы вызвать ответ. В павловских лабораториях с явлением суммации встретились при первых сочетаниях индифферентного раздражителя с безусловным, и оно получило название суммационного рефлекса.
Изучение свойств доминанты показало, что нейроны доминантного очага характеризуются обширным генерализованным рецептивным полем и способны отвечать на любые раздражители, не имеющие к ней
Исследования доминантного принципа образования временной связи B.C. Русинов проводил с помощью искусственно создаваемого очага повышенной возбудимости в коре головного мозга. Это достигалось различными методическими приемами: легким локальным давлением на мозговую ткань, поляризацией соответствующего участка коры анодом слабого постоянного тока, раздражением электрическим током, пульсирующим в определенном ритме, локальным влиянием слабого раствора стрихнина и другими. Общим результатом этих разнообразных воздействий было повышение возбудимости соответствующих пунктов коры, синхронизация активности их нервных элементов в определенном ритме и усиление способности к суммации возбуждения. Благодаря этому кортикальные пункты, подлежащие такому влиянию,
становятся функционально доминантными, т. е. очагами устойчивого повышенного возбуждения, с тенденцией сохранять приоритетное положение в рефлекторной деятельности мозга достаточно длительное время и после прекращения воздействия соответствующих факторов.
С помощью методики создания искусственного доминантного очага в работах B.C. Русинова [34] и его школы были красиво и убедительно показаны суть и механизмы доминанты. Так, если с помощью прямой поляризации участка коры создать очаг возбуждения, локализованный, например, в кортикальном пункте (проекционной зоне) какой-либо конечности животного, то движение этой конечности можно вызвать воздействием различных индифферентных раздражителей (световых, звуковых и т. п.). При этом двигательная реакция других конечностей не запускается. Помимо этого, B.C. Русиновым и Р.А. Павлыгиной было показано, что если поляризация осуществляется пульсирующим постоянным током, то корковый очаг возбуждения скрыто усваивает этот ритм. Тогда действие посторонних раздражителей запускает ритмические колебания (синхронизацию) активности элементов (с частотой предварительной поляризации) в самом корковом доминантном очаге, а соответственно запускаются ритмические движения конечности животного. Следует отметить, что сама поляризация моторной коры не вызывала двигательных реакций. Движения возникали лишь на афферентные раздражения (свет, звук, тактильные). Настройка на ритм возникала не сразу, а постепенно, при этом важное значение имеет афферентная импульсация. Доминантный очаг способен усваивать ритм афферентных воздействий, т.е. ритмически действующих стимулов (света, звука и др.). Усвоенный ритм сохраняется доминантным очагом надолго — в течение часов и даже дней [34].
175В доминантном очаге усвоенный ритм во время торможения сохраняется в скрытом (латентном) состоянии и вновь после растормаживания проявляет свое афферентное и эфферентное эффективное воздействие на рефлекторную деятельность мозга. Это обстоятельство позволило B.C. Русинову предположить, что как кратковременное, так и длительное сохранение нервных следов активности служит механизмом формирования, локализации временной связи (эн-граммы).
B.C. Русинов считает, что доминанта образуется как рефлекторная физиологическая система, но ее особенностью является обязательное наличие первичного очага в одном из отделов центральной нервной системы. Без признания первичного, центрального, инициирующего доминантного очага — своего рода пей-смекера, модулятора — в доминирующем рефлексе трудно понять, каковы механизмы центральной перестройки в структуре условнорефлекторной деятельности, на каком уровне центральной нервной системы осуществляется конкуренция мотиваций, из которых только одна должна стать доминирующей.
Хотя доминанта способна отозваться на самый широкий спектр внешних раздражителей, возникающие при этом временные связи не носят случайного характера. Доминанта отзывается преимущественно на биологически близкие стимулы, кроме того, она связана с выполнением поисковой фазы поведения не по принципу хаотических проб и ошибок, а с избирательной выборкой тех поведенческих действий, которые в прошлом приводили к успешным результатам.
сигнала является условием быстрого (после одного-двух сочетаний) обучения (образования условного рефлекса) [32; 34].
Однако следует отметить, что условные рефлексы, замыкающиеся после одного сочетания, не являются какой-то высшей, более совершенной формой обучения по сравнению с рефлексами, требующими многократных повторений для их прочной фиксации. Если бы все условные рефлексы замыкались после одного-двух сочетаний, живые существа оказались бы жертвами случайных обстоятельств. Ведь в огромном большинстве случаев одно-два совпадения стимула с подкреплением вовсе не означают, что такое совпадение будет и в дальнейшем. Именно устойчивость, а также способность к угасанию условных рефлексов при изменившихся ситуациях и обеспечивают живым существам широкие адаптивные возможности.
Механизму обратных связей Э.А. Асратян придавал решающее значение в условнорефлекторной деятельности, в организации поведенческих реакций.
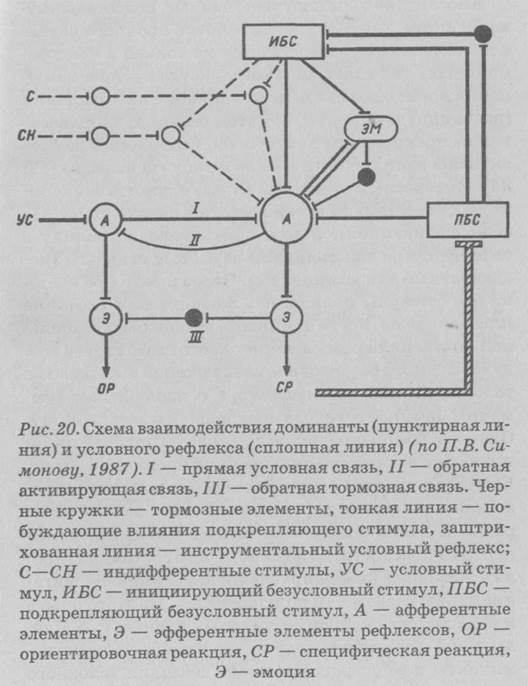
181П.В. Симонов (1987) [36] рассматривает взаимодействие доминанты и условного рефлекса в качестве функциональной единицы индивидуально приобретаемого поведения. Суммируя имеющиеся сведения об анатомических связях, данные о последствиях повреждений тех или иных мозговых образований, результаты регистрации суммарных и клеточных электрических процессов, а также основываясь на собственных экспериментальных данных, П.В. Симонов выдвинул гипотезу о событиях, последовательно и параллельно развивающихся в головном мозге высших млекопитающих в процессе организации поведенческого акта (рис. 21).
Внутренние (метаболизм) и внешние (боль, запах и т. д.) побуждающие безусловные стимулы активируют мотивационные структуры гипоталамуса (ГТ), который в свою очередь активирует гиппокамп (ГИП) и передние отделы новой коры. Благодаря гиппокам-пу широкий круг внешних стимулов усиливает доминантное состояние. Иными словами, гиппокамп участвует в формировании мотивационной доминанты, но выделение доминирующей доминанты связано с деятельностью ядер миндалевидного комплекса. В случае совпадения внешних стимулов с действием подкрепляющих безусловных раздражителей гиппокамп оказывается первым местом встречи сочетаемых афферентаций [36].
При сформированном поведенческом акте в результате совместной деятельности гиппокампа и фронтальной коры (ФК) отбираются те внешние стимулы или их энграммы, которые ранее сопровождались удовлетворением данной потребности. Путем сопоставления мотивационного возбуждения с наличными стимулами и энграммами, извлеченными из памяти (через височную кору), в миндалине (М) формируется эмоциональная окраска (оценка) этих
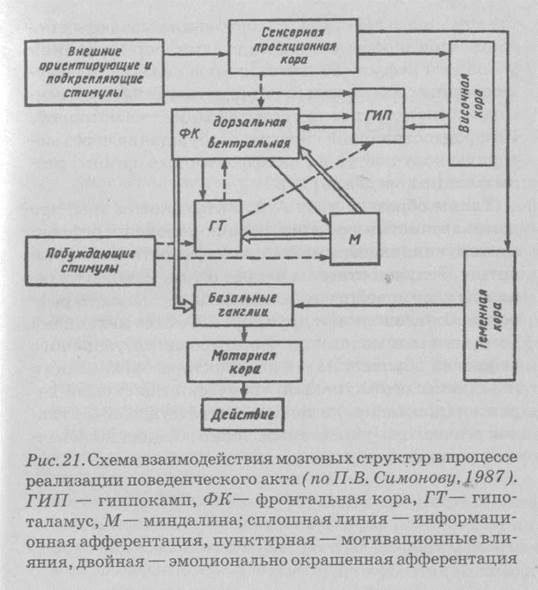
стимулов и энграмм, что ведет к выделению доминирующей мотивации, подлежащей первоочередному удовлетворению. Можно отметить, что нейроны миндалины обезьян с большей избирательностью реагируют на лицо человека или морду другой обезьяны, чем аналогичные нейроны височной коры.
Таким образом, идея А.А. Ухтомского о том, что доминанта есть ключ к пониманию условного рефлекса, получила экспериментальное и теоретическое развитие. Получен ответ на вопрос о том, каковы механизмы взаимодействия доминанты и условного рефлекса. Они дополняют друг друга. Синтез механизма доминанты и механизма формирования условного рефлекса обусловливает два фактора организации поведения: его активный, творчески-поисковый характер (доминанта) и точное соответствие объективной реальности (упроченный, тонко специализированный условный рефлекс).
Рефлекс в переводе с латинского языка означает "отражение". Впервые термин "отражение" или "рефлектирование" был применен Р. Декартом (1595-1650) для характеристики реакций организма в ответ на раздражение органов чувств. Он первым высказал мысль о том, что все проявления эффекторной активности организма вызываются вполне реальными физическими факторами. После Р. Декарта представление о рефлексе было развито чешским исследователем Т. Прохазкой, который развил учение
Содержание
Работа состоит из 1 файл
Контр. раб. по физиологии.doc
Министерство образования Республики Беларусь
Учреждение образования «Гродненский государственный
по предмету
- Принципы координации рефлекторной деятельности:
реципрокности, общего конечного пути, доминанта,
обратные связи
2.Ретикулярная формация. Основные черты строения.
Восходящие и нисходящие влияния
студентка 2 курса
Гаевая Ольга Анатольевна
зоологии и физиологии
человека и животных,
Кирилова Ольга Михайловна
- Принципы координации рефлекторной деятельности: реципрокности, общего конечного пути, доминанта, обратные связи.
Взаимодействие нервных клеток составляет основу целенаправленной деятельности нервной системы и прежде всего осуществления рефлекторных актов.
Основным механизмом деятельности ЦНС является рефлекс. Рефлекс- это ответная реакция организма на действия раздражителя, осуществляемая с участием ЦНС и направленная на достижение полезного результата.
Рефлекс в переводе с латинского языка означает "отражение". Впервые термин "отражение" или "рефлектирование" был применен Р. Декартом (1595-1650) для характеристики реакций организма в ответ на раздражение органов чувств. Он первым высказал мысль о том, что все проявления эффекторной активности организма вызываются вполне реальными физическими факторами. После Р. Декарта представление о рефлексе было развито чешским исследователем Т. Прохазкой, который развил учение
об отражательных действиях. В это время уже было отмечено, что у спинальных животных движения наступают в ответ на раздражение определенных участков кожи, а разрушение спинного мозга ведет к их исчезновению.
Дальнейшее развитие рефлекторной теории связано с именем И. М. Сеченова. В книге "Рефлексы головного мозга" он утверждал, что все акты бессознательной и сознательной жизни по природе происхождения являются рефлексами. Это была гениальная попытка ввести физиологический анализ в психические процессы. Но в то время не существовало методов объективной оценки деятельности мозга, которые могли бы подтвердить это предположение И. М. Сеченова. Такой объективный метод был разработан И. П. Павловым - метод условных рефлексов, с помощью которого он доказал, что высшая нервная деятельность организма, так же как и низшая, является рефлекторной.
Структурной основой рефлекса, его материальным субстратом (морфологической основой) является рефлекторная дуга - (reflex arc) - нервная цепь, участвующая в осуществлении рефлекса; простейшая рефлекторная дуга состоит из двух нейронов: чувствительного и двигательного. Тело первого нейрона (афферентного) находится в спинномозговом узле (или чувствительном узле черепного нерва). Периферические отростки этих клеток направляются в составе соответствующего спинномозгового или черепного нервов на периферию, где заканчиваются рецепторным аппаратом, который воспринимает раздражение. При возникновении простого рефлекса (такого как патсллярный рефлекс) в этом процессе могут принимать участие только два нейрона, тогда как другие рефлексы требуют взаимодействия нескольких вставочных нейронов рефлекторной дуги. Рефлекторная дуга состоит из афферентной, центральной и эфферентных частей, связанных между собой с помощью синоптических соединений. Афферентная часть представлена теми нервными элементами, которые формируют и проводят в центральном направлении нервные импульсы, необходимые для деятельности всей рефлекторной дуги. Центральная часть рефлекса может включать несколько последовательных нейронов, соединенных синаптическими контактами (называются полисимпатические), если рефлекс имеет один синапс (например, синапс между центральными окончаниями мышечных афферентов и мотонейронами), то он носит название моносимпатического (сухожильный рефлекс, или рефлекс растяжения).
В большинстве своем рефлексы служат для защиты организма и приспособления его к изменениям окружающей и внутренней среды. С их помощью адекватно координируются непроизвольные акты организма.
Характер рефлекса в значительной степени зависит от интенсивности раздражения и числа активируемых рецепторов.
В основе современного представления о рефлекторной деятельности лежит понятие полезного приспособительного результата, ради которого совершается любой рефлекс. Информация о достижении полезного приспособительного результата поступает в ЦНС по звену обратной связи в виде обратное афферентации, которая является обязательным компонентом рефлекторной деятельности. Принцип обратной афферентации введен в рефлекторную теорию П. К. Анохиным. Таким образом, по современным представлениям структурной основой рефлекса является не рефлекторная дуга, а рефлекторное кольцо, состоящее из следующих компонентов (звеньев):
• афферентный нервный путь;
• эфферентный нервный путь;
• рабочий орган (эффектор);
Рефлекторная деятельность центральной нервной системы всегда представляет собой сложнейшую систему рефлексов, и даже в искусственно изолированном участке мозга вызвать в изолированной форме только одну рефлекторную реакцию довольно сложно. Рефлекторные процессы неизбежно взаимодействуют друг с другом и изменяют друг друга. Неизбежность такого взаимодействия вытекает, в частности, из того обстоятельства, что различные типы рефлекторных реакций осуществляются очень часто через одни и те же двигательные нейроны, т.е. связаны с использованием одних и тех же мышц. Двигательные нейроны являются, как это определил Ч. Шеррингтон, общим конечным двигательным путем - конечным звеном осуществления ряда двигательных реакций, имеющих часто различное функциональное значение. Так, одни и те же мотонейроны могут принимать участие в защитном рефлексе, в рефлексе статического поддержания позы, в локомоторных движениях (бег, прыжки, ходьба и т. д.).
Основное правило взаимодействия рефлексов заключается в том, что функционально однонаправленные рефлексы усиливают, подкрепляют друг друга, а функционально противоположные, наоборот, взаимно тормозятся. Функциональная направленность при этом определяется характером конечной рефлекторной реакции. Так, например, при нанесении нескольких раздражений, каждое из которых вызывает сгибательный рефлекс, общая реакция будет существенно усиленной; то же самое касается одновременного вызова нескольких разгибательных рефлексов, образующих функционально противоположную группу. Если же на организм одновременно действуют раздражители, вызывающие сгибательную и разгибательную рефлекторные реакции, то обе они окажутся ослабленными вследствие тормозящих влияний соответствующих нервных центров друг на друга.
Взаимное подкрепление функционально однородных рефлексов характеризуется определенными закономерностями, которые особенно удобно наблюдать, если такие рефлексы вызываются раздражением двух близко расположенных рецептивных полей и связаны с рефлекторным возбуждением мотонейронов одних и тех же мышц.
Если раздражение двух рецептивных полей настолько слабо, что каждое из них хотя и возбуждает афферентные волокна, но все же недостаточно для того, чтобы вызвать разряд мотонейронов, то при одновременном их раздражении, тем не менее, может возникнуть отчетливая рефлекторная реакция. Это явление обычно называют суммацией, однако, употребляя такой термин, необходимо четко себе представлять, что физиологическое его понимание отличается от алгебраического - в этом случае при сложении внешне нулевых эффектов создается выраженный ответ. Другим часто применяемым в этом случае термином является облегчение. По-видимому, более правильно говорить об облегчении рефлекса, так как речь идет именно об облегчении возникновения разряда мотонейронов, а не просто о механическом сложении двух процессов.
Облегчение может возникать и при некотором усилении раздражения рецептивных полей - так, чтобы и одно, и другое раздражение вызывало небольшую рефлекторную реакцию. В результате сочетания раздражений рефлекс оказывается значительно большим, чем алгебраическая сумма отдельных реакций. При еще большем усилении стимулов можно получить суммарную рефлекторную реакцию, которая уже действительно будет равна алгебраической сумме двух рефлексов, взятых порознь. И, наконец, при сочетании особенно сильных рефлекторных реакций суммарный результат может оказаться лишь несколько превосходящим одну из этих реакций или равным ей. Для последнего случая применяется термин окклюзии рефлекторных реакций.
Представить себе возникновение приведенных выше различных форм взаимодействия рефлексов проще на схеме, предложенной Ч. Шеррингтоном. Схема изображает двигательное ядро; часть его нейронов дает аксоны в одну мышцу, другая часть - в другую. К мотонейронам подходят афферентные пути от соответствующих рецептивных полей. При поступлении слабой афферентной волны, которая не приводит к рефлекторному ответу, в какой-то части мотонейронов возникают только постсинаптические реакции, которые недостаточны для того, чтобы вызвать распространяющийся импульс. Эта афферентная волна, хотя и эффективна в отношении синаптического действия, недостаточна для создания рефлекторной реакции. То же самое вызывает слабая афферентная волна от другого рецептивного поля. Она также активирует какую-то группу мотонейронов, но вызывает в ней только локальные синаптические процессы.
1. повышенной возбудимстью (порог возбудимости должен быть равен возможному раздражителю)
2. стойкостью возбуждения (возбуждение для формирования новой Д должно длится некоторое время – для формирования голодовой Д у кролика – 3 суток без пищи)
3. способностью к суммированию возбуждения (величина влияния Д на текущую деятельность равна возбуждению Д)
4. способностью к затормаживанию центров, в нее не входящих
Владимирский Н.Д. и Кацнельсон Р.С. – вызывание Д на ганглии брюхоногого моллюска (отравление стрихнином); при возбуждении другого ганглия – ответная реакция, соответ. отравленному. таким образом, Д привлекает к себе возбуждение из самых различных источников.
Роль Д во внимании и предметном мышлении:
1. достаточно устойчивая Д, возникшая под влиянием внутренней секреции, рефлекторной деятельности и т.д., привлекает к себе в качестве поводов для возбуждения самые разнообразные рецепции (первый бал Наташи Ростовой)
2. из множества действующих рецепций Д выбирает для себя наиболее биологически адекватные (стадия адекватного выбора из окружающей среды) (Наташа и Андрей)
3. между Д и данными рецептивного содержания (комплексом раздражителей) формируется устойчивая связь, каждый из контрагентов которой будет вызывать и подкреплять исключительно друг друга.
Инстинкт на гормональной основе (м\б воспроизведен на нервно-рефлекторной основе, как только состояние внешней среды будет соответствующим). Бидль: половое влечение у людей кастратов.
Связь условного рефлекса и доминанты.
В.С. Русинов выделяет несколько стадий образования УР: простой суммационный рефлекс, доминанта и собственной УР. В начальной стадии образования (генерализации) УР = Д, а затем происходит дифференциация. Исследования Русиновым свойств Д – создание искусственного очага постоянного возбуждения. нейроны, входящие в состав Д. им. генерализованное рецептивное поле и способны отвечать на раздражители, которые не имеют прямого отношения к данной Д. например, при возбуждении участка коры ГМ (проекции какой-либо конечности) при предъявлении индиферентных стимулов (световой, звуковой) – ответная реакция конечности. Д способна усваивать ритм афферентных раздражителей (ритмические движения конечности). Возникновение Д возможно только при неоднократной стимуляции соответствующих РД.
В центральной нервной системе под влиянием тех или иных причин может возникнуть очаг повышенной возбудимости, который обладает свойством притягивать к себе возбуждения с других рефлекторных дуг и тем самым усиливать свою активность и тормозить другие нервные центры. Это явление носит название доминанты.
Доминанта относится к числу основных закономерностей в деятельности центральной нервной системы. Она может возникнуть под влиянием различных причин: голода, жажды, инстинкта самосохранения, размножения. Состояние пищевой доминанты хорошо сформулировано в русской пословице: "Голодной куме все хлеб на уме". У человека причиной доминанты может быть увлеченность работой, любовь, родительский инстинкт. Бсли студент занят подготовкой к экзамену или читает увлекательную книгу, то посторонние шумы не мешают ему, а даже углубляют его сосредоточенность, внимание.
Весьма важным фактором координации рефлексов является наличие в центральной нервной системе известной функциональной субординации, т. е. определенного соподчинения между ее отделами, возникающего в процессе длительной эволюции. Нервные центры и рецепторы головы как "авангардной" части тела, прокладывающей путь организму в окружающей среде, развиваются быстрее. Высшие отделы центральной нервной системы приобретают способность изменять активность и направление деятельности нижележащих отделов.
Важно отметить: чем выше уровень животного, тем сильнее власть самых высших отделов центральной нервной системы, "тем в большей степени высший отдел является распорядителем и распределителем деятельности организма" (И. П. Павлов).
У человека таким "распорядителем и распределителем" является кора больших полушарий головного мозга. Нет функций в организме, которые бы не поддавались решающему регулирующему влиянию коры.
Схема 1. Распространение (направление показано стрелками) нервных импульсов по простой рефлекторной дуге (увеличить рисунок)
1 - чувствительный (афферентный) нейрон; 2 - вставочный (кондукторный) нейрон; 3 - двигательный (эфферентный) нейрон; 4 - нервные волокна тонкого и клиновидного пучков; 5 - волокна корково-спинномозгового пути.
Условный рефлекс - это сложная многокомпонентная реакция, которая вырабатывается на базе безусловных рефлексов с использованием предшествующего индифферентного раздражителя. Он имеет сигнальный характер, и организм встречает воздействие безусловного раздражителя подготовленным. Например, в предстартовый период происходит перераспределение крови, усиление дыхания и кровообращения, и когда мышечная нагрузка начинается, организм уже к ней подготовлен.
Правила выработки условных рефлексов
Для выработки условного рефлекса необходимо:
1) наличие двух раздражителей, один из которых безусловный (пища, болевой раздражитель и др.), вызывающий безусловно-рефлекторную реакцию, а другой - условный (сигнальный), сигнализирующий о предстоящем безусловном раздражении (свет, звук, вид пищи и т.д.);
2) многократное сочетание условного и безусловного раздра жителей (хотя возможно образование условного рефлекса при их однократном сочетании);
3) условный раздражитель должен предшествовать действию безусловного;
4) в качестве условного раздражителя может быть использован любой раздражитель внешней или внутренней среды, который должен быть по возможности индифферентным, не вызывать обронительной реакции,не обладать чрезмерной силой и способен привлекать внимание;
5) безусловный раздражитель должен быть достаточно сильным, в противном случае временная связь не сформируется;
6) возбуждение от безусловного раздражителя должно быть более сильным, чем от условного;
7) необходимо устранить посторонние раздражители, так как они могут вызывать торможение условного рефлекса;
8) животное, у которого вырабатывается условный рефлекс, должно быть здоровым;
9) при выработке условного рефлекса должна быть выражена мотивация, например, при выработке пищевого слюноотделительного рефлекса животное должно быть голодным, у сытого - этот рефлекс не вырабатывается.
Условные рефлексы легче вырабатывать на экологически близкие данному животному воздействия. В связи с этим условные рефлексы делятся на натуральные и искусственные. Натуральные условные рефлексы вырабатываются на агенты, которые в естественных условиях действуют вместе с раздражителем, вызывающим безусловный рефлекс (например, вид пищи, ее запах и т.д.). Все остальные условные рефлексы искусственные, т.е. вырабатываются на агенты, в норме не связанные с действием безусловного раздражителя, например, пищевой слюноотделительный рефлекс на звонок.
Физиологической основой для возникновения условных рефлексов служит образование функциональных временных связей в высших отделах ЦНС. Временная связь - это совокупность нейрофизиологических, биохимических и ультраструктурных изменений в мозге, возникающих в процессе совместного действия условного и безусловного раздражителей. И.П.Павлов высказал предположение, что при выработке условного рефлекса происходит формирование временной нервной связи между двумя группами клеток коры - корковыми представительствами условного и безусловного рефлексов. Возбуждение от центра условного рефлекса может передаваться к центру безусловного рефлекса от нейрона к нейрону.
Следовательно, первый путь образования временной связи между корковыми представительствами условного и безусловного рефлексов является внутрикорти-кальным. Однако при разрушении коркового представительства условного рефлекса выработанный условный рефлекс сохраняется. По-видимому, образование временной связи идет между подкорковым центром условного рефлекса и корковым центре-безусловного рефлекса. При разрушении коркового представительства безусловного рефлекса условный рефлекс также сохраняется. Следовательно, выработка временной связи может идти между корковым центром условного рефлекса и подкорковьг центром безусловного рефлекса.
Если вам нужна помощь в написании работы, то рекомендуем обратиться к профессионалам. Более 70 000 авторов готовы помочь вам прямо сейчас. Бесплатные корректировки и доработки. Узнайте стоимость своей работы.
Разобщение корковых центров условного и безусловного рефлексов путем пересечения коры мозга не препятствует образованию условного рефлекса. Это свидетельствует о том, что временная связь может образоватьс между корковым центром условного рефлекса, подкорковьг центром безусловного рефлекса и корковым центром безусловного рефлекса.
Имеются различные мнения по вопросу о механизмах обрс зования временной связи. Возможно, образование временно связи происходит по принципу доминанты. Очаг возбуждения с безусловного раздражителя всегда сильнее, чем от условного, так как безусловный раздражитель всегда биологически более значим для животного. Этот очаг возбуждения является доминантным, следовательно притягивает к себе возбуждение от очага условного раздражения. Если возбуждение прошло по каким-либо нервным цепям, то в следующий раз оно по этим путям пройдет значительно легче (явление "проторения пути"). В основе этого лежат: суммация возбуждений, длительное повышение возбудимости синаптических образований, увеличение количества медиатора в синапсах, увеличение образования новых синапсов. Все это создает структурные предпосылки к облегчению движения возбуждения по определенным нейронным цепям.
Другим представлением о механизме формирования временной связи является конвергентная теория. В ее основе лежит способность нейронов отвечать на раздражения разных модальностей. По представлениям П.К.Анохина, условный и безусловный раздражители вызывают распространенную активацию корковых нейронов благодаря включению ретикулярной формации. В результате восходящие сигналы (условного и безусловного раздражителей) перекрываются, т.е. происходит встреча этих возбуждений на одних и тех же корковых нейронах. В результате конвергенции возбуждений возникают и стабилизируются временные связи между корковыми представительствами условного и безусловного раздражителей.
Условные рефлексы второго, третьего и более высоких порядков.
Если выработать прочный условный пищевой рефлекс, например, на свет, то такой рефлекс является условным рефлексом первого порядка. На его базе можно выработать условный рефлекс второго порядка, для этого дополнительно применяют новый, предшествующий сигнал, например звук, подкрепляя его условным раздражителем первого порядка (светом).
В результате нескольких сочетаний звука и света звуковой раздражитель также начинает вызывать слюноотделение. Таким образом возникает новая более сложная опосредованная временная связь. Следует подчеркнуть, что подкреплением для условного рефлекса второго порядка является именно условный раздражитель первого порядка, а не безусловный раздражитель (пища), так как если и свет и звук подкреплять пищей, то возникнут два отдельных условных рефлекса первого порядка. При достаточно прочном условном рефлексе второго порядка можно выработать условный рефлекс третьего порядка.
Для этого используется новый раздражитель, например, прикосновение к коже. В этом случае прикосновение подкрепляется только условным раздражителем второго порядка (звуком), звук возбуждает зрительный центр, а последний - пищевой центр. Возникает еще более сложная временная связь. Рефлексы более высокого порядка (4, 5, 6 и т.д.) вырабатываются только у приматов и человека.
Исследуя особенности межцентральных отношении, А. А. Ухтомский обнаружил, что если в организме животного осуществляется сложная рефлекторная реакция, например повторяющиеся акты глотания, то электрические раздражения моторных центров коры не только перестают вызывать в этот момент движения конечностей, но и усиливают и ускоряют протекание начавшейся цепной реакции глотания, оказавшейся главенствующей. Аналогичное явление наблюдалось при отравлении фенолом передних отделов спинного мозга лягушки.
Вложенные файлы: 1 файл
Dominanta.doc
Активность нервных центров непостоянна, и преобладание активности одних из них над активностью других вызывает заметные перестройки в процессах координации рефлекторных реакций.
Исследуя особенности межцентральных отношении, А. А. Ухтомский обнаружил, что если в организме животного осуществляется сложная рефлекторная реакция, например повторяющиеся акты глотания, то электрические раздражения моторных центров коры не только перестают вызывать в этот момент движения конечностей, но и усиливают и ускоряют протекание начавшейся цепной реакции глотания, оказавшейся главенствующей. Аналогичное явление наблюдалось при отравлении фенолом передних отделов спинного мозга лягушки. Повышение возбудимости мотонейронов приводило к тому, что отравленная лапка отвечала рефлексом потирания (отряхивания) не только на непосредственное раздражение ее кожи кислотой, но и на самые разнообразные посторонние раздражители:
поднимание животного со стола в воздух, удар по столу, где оно сидит, прикосновение к передней лапке животного и др.
В 1923 г. А. А. Ухтомский сформулировал принцип доминанты как рабочий принцип деятельности нервных центров.
Термином доминанта был обозначен господствующий очаг возбуждения в центральной нервной системе, определяющий текущую деятельность организма.
Основные черты, доминанты следующие: 1) повышенная возбудимость нервных центров, 2) стойкость возбуждения во времени, 3) способность к суммации посторонних раздражении и 4) инерция доминанты. Доминирующий (господствующий) очаг может возникнуть лишь при определенном функциональном состоянии нервных центров. Одним из условий его образования является повышенный уровень возбудимости нервных клеток, который обусловливается различными гуморальными и нервными влияниями (длительными афферентными импульсациями, гормональными перестройками в организме, воздействиями фармакологических веществ, сознательным управлением нервной деятельностью у человека и пр.).
Установившаяся доминанта может быть длительным состоянием, которое определяет поведение организма на тот или иной срок. Способность стойко поддерживать возбуждение во времени — характерная черта доминанты. Однако далеко не всякий очаг возбуждения становится доминантным. Повышение возбудимости нервных клеток и их функционального значения определяется способностью суммировать возбуждение при поступлении любого случайного импульса.
Восходящие нервные импульсы могут направляться не только по прямому специфическому пути — в соответствующие проекционные зоны мозга, но и через боковые ответвления — в любые зоны центральной нервной системы (см. § 6 этой главы). В связи с этим при наличии в каком-либо участке нервной системы очага с оптимальным уровнем возбудимости этот очаг приобретает возможность повышать свою возбудимость за счет суммирования не только собственных афферентных раздражении, но и посторонних, адресованных другим центрам. Не сила возбуждения, а способность накапливать и суммировать его превращает нервный центр в доминирующий. Явления суммации лучше всего выражены лишь при умеренном, оптимальном, повышении возбудимости нейронов. Это выражается в том, что доминанта легче всего подкрепляется слабыми раздражениями и гасится сильными.
Объединение большого числа нейронов в одну рабочую систему происходит путем взаимного сонастраивания на общий темп активности, т. е. путем усвоения ритма. Одни нервные клетки снижают свой более высокий темп деятельности, другие повышают низкий темп до какого-то среднего, оптимального, ритма. Доминирующая группа нервных центров, работающих в общем ритме, тормозит центры с иными ритмами активности. Значение явления усвоения ритма как механизма формирования доминирующего очага и механизма его функционального выделения из общей массы нервных клеток подтверждено в последнее время электрофизиологическими исследованиями на животных и человеке.
Важным свойством доминанты является инерция. Однажды возникшая доминанта может длительное время поддерживаться и после удаления первоначального стимула, например при осуществлении цепных двигательных рефлексов. Инерция выражается также в том, что доминанта может надолго сохраняться как следовое состояние (потенциальная доминанта). При возобновлении прежнего состояния или прежней внешней ситуации доминанта может возникнуть снова. Такое воспроизведение доминанты происходит в организме спортсмена условнорефлекторно в предстартовом состоянии когда в известной мере активизируются все те нервные центры, которые входили в рабочую систему во время предыдущих тренировок. Это проявляется в усилении всего комплекса функций, связанных с мышечной работой: центральных, мышечных, выделительных, сосудистых и др. Мысленное выполнение физических упражнений также воспроизводит (актуализирует) доминирующую систему центров, что обеспечивает тренирующий эффект представления движений и является основой так называемой идеомоторной тренировки.
В норме в нервной системе редко отсутствуют какие-либо доминанты. Бездоминантное состояние—это очень слабое возбуждение, разлитое более или менее равномерно по различным нервным центрам. Сходное состояние возникает у спортсменов в процессе полного расслабления, при аутогенной тренировке. Путем такого расслабления добиваются устранения мощных рабочих доминант и восстановления работоспособности нервных центров.
Как фактор поведения доминанта связана с высшей нервной деятельностью, с психологией человека. Доминанта является физиологической основой акта внимания. Она определяет характер восприятия раздражении из внешней среды, делая его односторонним, но зато более целеустремленным. При наличии доминанты многие влияния внешней среды остаются вне внимания, но зато более интенсивно улавливаются и анализируются те, которые особенно интересуют человека. Доминанта — мощный фактор отбора биологически и социально наиболее значимых раздражении.
Возникновение в коре больших полушарий доминантных состояний наблюдается в начале образования временных связей. Условный рефлекс образуется, когда доминантный очаг возбуждения начинает отвечать не на любое афферентное раздражение, а лишь на специфическое раздражение, ставшее сигнальным.
Поскольку доминанта связана с определенной реакцией, она определяет одностороннее выражение поведения. Чем выраженное доминанта, тем больше она тормозит другие текущие рефлексы. Таким образом, из многих степеней свободы выбирается одна — при наличии в определенных двигательных центрах доминанты работает интенсивно лишь та часть мускулатуры, которая управляется этими центрами, а остальная выключена из сферы деятельности в результате сопряженного торможения. При этом заторможены также многие вегетативные центры. В начальный момент интенсивной мышечной работы почти полностью могут исчезать условные рефлексы: слюноотделительные, мигательные и др. Это обеспечивает Целесообразность движений и экономичность энерготрат. Мощная Двигательная доминанта при статических усилиях за счет сопряженного торможения приводит к задержке дыхания и угнетению деятельности сердечно-сосудистой системы.
По мере формирования двигательного навыка система доминирующих нервных центров совершенствуется. Из нее исключаются все лишние нервные центры, остаются лишь те, которые необходимы и достаточны для осуществления двигательной задачи.
Читайте также: