
Гидротропизм корней растений реферат
Обновлено: 04.07.2024
- Гидротропизм (от Гидро и греч. trópos — поворот, направление) — изгибание некоторых органов растений под влиянием неравномерного распределения влажности в окружающей их среде.
Впервые явления гидротропизма обнаружены Ю. Саксом в 1872 году на корнях, которые дают отклонение в сторону большей влажности. В такой форме гидротропизм обозначается как положительный: в корнях он обусловлен не непосредственным влиянием влажности на область роста, а влиянием на последнюю раздражения от воспринимаемого, одностороннего распределения влажности верхушкой корня. Помимо корней, положительный гидротропизм проявляется и в некоторых других растительных объектах, например, в корневых волосках и пыльцевых трубках.
Связанные понятия
Фототропи́зм (от др.-греч. φῶς — свет и τρόπος — поворот) — изменение направления роста органов растений или положения тела (органов) у животных, в зависимости от направления падающего света.
Тропизмы (от греч. τροπος — рост, направление) — реакция ориентирования клетки, то есть направление роста или движения клеток относительно раздражителя (химического, светового и др.).
На́стии, или насти́ческие движе́ния (от др.-греч. σπαθητός ‘уплотнённый’) — движения дорсовентральных органов растений, которые обусловлены особенностями самого растения и проявляются при воздействии факторов окружающей среды (температура, свет, влажность и др.). В отличие от тропизмов, настии являются более быстрыми и возникают в ответ на ненаправленные, рассеянные в окружающей среде раздражители.
Ростовые корреляции — это зависимость роста и развития одних органов или частей от других.
Световая точка компенсации, или световой компенсационный пункт (СКП) — наименьшая интенсивность света на световой кривой, при которой активность фотосинтеза и дыхания уравновешиваются по газообмену. В этой точке количество фиксируемого растением CO2 в точности соответствует его количеству, выделяемому растением в результате дыхания и фотодыхания, а потребление O2 в точности соответствует его выделению в результате фотосинтеза.
Основна́я гидрофизи́ческая характери́стика (ОГХ, кривая водоудерживания) — в физике почв изотермическая равновесная зависимость между капиллярно-сорбционным (матричным) давлением почвенной влаги и влажностью (обычно объёмной). Форма ОГХ специфична для каждого почвенного образца и характеризует структуру порового пространства почвы, гранулометрический и минералогический состав. Характеризуется гистерезисом, то есть несовпадением форм кривой при увлажнении и иссушении образца. В виду доминирования.
Закон оптимума (в экологии) — любой экологический фактор имеет определённые пределы положительного влияния на живые организмы.
Гигрофи́ты (от др.-греч. ὑγρός — влажный + φυτόν — растение) — растения, обитающие в местах с высокой влажностью воздуха и (или) почвы. Категория гигрофитов выделяется на основе особенностей физиологии и морфологии растений; наиболее характерный признак гигрофитов — отсутствие приспособлений, ограничивающих расходование влаги.
Ценоге́нное по́ле образуется при слиянии фитогенных полей в процессе смыкания крон и образования биогрупп деревьев.
Экологи́ческие фа́кторы — свойства среды обитания, оказывающие какое-либо воздействие на организм. Например, наличие минеральных веществ, доступ кислорода, влажность почвы, температура почвы, рыхлость почвы. Индифферентные элементы среды, например инертные газы, экологическими факторами не являются.
Ко́рень (лат. radix) — осевой, обычно подземный вегетативный орган высших сосудистых растений, обладающий неограниченным ростом в длину и положительным геотропизмом. Корень осуществляет закрепление растения в почве и обеспечивает поглощение и проведение воды с растворёнными минеральными веществами к стеблю и листьям.
Карликовые растения — растения низкого роста, по естественным или искусственным причинам имеющие отклонение от развития, нормального для данного вида или рода.
Аэренхима (или эренхима) — воздухоносная ткань у растений, построенная из клеток, соединённых между собой так, что между ними остаются крупные заполненные воздухом пустоты (крупные межклетники).
Почва — природное тело, формирующееся в результате преобразования поверхностных слоёв суши Земли при совместном воздействии факторов почвообразования. Состоит из почвенных горизонтов, образующих почвенный профиль, характеризуется плодородием. Многообразие почв отражено в разных типах почв.
Зимосто́йкость — способность растений противостоять комплексу воздействий внешней среды на протяжении зимнего и ранневесеннего периодов.
Геометрические закономерности в природе проявляются в виде повторяющихся форм и их сочетаний (паттернов). Они проявляются в различных природных объектах и явлениях, а иногда могут быть описаны при помощи математических моделей. Повторяющиеся элементы в природе принимают различные формы и проявляются в симметрии, деревьях, спиралях, изгибах рек, волнах, пене, геометрических узорах, трещинах, полосках и т. д.. Уже первые древнегреческие философы, такие как Платон, Пифагор и Эмпедокл, изучали такие.
В биологии развития фотоморфогенез — это процессы, происходящие в растении под влиянием света различного спектрального состава и интенсивности. Фотоморфогенез растений, как правило, исследуется с использованием источников света с контролируемым спектром.
Ретарданты — синтетические вещества разной химической природы, которые подавляют рост стеблей и побегов. Представляют собой одну из разновидностей регуляторов роста. Основной целью применения ретардантов является получение растений с сильным ветвлением, крепким стеблем и мощной корневой системой.
Кауле́рпа (лат. Caulerpa) — род морских зелёных водорослей из порядка бриопсидовых (Bryopsidales), выделяемый в монотипное семейство каулерповых (Caulerpaceae). Как и другие бриопсидовые, лишены межклеточных перегородок (септ), так что всё слоевище, или таллом, которое зачастую обладает внушительными размерами и причудливой формой, представляет собой единственную клетку с многочисленными ядрами. Некоторые виды этого растения способны достигать длины 2,8 м, что позволяет считать их крупнейшим одноклеточным.
Корневые клубеньки встречаются на корнях растений (преимущественно у семейства бобовых), которые ассоциированы с симбиотическими азотфиксирующими бактериями.
Расте́ния-поду́шки, или подушкообра́зные растения, или растения подушкови́дной фо́рмы — жизненная форма растительных организмов, для которой характерны многочисленные короткие, интенсивно ветвящиеся побеги, без явно выраженного главного ствола, между которыми накапливается субстрат состоящий из отмерших частей растения и характеризующийся своеобразной, круглой, подушкообразной формой. Годичный прирост всех побегов одинаков и ничтожен по величине. Принадлежат к различным семействам и родам (азорелла.
Структура почвенного покрова (СПП) — закономерное пространственное размещение почв на небольших территориях, выявляемое при детальном картографировании их почвенного покрова и образованное многократным повторением одного или нескольких различных основных образующих её элементов — почвенных комбинаций (ПК), каждая из которых содержит все почвы, являющиеся компонентами СПП с характерными для них взаимосвязями.
Эпибле́ма (греч. ἐπίβλημα – покрывало, покрытие от греч. ἐπί — на, над и греч. βλημα — бросаю, кладу) или ризоде́рма, — первичная покровная ткань молодых корней растений. Формирует корневые волоски.
Моро́зная тре́щина, или морозобо́й (неофиц.), или морозобо́ина (неофиц.) — повреждение древесины стволов крупных деревьев в виде радиальной трещины, возникающее под действием резкого их охлаждения. В зависимости от степени развития, на поверхности ствола такие повреждения наблюдаются как открытые или закрытые трещины со вздутиями или гребнями, возникающими снаружи ствола от разрастания коры и древесины, реже — как открытые без вздутий. Является пороком древесины из группы трещин.
Дерматоге́н (от греч. дермато и ген) — относительно однородный слой поверхностных клеток у растений, получивший своё название от того, что из него образуется впоследствии кожица (δέρμα — кожа). Расположен снаружи, имеет плотную структуру, относится к первичной образовательной ткани (меристема), расположен в растущих кончиках корней. Дерматоген имеет межклетники, его клетки обычно делятся так, что образующиеся перегородки имеют антиклинальное направление, то есть они растут перпендикулярно к поверхности.
Дѐндрохроноло́гия (от др.-греч. δένδρον — дерево, χρόνος — время, λόγος — слово, учение) — научная дисциплина о методах датирования событий, природных явлений, археологических находок и древних предметов, основанная на исследовании годичных колец древесины.
Лист (множ. ли́стья, собир. листва́; лат. folium, греч. φύλλον) — в ботанике наружный орган растения, основными функциями которого является фотосинтез, газообмен и транспирация. Для этой цели лист, как правило, имеет пластинчатую структуру, чтобы дать клеткам, содержащим в хлоропластах специализированный пигмент хлорофилл, доступ к солнечному свету. Лист также является органом дыхания, испарения и гуттации (выделения капель воды) растения. Листья могут задерживать в себе воду и питательные вещества.
Псилотовидные (лат. Psilotopsida) — класс высших растений, который включает в себя одно семейство — Псилотовые. Эти растения лишены корней, часто являются эпифитами на стволах пальм, древовидных папоротников, саговников; растут на почвах с большим количеством органических веществ и в трещинах скал.
Эффект лотоса — эффект крайне низкой смачиваемости поверхности, который можно наблюдать на листьях и лепестках растений рода Лотос (Nelumbo), и других растений, как например настурция, тростник обыкновенный и водосбор.
Миграция пестицидов в почвенном профиле происходит в основном за счет капиллярно-гравитационного перемещения воды. Скорость миграции и глубина проникновения пестицидов зависят от множества факторов, связанных как с почвенно-климатическими особенностями (гранулометрическим составом, содержанием коллоидов и сорбционной способностью почв, количеством осадков), так и со свойствами и дозами вносимых в почву препаратов.
Кривая светового насыщения фотосинтеза — это графическое представление эмпирической взаимосвязи между интенсивностью света и фотосинтезом. По сути своей она представляет собой модификацию уравнения Михаэлиса-Ментен. Кривая показывает положительную корреляцию между интенсивностью света и скоростью фотосинтеза: по оси х отложены значения независимой переменной (освещенность), а по оси y — значение зависимой переменной (скорость фотосинтеза).
Холодоустойчивость растений в значительной мере связана с преобладанием в молекулах фосфолипидов ненасыщенных жирных кислот, обладающих низкой температурой плавления, благодаря чему цитоплазматические мембраны даже при сильном охлаждении не застывают и сохраняют довольно высокую проницаемость для воды и растворенных веществ. Также характерна повышенная активность ферментов, в частности АТФаз, и способность к синтезу большого набора изоферментов, имеющих низкий температурный оптимум. Все, вышеперечисленные.
Транспирация — процесс движения воды через растение и её испарение через наружные органы растения, такие как листья, стебли и цветы. Вода необходима для жизнедеятельности растения, но только небольшая часть воды, поступающей через корни используется непосредственно для нужд роста и метаболизма. Оставшиеся 99-99,5 % теряются через транспирацию. Поверхность листа покрыта порами, называемыми устьицами и у большинства растений большая часть устьиц находится на нижней части листа. Устьица ограничены замыкающими.
Аменсали́зм (от лат. mensa — трапеза) — тип межвидовых взаимоотношений, при котором один вид, именуемый аменсалом, претерпевает угнетение роста и развития, а второй, именуемый ингибитором, таким испытаниям не подвержен.Антибиоз и аллелопатия — схожие виды взаимоотношений.
Экосистема озера включает биотические (живые) растения, животных и микроорганизмы, а также абиотические (неживые) физические и химические взаимодействия.Озерные экосистемы являются яркими примерами стоячих экосистем. К стоячим относятся застойные или слаботекущие воды (от латинского Lentus-вялый). Стоячие воды колеблются от прудов, озер до водно-болотных угодий, и большая часть этой статьи относится к стоячим экосистемам в целом. Стоячие экосистемы можно сравнить с текучими экосистемами, в которые.
hidrotropismo это реакция роста растений на концентрацию воды. Ответ может быть положительным или отрицательным.
Корни, например, положительно гидротропны, так как рост корней растений происходит в направлении более высокого уровня относительной влажности. Растение может обнаружить это в корневой крышке и затем послать сигналы на удлиненную часть корня..
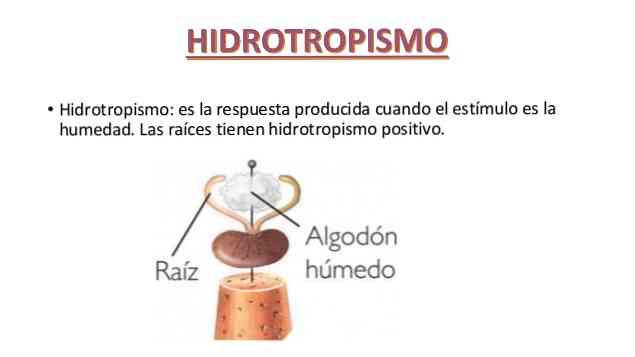
Положительный гидротропизм - это тот, в котором тело имеет тенденцию расти к влаге, в то время как отрицательный гидротропизм - это когда организм от него отрастает..
Гидротропизм - это форма тропизма (ориентировочная реакция организма на раздражитель), характеризующаяся ростом или реакцией движения клетки или организма на влагу или воду..
Механизм гидротропизма
Класс гормонов растений, называемый ауксинами, координирует этот процесс роста корней..
Ауксины играют ключевую роль в сгибании корней растений к воде, потому что они заставляют одну сторону корня расти быстрее другой, и, следовательно, сгибание корня.
Процесс гидротропизма начинается с того, что капюшон корня захватывает воду и посылает сигнал в удлиненную часть корня..
Гидротропизм трудно обнаружить в подземных корнях, так как корни не легко заметны.
Вода легко перемещается в почве, и содержание воды в почве постоянно меняется, поэтому любой градиент влажности почвы не является стабильным.
Почему гидротропизм так важен для растений?

Эта способность сгибать и выращивать корень в направлении градиента влажности, который обеспечивает гидротропизм, имеет важное значение, поскольку растениям для роста необходима вода. Вода вместе с растворимыми минеральными питательными веществами поглощается корневыми волосками..
Затем в сосудистых растениях вода и минералы транспортируются во все части растения через транспортную систему, называемую ксилемой..
Вторая транспортная система у сосудистых растений называется флоэмой. В флоэме также содержится вода, не содержащая растворимых минералов, но главным образом с растворимыми органическими питательными веществами..
Это имеет биологическое значение, так как гидротропизм помогает повысить эффективность растения в его экосистеме..
Заблуждения о гидротропизме
1- Гидротропизм и рост корней во влажных областях
Больший рост корней во влажных областях почвы, чем в сухих областях почвы, обычно не является результатом гидротропизма.
Гидротропизм требует, чтобы корень изогнулся от сушилки до влажного участка почвы. Корни требуют воды, чтобы расти, поэтому корни, которые оказываются во влажной почве, будут расти и разветвляться гораздо больше, чем в сухой почве..
2- Поглощение воды
Корни не могут чувствовать воду в трубах неповрежденной из-за гидротропизма и должны ломать трубы, чтобы получить воду.
3- Расстояние, необходимое для поглощения воды
Корни не могут чувствовать воду в нескольких футах от гидротропизма и расти к ней.
В лучшем случае гидротропизм, вероятно, действует на расстоянии нескольких миллиметров..
Исследования гидротропизма
Исследования гидротропизма были в первую очередь лабораторным явлением для корней, выращенных во влажном воздухе вместо почвы.
Его экологическое значение в корнях, культивируемых в почве, неясно, потому что так мало исследований гидротропизма изучило корни, культивируемые в почве..
Недавнее выявление мутантного растения, лишенного гидротропного ответа, помогло выяснить его роль в природе.
Гидротропизм может быть важен для растений, выращиваемых в космосе, где он может позволить ориентироваться корням в условиях микрогравитации..
На самом деле, этот ответ на рост растений не так легко изучить. Эксперименты, как уже упоминалось, проводятся в лабораториях, а не в естественной среде.
Тем не менее, каждый раз, когда вы узнаете больше о сложной природе этого процесса роста растений.
Наиболее популярные растения для изучения этого эффекта: горох (Pisum Sativum), кукурузное растение (Zea mays) и тале кислый (Arabidopsis thaliana).
Другой подход к изучению гидротропизма заключается в использовании инструментов для изменения направления вектора силы тяжести, получаемого растениями..

Хотя невозможно устранить влияние гравитации на Землю, существуют машины, которые вращают растения вокруг оси или, в некоторых случаях, в трех измерениях в попытке нейтрализовать влияние гравитации, которые называются позиционирующими машинами. случайный.
Фактически, гидротропизм в корнях был более очевидным, когда растения гороха и огурца выращивались на одной из этих машин.
Еще более интересный подход к изучению заключается в использовании условий микрогравитации, присутствующих во время космического полета..
Идея состоит в том, что в отсутствие значительных гравитационных сил преобладающая гравитропическая реакция корней эффективно отрицается, так что другие корневые тропизмы (такие как гидротропизм) становятся более очевидными, выше гравитропизма. Это поворотное или растущее движение растения или гриба в ответ на гравитацию.
Еще одним препятствием для изучения гидротропизма является сложность создания системы, в которой существует воспроизводимый градиент влажности..
Классические методы немецких ботаников, также используемые Дарвином, включали помещение семян в висящий цилиндр из влажных опилок, в результате чего корни сначала росли вниз, а затем снова росли во влажном субстрате..
Следует отметить, что одним из менее известных тропизмов является гидротропизм, рост, направленный в ответ на градиенты воды или влаги..
Хотя гидротропизм был изучен в корнях растений немецкими ботаниками девятнадцатого века и дарвинами, существование этого тропизма ставилось под сомнение до последних лет.
Эти процессы просто должны быть более изучены. Каждое научное исследование будет углублять понимание этих сложных механизмов.
Геотропизм заключается в изгибе корня по направлению действия силы тяжести (положительный геотропизм), а стебля в противоположном направлении (отрицательный геотропизм). Если перевернуть проросток корнем вверх, а стеблем вниз, то через небольшой промежуток времени корень изогнется вниз, а стебель вверх, т. е. они примут нормальное для себя положение. Совершенно ясно, что если бы растения не обладали такой способностью, то должны были бы погибать в массовом количестве. Очевидно, в процессе естественного отбора сохранились лишь растения, выработавшие в себе эту способность. Вернее будет сказать, что само образование стебля и корня стало возможным лишь благодаря наличию геотропической реакции, возникшей, очевидно, вместе с образованием данных органов, а может быть, и еще раньше, так как многие водоросли обладают отрицательным геотропизмом.
В понимании геотропизма до середины XX века царила гипотеза Холодного—Вента, объяснявшая следующим образом неравномерный рост стебля и корня. Авторы этой теории считают, что неравномерно распределяющийся ауксин вызывает различную реакцию стебля и корня. По их мнению, корень очень чувствителен к высокой концентрации ауксина, и те его концентрации, которые ускоряют рост стебля, замедляют рост корня. Поэтому нижняя поверхность корня, где скопляется больше ауксина, растет медленнее, и корень изгибается книзу, а у стебля, наоборот, быстрее, и он поэтому изгибается кверху.
Данное заключение авторы теории сделали на основании косвенных данных. Методика опытов Н. Г. Холодного была следующей. Для опытов срезались колеоптили кукурузы и прикладывались к корням люпина или кукурузы, у которых предварительно были отрезаны верхушки корня. В этих условиях колеоптили тормозили рост корней. На этом основании и было сделано заключение о том, что выделяемые точкой роста стебля ауксины тормозят рост корня и стимулируют рост стебля. Установленные авторами факты были вполне правильными, но заключение оказалось неверным. Дело в том, что колеоптиль выделяет очень большие количества ауксина, которые уже тормозят рост корня. В неповрежденных корнях, верхушки которых сами выделяют ауксин, все происходит совершенно иным образом. Непосредственные наблюдения за распределением гетероауксина в стебле и корне ряда растений при геотропическом изгибе и при нормальном росте показали наличие большого содержания гетероауксина в тех местах, где наблюдается наиболее интенсивный рост. Для определения гетероауксина использовался микрохимический метод. Срезы растений обрабатывались раствором железоаммиачных квасцов (0,5%) и серной кислотой (разбавленной до 25—50% по объему) и выдерживались в термостате при 40—50 °C. При наличии гетероауксина происходит розовато-фиолетовое окрашивание. При нормальном росте происходит равномерное распределение гетероауксина, а при геотропическом изгибе гетероауксин распределяется неравномерно. Гетероауксина больше там, где происходит более интенсивный рост. При этом вопреки теории Холодного и в стебле и корне гетероауксин скопляется одинаково на той стороне, где идет более интенсивный рост (рис. 207).
Очевидно, в процессе эволюции сохранились только те формы, у которых под влиянием действия силы тяжести происходило различное распределение протоплазмы в стебле и корне. Характер этого распределения у разных растений был, несомненно, неодинаков. У большинства растений под влиянием силы тяжести в стеблях плазма скопляется на нижней поверхности клеток, а у корней на верхней. При этом допущении сразу же станет ясным, что у обоих органов идет рост интенсивнее там, где больше скопляется протоплазмы. При вертикальном росте протоплазма распределяется в клетке равномерно. Предлагаемая нами концепция гораздо проще и понятнее объясняет не только причину отрицательного геотропизма стебля и положительного геотропизма корня, но и плагиотропный рост растительных организмов, т. е. рост, параллельный земной поверхности, особенно резко выраженный у корневищ многих растений. Таким образом, понять тропизмы растения можно, только пользуясь историческим методом, значение которого так хорошо в свое время подчеркнул К. А. Тимирязев. Возникновение самого механизма изгиба (неравномерное распределение протоплазмы) можно понять, только учтя историческую и творческую роль отбора, а также и приспособительную реакцию растения к условиям существования и их изменениям, меняющим и природу самого растения.
Под действием различных раздражителей проницаемость мембран меняется. Это приводит к изменению величины биопотенциалов и возникновению токов действия. Возбуждение, вызванное раздражителем, может передаваться по растению от корней к листьям, регулируя, например, работу устьиц, скорость фотосинтеза. При изменении освещения, температуры воздуха токи действия могут передаваться и в противоположном направлении – от листьев к корням, что приводит к изменению активности работы корня.
Интересно, что вверх по растению биотоки распространяются в 2,5 раза быстрее, чем вниз.
С наибольшей скоростью возбуждение у растений идет по проводящим пучкам, а в них – по клеткам-спутницам ситовидных трубок. Скорость распространения потенциала действия (электрических импульсов) по растению у различных видов не одинакова. Быстрее всех реагируют насекомоядные растения и мимоза – 2–12 см/с. У других видов растений эта скорость значительно ниже – около 25 см/мин.
41. Опыт с зеленой горошиной
Этот опыт впервые был поставлен крупнейшим исследователем проблемы раздражимости растений индийским ученым Д.Ч. Босом. Он показывает, что резкое повышение температуры вызывает в семенах появление токов действия. Для опыта нужны несколько зеленых (несозревших) семян гороха посевного (бобов, фасоли), гальванометр, препаровальная игла, спиртовка.
Соедините внешнюю и внутреннюю части зеленой горошины с гальванометром. Очень осторожно в бюксе нагрейте горошину (не повреждая) приблизительно до 60 °С.
При повышении температуры клеток гальванометр регистрирует разность потенциалов до 0,1–2 В. Вот что отметил по поводу этих результатов сам Д.Ч. Бос: если собрать 500 пар половинок горошин в определенном порядке в серии, то суммарное электрическое напряжение составит 500 В.
Самыми чувствительными у растений являются клетки точек роста, находящиеся на верхушках побегов и корней. Многочисленные, обильно ветвящиеся побеги и быстро нарастающие в длину кончики корней как бы ощупывают пространство и передают информацию о нем в глубь растения. Доказано, что растения воспринимают прикосновение к листу, реагируя на него изменением биопотенциалов, перемещением электрических импульсов, изменением скорости и направления передвижения гормонов. Например, кончик корня реагирует более чем на 50 механических, физических, биологических факторов и всякий раз при этом выбирает наиболее оптимальную программу для роста.
Убедиться в том, что растение реагирует на прикосновения, особенно частые, надоедливые, можно на следующем опыте.
42. Стоит ли трогать растения без надобности
Познакомьтесь с тигмонастиями – двигательными реакциями растений, вызванными прикосновениями.
Для опыта в 2 горшка высадите по одному растению, желательно без опушения на листьях (бобы, фасоль). После появления 1–2 листьев начинайте воздействие: листья одного растения слегка трите между большим и указательным пальцем 30–40 раз ежедневно в течение 2 недель.
К концу второй недели различия будет видны отчетливо: растение, подвергавшееся механическому раздражению, отстает в росте (рис. 20).
Рис. 20. Влияние на рост растений механического воздействия
Результаты опыта свидетельствуют, что длительное воздействие на клетки слабыми раздражителями может привести к торможению процессов жизнедеятельности растений.
Постоянным воздействиям подвергаются растения, высаженные вдоль дорог. Особенно чувствительны ели. Их ветви, обращенные к дороге, по которой часто ходят люди, ездят машины, всегда короче ветвей, расположенных на противоположной стороне.
Раздражимость растений, т.е. их способность реагировать на разные воздействия, лежит в основе активных движений, которые у растений не менее разнообразны, чем у животных.
Перед тем как приступить к описанию опытов, раскрывающих механизм движения растений, целесообразно ознакомиться с классификацией этих движений. Если растения на осуществление движений затрачивают энергию дыхания, это физиологически активные движения. По механизму изгиба они подразделяются на ростовые и тургорные.
Ростовые движения обусловлены изменением направления роста органа. Это сравнительно медленные движения, например изгибы стеблей к свету, корней к воде.
Тургорные движения осуществляются путем обратимого поглощения воды, сжатия и растяжения специальных двигательных (моторных) клеток, расположенных у основания органа. Это быстрые движения растений. Они свойственны, например, насекомоядным растениям, листьям мимозы.
Более подробно типы ростовых и тургорных движений будут рассмотрены ниже по мере выполнения опытов.
Для осуществления пассивных (механических) движений прямых затрат энергии клетки не требуется. В механических движениях в большинстве случаев цитоплазма не участвует. Наиболее распространены гигроскопические движения, которые вызываются обезвоживанием и зависят от влажности воздуха.
Гигроскопические движения
В основе гигроскопических движений лежит способность оболочек растительных клеток к поглощению воды и набуханию. При набухании вода поступает в пространство между молекулами клетчатки (целлюлозы) в оболочке и белка в цитоплазме клетки, что приводит к значительному увеличению объема клетки.
43. Движения чешуй шишек хвойных, сухого мха, сухоцветов
Изучите влияние температуры воды на скорость движения семенных чешуй шишек.
Для опыта нужны по 2–4 сухие шишки сосны и ели, высушенные соцветия акроклиниума розового или гелихризума большого (бессмертники), сухой мох кукушкин лен, часы.
Рассмотрите сухую шишку сосны. Семенные чешуи подняты, хорошо видны места, к которым были прикреплены семена (рис. 21).
Рис. 21. Шишки сосны
Опустите половину шишек сосны в холодную воду, а вторую – в теплую (40–50 °С). Наблюдайте за движением чешуй. Отметьте время, которое потребовалось для полного их смыкания.
Достаньте шишки из воды, стряхните и проследите за движением чешуй в процессе высыхания.
Отметьте время, за которое чешуи вернутся в исходное состояние, занесите данные в таблицу.
Повторите опыт с теми же шишками несколько раз. Это позволит не только получить более точные данные, но и убедиться в обратимости изучаемого вида движений.
Результаты опыта позволят сделать важные выводы.
1. Движение семенных чешуй шишек обусловлено потерей и поглощением ими воды. Об этом же свидетельствует прямая зависимость движения чешуй от температуры воды: при ее повышении скорость движения молекул воды возрастает, набухание чешуй происходит быстрее.
2. Чтобы набухание чешуй могло изменить их положение в пространстве, строение и химический состав клеток на внешней и внутренней стороне чешуи должны быть различными. Это действительно так. Оболочки клеток верхней стороны чешуй шишек хвойных более эластичны, растяжимы по сравнению с клетками нижней стороны. Поэтому при погружении в воду они поглощают ее больше, быстрее увеличивают свой объем, что приводит к удлинению верхней стороны и движению чешуи вниз. В процессе обезвоживания клетки верхней стороны теряют воду тоже быстрее клеток нижней стороны, что приводит к загибанию чешуи вверх.
Интересно наблюдать вызываемые набуханием движения листьев кукушкина льна либо других листостебельных мхов. У живых растений листья направлены в сторону от стебля, а у сухих – прижаты к нему. Если опустить сухой стебелек в воду, через 1–2 мин листья переходят из вертикального положения в горизонтальное.
Очень красивы движения высушенного соцветия бессмертника. Если сухое соцветие опустить в воду, через 1–2 мин листочки обертки приходят в движение и соцветие закрывается.
Задание. Сравните скорость движения чешуй шишек различных видов хвойных. Зависит ли она от размера шишек? Сравните скорость движения чешуй шишек сосны и ели, листьев мхов и листочков обертки соцветия бессмертника, выявите черты сходства и различия.
44. Гигроскопические движения семян. Гигрометр из семян аистника
Гигроскопические движения играют важную роль в распространении семян различных растений.
Изучите механизм самозакапывания семян аистника, перемещения по почве семян василька полевого.
Для опыта нужны семена аистника (грабельника), василька синего, лист плотной бумаги, часы, предметное стекло.
Аистник – распространенное в Белоруссии растение. Свое название получило благодаря сходству плода с головой аиста (рис. 22).
Рис. 22. Аистник
Рассмотрите внимательно строение сухого плода аистника. Доли зрелого коробочковидного плода снабжены длинной остью, в нижней части спирально закрученной. Плод покрыт жесткими волосками.
На предметное стекло нанесите каплю воды и опустите в нее сухой плод. Закрученная спиралью нижняя часть начинает раскручиваться, и плод, не имеющий опоры на стекле, совершает вращательные движения.
После полного выпрямления ости перенесите плод на сухую часть стекла. По мере высыхания нижняя часть снова закручивается в спираль и вызывает вращение плода.
Проведите хронометраж опыта, сравнивая скорости процессов раскручивания и закручивания спирали.
Механизм движения плода аистника тот же, что и чешуй шишек хвойных, – различие в гигроскопичности клеток ости.
Наблюдения за движением плода в капле воды позволяют понять поведение его в почве. Когда плод падает на землю, верхний конец ости, загнутый под прямым углом, цепляется за окружающие его стебельки и остается неподвижным. При закручивании и раскручивании спирального участка нижняя часть плода с семенем ввинчивается в землю. Путь назад преграждают жесткие, отогнутые вниз волоски, покрывающие плод.
Чтобы изготовить примитивный гигрометр, в кусочке картона или дощечке, покрытой белой бумагой, проделайте отверстие и закрепите в нем нижний конец плода. Для калибровки прибора сначала высушите, затем смочите ость водой и отметьте крайнее положение (рис. 23). Размещать прибор лучше на улице, где колебания влажности выражены более резко, чем в помещении.
Рис. 23. Гигрометр из аистника
Аистник – не единственное растение, способное к самозакапыванию семян. Сходное строение и механизм распространения имеют ковыли, овсюг, лисохвост.
Плоды василька (семянки с хохолком из твердых щетинок) не способны к самозакапыванию. При колебаниях влажности почвы щетинки попеременно опускаются и поднимаются, толкая плод вперед.
Задание. Соберите семена василька, лисохвоста, овсюга. Изучите поведение их во влажной и сухой среде, сравните с аистником.
Тропизмы
Умнейшее создание природы,
Всегда растущее из рода в роды –
В земле корнями, в небе – головой.
45. Гидротропизм корня
Одно из наиболее интересных видов движения – движение корня к воде (гидротропизм). Наземные растения испытывают постоянную потребность в воде, поэтому корень всегда растет в ту сторону, где содержание воды выше. Гидротропизм присущ прежде всего корням высших растений. Наблюдается также у ризоидов мхов и заростков папоротников. Для опыта нужно 10–20 наклюнувшихся семян гороха (люпина, ячменя, ржи), 2 чашки Петри, немного пластилина.
Плотно прикрепленным ко дну пластилиновым барьером разделите площадь чашки на 2 равные части. На барьер положите наклюнувшиеся семена, слегка вдавливая их в пластилин, чтобы при росте корня семена не сдвинулись с места. Корешки должны быть направлены строго вдоль барьера (рис. 24).
Рис. 24. Схема расположения семян при изучении гидротропизма корня
Эти этапы работы в контрольной и опытной чашках одинаковы. Теперь предстоит создать различные условия увлажнения. В контрольной чашке влажность в левой и правой частях должна быть одинакова. В опытной чашке вода наливается только в одну половину, а вторая остается сухой.
Обе чашки накройте крышками и поместите в теплое место. Ежедневно наблюдайте за положением корешков. Когда ориентация их станет хорошо заметной, подсчитайте количество семян, корни которых проявили положительный гидротропизм (рост органа в сторону воды).
Наблюдения за движением корешка к воде ясно показывают, что тропизмы – это ростовые движения. Корешок растет в сторону воды, при этом происходит, если это необходимо растению, изгиб корня.
Гидротропизм – частный случай хемотропизма (ростовой реакции растений на неравномерное распределение в окружающей среде какого-либо вещества). Способность корней растений, грибных гифов, пыльцевых трубок, проростков паразитических растений (повилики, например) распознавать химическое вещество на некотором расстоянии от него удивительна. Установлено, что воспринимает воздействие химических веществ зона роста органа, а изгиб образуется на некотором расстоянии от нее, т.е. происходит передача раздражения по корню (рис. 25).
Рис. 25. Хемотропический изгиб корней
Задание. По описанной выше схеме опыта проверьте способность растений распознавать не только воду, но и нужные растению растворы минеральных солей, например 0,3%-ный раствор нитрата калия или нитрата аммония.
46. Влияние силы земного тяготения на рост стебля и корня
Задание. Изучите характер геотропической реакции стеблей разного порядка двудольного растения. Для этого вырастите проростки, закройте поверхность почвы, чтобы она не высыпалась, и переверните горшки. Наблюдения ведите до тех пор, пока не появятся боковые стебли первого и второго порядка.
47. Влияние этилена на геотропическую реакцию проростков гороха
Рост растений регулируется не только биоэлектрическими сигналами, но и гормональной системой. Главную роль в регуляции скорости роста играет количественное содержание гормона ауксина и его взаимодействие с другими гормонами, в частности абсцизовой кислотой и этиленом.
В отличие от стимулирующего рост ауксина абсцизовая кислота тормозит деление клеток нижней стороны органа. Это вызывает замедление ее роста, и корень начинает изгибаться по направлению к центру Земли.
Для опыта нужны зрелые яблоки (источник этилена), 2 стеклянных колпака, 2 горшка с проростками гороха.
Стеклянные колпаки установите на подставку. Под ними разместите горшки с 2–3-дневными проростками гороха. В опытном варианте под колпак положите яблоки. Растения поставьте в темноту.
По мере накопления этилена в воздухе он начинает проникать в проростки гороха. Через несколько дней становятся заметны нарушения нормальной отрицательной геотропической реакции побегов, которые начинают расти горизонтально, а при высокой концентрации этилена в воздухе даже полегают.
Результаты опыта свидетельствуют о регуляторных функциях этилена в жизни растений. Увеличение его содержания в клетках приводит к изменению скорости их роста.
Задание. Изучите влияние этилена на рост проростков томатов.
Естественно, геотропическая ориентация органов растений в непрерывно меняющихся условиях среды не может всегда оставаться постоянной. По мере формирования и распускания бутонов изменяется ориентация цветоножки, например у мака. Молодые ветки ели растут под более острым углом, чем старые.
Можно изучить смену отрицательного геотропизма цветоножек арахиса (земляного ореха) на положительный, вырастив его в комнатных условиях. После отцветания цветоножка арахиса, на которой сидит завязь, удлиняется, загибается к земле и углубляется в нее. Таким образом, цветки находятся над землей, а плоды созревают в земле. Хотя это ограничивает способность вида к распространению, созревшие семена находятся в идеальных условиях для прорастания.

18.12.2018
Наблюдая окружающий нас растительный мир, можно заметить много удивительных особенностей, общих для большинства его представителей. Например, то, что ростовые процессы в растениях осуществляются по определенным правилам. Рост корневой системы происходит в направлении к центру Земли, а основной стебель развивается в противоположную сторону, благодаря чему растения занимают вертикальное положение.
Каждый отдельно взятый растительный организм использует собственные ростовые процессы для наиболее удобного и безопасного расположения в окружающем пространстве. Поэтому направление роста определенных частей растения связано с односторонним направленным действием различных раздражителей окружающей среды (сила гравитации, световое излучение, химические факторы и пр.). Это биологическое явление получило название тропизм (от греч. τροπος – направление, поворот).
Различают такие типы тропизма у растений:
- геотропизм или гравитропизм – движение, рост в ответ на действие гравитации;
- апогеотропизм – отрицательный геотропизм;
- гелиотропизм – суточное движение или сезонное движение частей растения в ответ на направление солнца (например, подсолнечника) и афелиотропизм – отрицательный гелиотропизм;
- фототропизм – движение или рост в ответ на световой раздражитель, а также апототропизм – отрицательный фототропизм;
- гидротропизм – движение или рост в ответ на воду и гигротропизм – движение или рост в ответ на влагу или влажность (у растений корневая система ощущает изменения влажности в почве и сигнализирует об этом, вызывая клеточные преобразования, которые заставляют корень искривляться к области повышенной влажности);
- аэротропизм – рост растений по направлению к источнику кислорода или от него;
- термотропизм – движение или рост в ответ на направленное температурное воздействие;
- магнитотропизм – движение или рост растения под действием магнитного поля;
- электротропизм – движение или рост в ответ на действие источника электрического поля;
- хемотропизм – движение или рост в ответ на химическое воздействие;
- тигмотропизм – движение или рост растения в ответ на прикосновение.
Если направление роста совпадает с направлением действия влияющего фактора, то явление тропизма называется положительным, а если рост происходит в прямо противоположном направлении, то имеет место отрицательный тропизм.

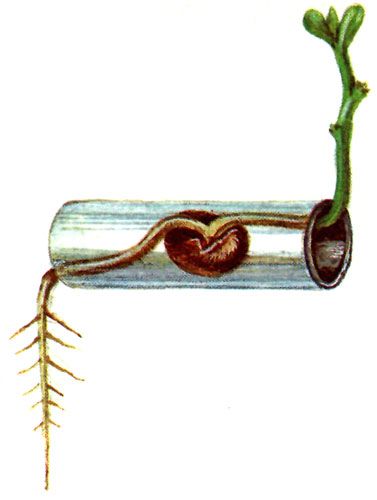
Геотропизм присущ всем высшим и многим низшим растениям. Впервые способность растений занимать вертикальное положение в пространстве была доказана Чарльзом Дарвином. Согласно его наблюдениям центральные корни растений развиваются в направлении действия силы земного притяжения, стремясь достичь влаги в глубинных слоях почвы. А стебли и побеги – наоборот, растут в противоположную сторону, навстречу теплу и свету солнечных лучей, преодолевая гравитацию. В первом случае наблюдается явление положительного геотропизма, во втором – отрицательного.
Геотропизм – это реакция растительного организма на силу гравитации, земного притяжения, которая определяет положение растений в пространстве. Существование геотропизма доказывает простой опыт: если растущее в горшке растение расположить горизонтально, то спустя непродолжительный отрезок времени в его растущих точках можно наблюдать изменения направления роста. Растущая часть стебля совершит поворот на 90° вверх, демонстрируя отрицательный геотропизм.

У прорастающего в темноте семени, независимо от ориентации его в пространстве, также происходит направленный рост зародышевого корешка в сторону действия силы тяжести, и в противоположную ему – у проросткового побега. Не менее показателен геотропизм растений на примере растущих вдоль горных склонов деревьев. Их стволы расположены не перпендикулярно к поверхности, а вдоль направления действия гравитационной силы. Геотропизм является неотъемлемой частью роста растений, ориентируя их положение для максимального контакта с солнечным светом, а также обеспечивая правильное направление роста корней.
Как и все типы тропизма, геотропизм происходит в результате четырех последовательных действий. Первое из них – это восприятие растением источника полезного ему стимулирующего воздействия гравитации (света, тепла, влаги и пр.). Затем происходит преобразование полученного сигнала, в результате которого гормоны роста ауксины, находящиеся в растущих частях растения, перераспределяются в его клетках. Четвёртый, заключительный этап: отображение произошедших действий на самих ростовых процессах. Рост за счет геотропизма обусловлен изменениями концентрации гормона роста ауксина в растительных клетках.
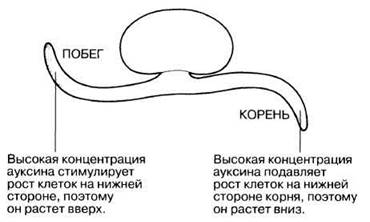
Изменение направления роста корня под действием силы земного тяготения происходит в ответ на информацию, полученную с помощью статолитов – особых органелл, расположенных в клетках на кончике корня. Статолиты также находятся в энтодермическом слое гипокотиля, стебля и соцветия. Геотропический сигнал приводит к перераспределению ауксина, известному как полярный перенос ауксина таким образом, чтобы вызвать дифференциальный рост, приводящий к искривлению корня (или стебля).
Полный процесс этого взаимодействия был описан в 1920-х годах ученым Киевского университета Н. Холодным (1927 г.) и Фрицем Вентом из Калифорнийского технологического института (1928 г.).
Читайте также: