
Геном прокариот и эукариот реферат
Обновлено: 04.07.2024
Название работы: Особенности генома вирусов, прокариот и эукариот
Предметная область: Биология и генетика
Описание: Гены одного оперона участка генетического материала который состоит из одного двух и больше сцепленных структурных генов которые кодируют белки ферменты которые осуществляют последовательные этапы биосинтеза какого-нибудь метаболита. Структура генов в бактериофагов и вирусов в основном похожая с бактериями но более осложненная и связанна с геномом хозяев.
Дата добавления: 2015-09-13
Размер файла: 22.52 KB
Работу скачали: 15 чел.
Особенности генома вирусов, прокариот и эукариот.
Ген представляет собой последовательность нуклеотидов ДНК размером от нескольких сотен до миллиона пар, в которых закодированная генетическая информация о первичной структуре белка (число и последовательность аминокислот). Для регулярного правильного считывания информации в гене должны быть: кодон инициации, смысловые кодоны и кодон терминации. Три подряд расположенные нуклеотида представляют собой кодон, что и определяет, какая аминокислота будет располагаться в определенной позиции в белке. Например, в молекуле ДНК последовательность ТАС является кодоном для аминокислоты метионина, а последовательность ТТТ кодирует фениланин.
Для прокариотических организмов является характерной относительно простая структура генов. Так, структурный ген бактерии, фага или вируса, как правило, контролирует лишь одну ферментативную реакцию. Специфической для прокариотов является оперонная система организации нескольких генов. Гены одного оперона (участка генетического материала, который состоит из одного, двух и больше сцепленных структурных генов, которые кодируют белки (ферменты), которые осуществляют последовательные этапы биосинтеза какого-нибудь метаболита). Опероны прокариотических организмов расположены в кольцевой хромосоме бактерии рядом и контролируют ферменты, которые осуществляют последовательные или близкие реакции синтеза (лактозный, гистидиновый и другие опероны).
Оперон начинается и заканчивается регуляторными участками - промотором в начале и терминатором в конце. Благодаря этому все гены, которые входят в состав оперона, могут синтезировать свои продукты вместе.
Концепция оперона была предложена 1961 г. французскими учеными Франсуа Жакобом и Жаком Моно, за что они получили Нобелевскую премию 1965 г.
Структура генов в бактериофагов и вирусов, в основном, похожая с бактериями, но более осложненная и связанна с геномом хозяев. Например, у фагов и вирусов выявлены перекрывание генов, а полная зависимость вирусов эукариот от метаболизма клетки-хозяина привела к появлению экзон-интронов в структуре генов.
А вот в оперон эукариотических организмов входит, как правило, лишь один структурный ген и его регуляторные элементы. Эукариотические гены, в отличие от бактериальных, имеют прерывчатое мозаичное строение. Кодирующие последовательности (экзони) чередуются с не кодирующими (интроны). Экзон участок гена, который несет информацию о первичной структуре белка. В гене экзоны разделенны некодирующими участками интронами. Интрон участок гена, который не несет информации о первичной структуре белка и расположен между экзонами. В результате подобной структуры гены эукариот имеют более длинную нуклеотидную последовательность, чем соответствующая зрелая и-РНК, последовательность нуклеотидов в которой отвечает последовательности экзонов. В процессе транскрипции информация о гене списывается с ДНК на промежуточную и-РНК, что которая из экзонов и интронов. Потом специфические ферменты - рестриктазы - разрезают эту про-и-РНК по границам экзон-интрон, после чего экзонные участки ферментативно соединяются вместе, образовывая зрелую м-рнк (этот процесс называется сплайсингом). Количество интронов может варьировать в разных генах от нуля до многих десятков, а длина - от нескольких пар основ до нескольких тысяч.
Рядом со структурными и регуляторными генами у эукаритоических организмов найдены участки повторяемых нуклеотидных последовательностей, функции которых изучены недостаточно, а также мигрирующие элементы (мобильные гены), способны перемещаться по геному. Найдено также так называемые псевдогены у эукариот, которые представляют собой копии известных генов, которые работают в других частях генома и лишены интронов или инактивированы мутациями.
Геном эукариот устроен намного сложнее, чем у прокариот. Генетический аппарат эукариотической клетки обособлен в виде клеточного ядра, внутри которого располагаются основные носители наследственности — хромосомы. Количество хромосом видоспецифично и колеблется от двух (лошадиная аскарида) до тысячи (низшие растения). Количество ДНК в клетках эукариот намного выше, чем у бактерий. Оно оценивается с помощью величины С — количества ДНК на гаплоидное число хромосом, т.е. на геном. Оно колеблется у разных видов от 10 4 до 10 11 и часто не коррелирует с уровнем организации вида. Самые большие значения величины С, превышающие содержание ДНК в геноме человека, характерны для некоторых рыб, хвостатых амфибий, лилейных.
Одной из особенностей генома эукариот является структурная и функциональная связь ДНК с белками. Она обусловлена особенностями процесса передачи генетической информации и регуляторной функцией белков. Информация передается от клетки к клетке в процессе сложного процесса клеточного деления (митоза или мейоза). Для полного и точного распределения ее между дочерними клетками в интерфазе происходит процесс удвоения количества ДНК, а в начале деления (профазе) — процесс конденсации интерфазных хромосом. В итоге хромосомы приобретают вид компактных плотных тел. Компактизация хромосом исключает риск их запутывания во время расхождения к разным полюсам в анафазе. В этих структурных преобразованиях хромосом участвуют ядерные белки — гистоны, которые осуществляют суперспирализацию ДНК. Гистоны выступают также в качестве регуляторов матричной активности интерфазных хромосом, т.к. связь гистона с функционирующим участком хромосомы переводит его в гетерохроматическое, т.е. сильно спирализованное и, следовательно, неактивное состояние.
Присутствие в составе эукариотических хромосом белков, количество которых удваивается синхронно с удвоением ДНК, делает процесс репликации хромосом более длительным.
Характерной особенностью генома эукариот является избыточность ДНК, количество которой намного превышает то, которое необходимо для кодирования структуры всех клеточных белков. Одной из причин избыточности является наличие повторяющихся последовательностей нуклеотидов. Их существование впервые было установлено в конце 60-х гг. ХХ в. американскими исследователями Р. Бриттеном и Д. Девидсоном при изучении кинетики ренатурации ДНК (воссоединения одиночных цепей). В настоящее время установлено, что в составе эукариотической ДНК присутствуют два типа повторов — умеренноповторяющиеся п.н. и высокоповторяющиеся п.н. Умеренные повторы встречаются в виде десятков и сотен копий; средний размер их составляет ≈ 300-400 п.н. Они могут быть прямыми и инвертированными (палиндромы). Между повторами располагаются неповторяющиеся участки ДНК. Высокоповторяющиеся п.н. представляют собой короткие фрагменты ДНК (десятки п.н.), которые представлены большим количеством копий (до 106). В ряде случаев состав оснований в этих повторах отличается от такового в геноме в целом, в результате чего повторы могут образовывать отдельную фракцию с определенной плавучей плотностью. Эта фракция называется сателлитной ДНК. Она никогда не транскрибируется, в связи с чем ее называют также “молчащей”. Установлено, что сателлитная ДНК локализована в гетерохроматических районах хромосом: в теломерах, около центромеры, в ядрышке. Считается, что она выполняет регуляторную функцию, обеспечивая структурные преобразования хромосом во время процесса передачи генетической информации от клетки к клетке.
Избыточность ДНК в геноме эукариот в значительной мере создается также за счет того, что в его составе много нуклеотидных последовательностей, которые не кодируют структуру белков. Некоторые из них входят в состав генов, как например, интроны — вставки. Кроме того, есть так называемые сигнальные последовательности, которые не транскрибируются, а служат лишь для связывания белков-регуляторов. К их числу относятся промоторы, участки, контролирующие спирализацию хромосом; участки прикрепления хромосом к веретену и др.
Лишь немногие гены присутствуют в эукариотическом геноме в единственной копии. Основная их масса представлена разным числом копий. Расположенные рядом идентичные гены образуют кластеры. Существование кластеров говорит о большой роли дупликаций генов в эволюции геномов. Пример кластеров: гены белков эритроцитов — глобинов. Гемоглобин является тетрамером, состоящим из 4-х полипептидных цепей: 2α и 2β. Каждый тип цепей кодируется генами, организованными в кластер. У человека α-кластер располагается в 11-й хромосоме, а β-кластер — в 16-й хромосоме. β-кластер занимает участок ДНК в 50 тыс. п.н. и включает в себя пять функционально активных генов и один псевдоген. Псевдогены — это нефункционирующие, реликтовые гены, произошедшие в результате мутационных изменений от некогда активных генов. Они не экспрессируются. Гены в составе кластера отделены друг от друга спейсерами — нетранскрибируемыми вставками, в которых иногда могут присутствовать регуляторные участки.
Основным отличием эукариотических генов от генов прокариот является то, что большинство из них имеют прерывистую структуру и состоят из кодирующих участков — экзонов и некодирующих вставок — интронов. Длина экзонов от 100 до 600 п.н., а интронов — от нескольких десятков до многих тысяч п.н. Интроны могут составлять до 75% от длины гена. Прерывистая структура генов создает основу для более тонкого контроля их работы.
В результате транскрипции прерывистых генов образуется первичный продукт — про-иРНК, которая является полной копией гена и содержит в себе участки, соответствующие как экзонам, так и интронам. В процессе транскрипции участвуют три разных типа РНК-полимераз, которые считывают разные гены. РНКП-I считывает гены, кодирующие структуру разных форм рРНК (5,8S, 18S, 28S). РНКП-II ведет транскрипцию генов, кодирующих структуру белков и некоторых мяРНК. И, наконец, РНКП-III считывает гены 5S рРНК, транспортных РНК и мяРНК. В инициации процесса транскрипции принимает участие белковый комплекс, состоящий из различного числа белковых факторов транскрипции. У млекопитающих в его состав входят 12-14 полипептидов с общей массой в 600 кДА. В регуляции интенсивности транскрипции принимают участие специфические регуляторные участки — энхансеры и сайленсеры. Первые усиливают, вторые ослабляют процесс транскрипции. Они могут быть удалены от промотора на тысячи п.н. Под их контролем синтезируются регуляторные белки. В процессе транскрипции промотор и энхансер (или сайленсер) сближаются за счет структурных изменений ДНК, и регуляторные белки взаимодействуют с факторами транскрипции или с РНК-полимеразой.
Для того, чтобы про-иРНК могла играть роль матрицы для синтеза белка, она должна пройти период созревания (процессинг). Главное событие этого периода — удаление из про-иРНК участков, соответствующих интронам, и соединение в единую цепочку оставшихся экзонов. Процесс “сшивания” экзонов называется сплайсингом. В осуществлении сплайсинга большая роль принадлежит малым ядерным РНК (мяРНК) и белкам. Процесс протекает аналогично у всех эукариот. Молекулы мяРНК комплементарно взаимодействуют как с про-иРНК, так и друг с другом. Они обеспечивают удаление интронов и удерживают экзоны вблизи друг от друга.
Процесс сплайсинга может носить альтернативный характер, т.е. сшивание экзонов может осуществляться в разных комбинациях. Многие гены содержат десяток и более экзонов, поэтому число вариантов зрелой иРНК = 2 n , где n — число экзонов. Альтернативный сплайсинг делает систему записи информации экономичной, так как с одного гена можно считывать информацию для синтеза разных белков. Кроме того, он создает возможность регулирования потока информации в зависимости от потребности клетки в том или ином белковом продукте. Альтернативный сплайсинг, в частности, используется при синтезе иммуноглобулинов, факторов транскрипции и других белков.
Полное созревание иРНК включает модификацию обоих ее концов: навешивание кэп-структуры с 5'-конца и присоединение полиадениловой цепочки с 3'-конца. Кэп-структура образуется за счет присоединения к концевому основанию иРНК 5'-конца гуанинового нуклеотида.
Схема сплайсинга

Механизм трансляции у эукариот принципиально не отличается от прокариотического. Однако в обслуживании этого этапа синтеза белка принимает участие значительно большее количество белковых факторов трансляции, чем у бактерий.
При характеристике структуры генома эукариот нельзя не сказать о специализированных концевых участках хромосом — теломерах. Теломерная ДНК состоит из многократно повторяющихся коротких блоков нуклеотидов. Впервые теломерная ДНК была изучена у одноклеточных простейших.
В ее состав входят блоки по 6-8 пар нуклеотидов. В одной цепи — это блок TTGGGG (G-богатая цепь), в другой — AACCCC (C-богатая цепь). У человека эта последовательность отличается одним основанием TTAGGG, у растений имеется универсальный блок TTTAGGG. Протяженность теломерной ДНК у человека колеблется от 2 до 20 тыс. п.н. Теломерная ДНК никогда не транскрибируется и входит в состав сателлитной ДНК. С теломерными районами хромосом взаимодействует фермент теломераза, который устраняет возникающие в них повреждения. С укорочением теломер в результате потери концевых участков, вызванной снижением активности этого фермента, связывают процесс старения клеток.
Существенным отличием функционирования эукариотического генома по сравнению с прокариотическим является многоуровневый характер регуляции действия генов. У прокариот возможен только один тип регуляции — на уровне транскрипции с помощью оперонной системы. У эукариот, благодаря прерывистой структуре генов, к этому типу регуляции добавляется еще посттранскрипционная (сплайсинг, модификация) регуляция и регуляция на уровне трансляции (неоднозначность трансляции). Кроме того, присутствие в хромосомах гистонов позволяет осуществлять групповой контроль за действием генов с помощью механизма структурных преобразований ДНК — перевода участков хромосом из активного (эухроматического) в неактивное (гетерохроматическое) состояние. Такие преобразования иногда затрагивают целые хромосомы и даже весь геном целиком. В качестве примера хромосомного уровня регуляции можно привести образование в клетках женского пола млекопитающих и человека полового хроматина (тельца Барра). Это — крупная гранула хроматина, представляющая собой одну из двух Х-хромосом, максимально конденсированную, и, следовательно, неактивную. Примером инактивации всего генома служит процесс спермиогенеза у животных, во время которого конденсацией охвачены все хромосомы сперматозоида, что делает их неактивными. Это является защитным механизмом для половых клеток в случае повреждения их ДНК (например, при облучении). Возникающие в них мутации, если они не летальны, могут проявиться только при восстановлении функциональной активности мужского генома при дифференциации зародыша. Однако рецессивность большинства мутаций отодвигает их возможное проявление, по крайней мере, до следующего поколения (до перехода в гомозиготное состояние) или вообще исключает его.
Перейти к чтению других тем книги "Генетика и селекция. Теория. Задания. Ответы":
Рефераты и конспекты лекций по географии, физике, химии, истории, биологии. Универсальная подготовка к ЕГЭ, ГИА, ЗНО и ДПА!
В ходе эволюции у эукариот сформировался геном, который существенно отличается от генома прокариот. Геном высших организмов в первую очередь характеризуется таким свойством как избыточность. Содержание ДНК у эукариот в расчете на 1 клетку в бреднем на 2-3 порядка выше, чем у прокариот
, и для разных организмов оно колеблется в интервале от 1010 до 1013 Да. Хотя такой размер генома теоретически позволяет кодировать свыше 106 различных белков, оценки числа функционирующих генов у эукариот колеблются в интервале от 104 до 27105. Возможно существуют "молчащие" гены, необходимые клетке для реализации редко встречающихся программ. Однако это не является единственной причиной большого количества ДНК в клетках эукариот. Повышение количества ДНК у эукариот связано прежде всего с усложнением регуляторных процессов в клетках. Это усложнение выражается в том, что размеры единиц транскрипции у эукариот, как правило, на порядок превышают размеры зрелых молекул мРНК, а также в том, что значительную часть генома у эукариот составляют последовательности, служащие исключительно для целей регуляции.
Однако общее количество ДНК в клетке нельзя рассматривать как простой показатель функциональной и структурной сложности организма (рис. 122). Например, у некоторых рыб количество ДНК на ядро в 10 раз больше, чем у человека. Содержание ДНК может сильно варьировать даже у отдельных организмов одного рода. Все это говорит о реальной избыточности ДНК в клетках высших организмов.
Другая структурная особенность генома эукариот - это блочный характер организации. Блочность строения проявляется на разных уровнях. Это и длинные блоки с разным CG- содержанием, и многочисленные короткие участки, сильно обогащенные определенными (пуриновыми или пиримидиновыми) нуклеотидами, и гомополимерные последовательности.
Блочность строения проявляется и в структуре индивидуальных генов, которые часто состоят из отделенных друг от друга зон (экзонов), соответствующих, по-видимому, отдельным доменам кодируемых ими белков, разделенных некодируемыми последовательностями (интронами). Наличие генов с интронами характерно для эукариот, тогда как в геноме прокариот интронов не обнаружено.
Геному эукариот свойственна также способность к перестройкам. Ярким примером геномных перестроек являются процессы перемещения отдельных участков генов, происходящие при формировании генов иммуноглобулинов. Другим примером может служить массовое увеличение числа копий (амплификация) некоторых структурных генов. Примером амплификации могут служить многочисленные повторы генов рРНК в петлях ядрышковых организаторов. Однако геномные перестройки, по-видимому, все-таки носять ограниченный характер, вследствие чего общий план организации генома отдельных организмов сохраняется на протяжении многих поколений.
Уникальные (неповторяющиеся) последовательности – наиболее сложный и, как правило, основной по массе компонент генома эукариот. Однако доля их у разных организмов может составлять от 15 до 98%. В целом процентное содержание уникальных последовательностей у низших эукариот заметно выше, чем у высших.
Особенно мало уникальной ДНК у растений. Во фракции уникальной ДНК находится большинство структурных генов эукариот. Однако далеко не все уникальные после- довательности представляют собой участки, кодирующие белки. Доля таких участков, как правило, не превышает 1 - 2%. Большая часть транскрибирующихся уникальных последовательностей не кодирует мРНК цитоплазмы, а выполняет какие-то специальные функции в ядре. Более половины уникальной ДНК вообще не транскрибируется в клетках, хотя и сохраняется в эволюции. Возможно она служит чисто структурным целям, возможно является средством борьбы с мутациями. Назначение информационной избыточности уникальной ДНК, как и избыточности ДНК вообще, - проблема, которая еще ждет свого решения.
В ДНК эукариот содержатся последовательности, повторяющиеся от нескольких десятков до миллиона раз на геном. Хорошо изученным компонентом часто повторяющихся последовательностей является сателлитная ДНК, которая обычно обнаруживается в виде отдельного (сателлитного) пика при центрифугировании молекул ДНК в градиентах плотности CsCl. Сателлитные ДНК состоят из коротких (2 - 20 пар нуклеотидов) повторяющихся последовательностей, образующих длинные блоки. Количество таких сателлитных ДНК в ДНК разных организмов варьирует от от 1 до 80%, а число отличающихся по плавучей плотности сателлитных ДНК в одном организме может достигать 10 - 12. Одной из интересных общих черт строения сателлитной ДНК является присутствие в каждой цепи лишь 2 - 3 из 4 возможных нуклеотидов. В хромосомах они локализуются преимущественно в центромерных районах, поэтому предполагаемые функции сателлитной ДНК связывают с ролью этих последова- тельностей в поддержании структурной целостности хромосом, в стабилизации центромер, с участием в узнавании гомологичных хромосом при мейозе, а также с участием в контроле за размерами ядра и клеточным ростом. Другой компонент часто повторяющихся последовательностей - это обращенные повторы (палиндромы). Значительная часть обращенных повторов транскрибируется в составе РНК и образуют структуры, называемые шпильки. Предполагается, что такие шпильки служат сигналами для ферментов, осуществляющих процессинг и сплайсинг РНК, ДНК-полимеразы и участвуют в терминации транскрипции для РНК-полимераз.
Наряду с частыми повторами в ДНК высших организмов присутствуют последовательности, повторяющиеся с меньшей частотой (от 10 до 104 копий на гаплоидный геном) - умеренно повторяющиеся последовательности. Это полифункциональный класс последовательностей, в который входят транскрибируемые структурные гены и контролирующие (регуляторные) элементы генома. Кроме того, предполагают, что в геноме млекопитающих некоторые семейства умеренных повторов имеют вирусную природу. В геноме эукариот находят также мобильные генетические элементы, аналогичные прокариотическим IS- и Tn-элементам. Большая часть изученных к настоящему времени генов у эукариот не являются уникальными, а представлены или множеством сходнях копий, или ограниченным набором дуплицированных и дивергировавших в разной мере последовательностей, называемыми мультигенными семействами. Повторяемость генов в мультигенных семействах необходима или для усиления дозы генов (дозовая повторяемость), или для обеспечения синтеза различных вариантов родственных полипептидов (вариантная повторяемость). Дозовая повторяемость характерна для генов, кодирующих тРНК и рРНК, а также гистоновых генов. Вариабельная повторяемость свойственна таким генам, как глобиновые, актиновые, иммуноглобулиновые и многим другим. Наряду с нормально функционирующими генами в мультигенных семействах у эукариот встречаются псевдогены (j-гены), гомологичные функционирующим, но по разным причинам не способные функционировать. Играют ли они какую-либо специальную роль или просто являются "поломанными" генами, не известно, хотя есть косвенные указания на то, что их присутствие нужно для нормального функционирования соседних генов.
Гены прокариот состоят из двух основных элементов: регуляторной части и собственно кодирующей части(рис. 27). Регуляторная часть обеспечивает первые этапы реализации генетической информации, а кодирующая часть содержит информацию о структуре полипептида, тРНК, рРНК. У прокариот структурные гены, кодирующие белки одноо метаболического пути, часто бывают объединены и называются опероном. Так, например, в лактозном опероне E. coli содержится 3 структурных гена. Для биосинтеза аминокислоты гистидин требуется 9 ферментов и ее оперон содержит 9 структурных генов.
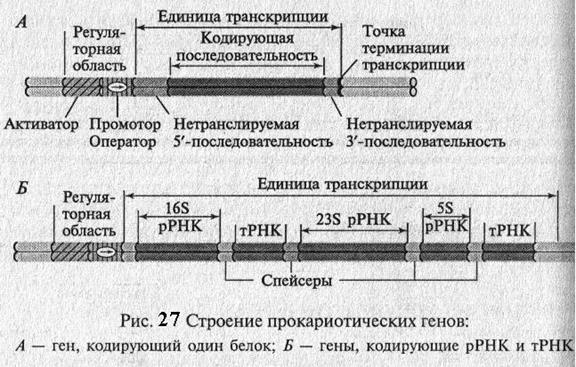
Гены, кодирующие белки, обычно содержат на 5'- и 3'- концах гена или оперона нетранслируемые последовательности (5' – НТП и 3' – НТП), которые играют важную роль в стабилизации иРНК. Гены тРНК и рРНК отделены друг от друга спейсерами (от англ. – spacer – распорка), т.е. последовательностями, которые вырезаются в ходе их созревания (процессинга)(рис. 27).
(А. С. Коничев, Г. А. Севастьянова, 2005, с. 157)
Экзоны – это последовательности участвующие в образовании зрелых РНК. Они могут быть как кодирующие так и некодирующие. Наследственная информация экзонов реализуется в синтезе определенных белков, а роль интронов до конца еще не выяснена.
Возможное значение интронов:
1. Интроны снижают частоту мутаций, соотношение интронов и экзонов у человека 3:2.
2. Интроны поддерживают структуру ДНК, т.е. играют конститутивную роль.
3. Интроны необходимы для процесса созревания иРНК. Без интронов нарушен выход иРНК в цитоплазму. При введение в ядро искусственной иРНК без интронов, она остается в ядре и в цитоплазму не выходит.
4. В последние годы четко установлено, что некоторые интроны кодируют белки – ферменты, которые их вырезают.
5. Превращаются в малые ядерные РНК (мяРНК).

(А. С. Коничев, Г. А. Севастьянова, 2005 г., с. 157)
Гены высших организмов чаще оказываются прерывистыми, но есть и непрерывистые, например, гены интерферонов, гены гистонов. Степень прерывистости может быть различной – от одного интрона как у гена актина до нескольких десятков, как у гена коллагена(рис.29).

Рис. 29. Карты некоторых прерывистых генов. Жирные линии – экзоны, тонкие - интроны (А. С. Коничев, Г. А. Севастьянова, 2005 г., с. 158)
Длина интронов часто оказывается длинней экзонов: 5 – 20 тыс. и 1 тыс. соответственно. Прерывистость гена считалось достоянием эукариот. Но в 1983г. группа ВЕЗЕ (США)обнаружила их у некоторых археобактерий. Интроны содержаться во всех типах РНК, Интроны в составе иРНК вырезаются при участии мяРНП, которые образуют с интроном сплайсосому. При помощи сплайсосом, узнается начало и конец интрона, их концы соединяются в цепи РНК и интрон вырезается (рис.32).
Эволюционное возникновение мозаичной (итрон – экзонной) структуры генов эукариот в настоящее время не находит объяснения. С точки зрения У. Гильберта появление интронов обеспечило возможность обмена экзонами между неродственными генами. В результате это привело к появлению белков с новыми функциями (гипотеза позднего возникновения интронов). По другой гипотезе интроны это эволюционные реликты, они были частью гигантских генов. Прокариоты являются эволюционным тупиком т.к. не содержат интронов.
Регуляция работы гена
Гены функционируют в клетке не сами по себе, а входят в состав более сложной генной регуляторной системы. Количество структурных генов в разных оперонах различно. Участок ДНК, на на котором проходит считывание информации, называется единицей транскрипции (Рис. 27, 28). Он ограничен промотором (зона начала транскрипции) и терминатором (зона остановки транскрипции).
1. Промотор – это строго определенная нуклеотидная последовательность, которая узнается ферментом транскрипции – РНК – полимеразой.
У E. coli промотор – это пара нуклеотидных последовательностей из 6-7 и 9 нуклеотидов каждая, отдельных друг от друга 25 нуклеотидами.
Промотор выполняет следующие функции:
а) это место присоединения РНК – полимеразы к молекуле ДНК
У про- и эукариот последовательности промоторов разные. Это учитывается в генной инженерии, в случае встраивания в геном бактерии генов человека.
Промоторы эукариот разнообразны по числу и строению элементов. Промотор эукариотического гена – это участок ДНК, на котором собираются белки транскрипции, узнающие свои сайты связывания и взаимодействующие друг с другом и с иРНК – полимеразой. В составе эукариотического гена имеются особые цис-действующие элементы регуляции - усилители или активаторы, глушители или угнетатели транскрипции. Они разнообразны по строению, положению и функциям. Они могут располагаться как на 5'- так и на3'- конце фрагмента ДНК, включающего ген, так и в составе интронов.
2. Оператор – это нетранскрибирумая последовательность нуклеотидов, участок связывания белка - репрессора. Он располагается в непосредственной близости к промотору или перекрывается с ним. У многих опероновимеется не один, а несколько сайтов связывания с регуляторными белками, которые не обязательно располагаются рядом, а могут находиться по разные стороны от промотора. Поэтому сейчас принято говорить о сайтах связывания регуляторов. Связывание белка – репрессора с оператором либо создает стерические (пространственные) затруднения для связывания РНК-полимеразы с промотором, либо препятствует продвижению ее по смысловой цепи ДНК, и определяет точку начала транскрипции. Следует отметить, что ни промотор, ни оператор в РНК не транскрибируются и зоны промотора и оператора могут перекрываться.
3. Терминатор – участок молекулы ДНК, где заканчивается процесс транскрипции.
Оператор и структурные гены образуют оперон. Именно так назвали эту структуру французские ученые Франсуа Жакоб и Жак Моно, которые первыми в 1959 -1961г. работая с бактериальными клетками предложили механизм регуляции работы гена или генной экспрессии. За эту работу в 1965г. они получили Нобелевскую премию. Как выяснили Жакоб и Моно работой оперона управляют гены – регуляторы. Они не входят в состав оперона, но являются необходимой частью регуляторной системы. Гены – регуляторы у прокариот находятся на той же хромосоме, что и оперон. У эукариот они могут располагаться далеко от промотора эукариотического гена и оказывать дистанционное влияние на его транскрипцию. Гены - регуляторы контролируют синтез белка – репрессора связывающегося с оператором. Синтез белков – репрессоров, как и всех белков, идет на рибосомах в цитоплазме. Транскрипция определяется белком-репрессором, который может закрывать оператор (репрессор активен) или открывать его (репрессор неактивен), т.е. возможны два варианта регуляции активности генов.
I. Ген – регулятор отвечает за синтез активного белка – репрессора. Белок – репрессор имеет два активных центра:
1. центр связывания с оператором
2. центр связывания с субстратом. Под субстратом (индуктором) понимают любое вещество, информация о синтезе или распаде которого закодирована в данном опероне или гене. Это могут быть гормоны, аминокислоты, углеводы, питательные вещества, яды и т.д.
Субстрата в клетке нет, поэтому активная форма белка – репрессора соединяется с оператором, т.е. оператор закрыт и через него не может пройти фермент РНК – полимераза, транскрипция не идет (рис.30). Открытие оператора идет с помощью субстрата (индуктора), поступающего в клетку. Индуктор взаимодействует с белком – репрессором, что приводит к изменению его конформации (пространственной структуры). У инактивированного белка репрессора резко снижается родство к зоне оператора и он отсоединяется от него. Оператор свободен и это позволяет РНК-полимеразе начать транскрипцию. Она продолжается до тех пор, пока в клетке есть субстрат, т.е. пока есть необходимость в продуктах данного оперона или гена (рис.31).
При сокращении количества субстрата его уже не хватает на молекулы белка – репрессора и активный белок – репрессор присоединяется к оператору. Транскрипция прекращается. Следует отметить, что в клетке белок реперссор синтезируется постоянно и его количество строго определенное. Например, в клетке E. сoli находится около 10 молекул белка-репрессора, который регулирует работу лактозного оперона.
II. Ген – регулятор отвечает за синтез неактивной формы белка – репрессора, т.е. он не может присоединится к оператору. Оператор свободен иРНК – полимераза свободно проходит к структурным генам. Оперон будет работать до тех пор, пока есть необходимость в продуктах данного оперона.
Когда данный продукт клетке уже не нужен (он не расходуется в биохимических процессах и накапливается в клетке), субстрат взаимодействует с неактивным белком – репрессором, активирует его. Белок-репрессор закрывает оператор и выключает транскрипцию.
Такой способ регуляции метаболизма в клетке чрезвычайно экономичен, т.к. клетка синтезирует продукт в таком количестве, которое необходимо для поддержания определенного уровня обменных процессов. При избытке конечного продукта данный метаболический путь выключается. То есть, мы видим взаимодействие между внутриклеточной средой и генетическим аппаратом для обеспечения тонкой регуляции клеточного метаболизма.
Например, образование некоторых ферментов индуцируется присутствием их субстрата:
1. наличие в крови алкоголя индуцирует в клетках печени усиленный синтез фермента, разрушающего алкоголь – алкогольдегидрогеназы.
2. действие половых гормонов при формировании вторичных половых признаков также основано на усилении транскрипции определенных генов.
3. по такому типу работают гены железистых клеток, вырабатывающие секреты для жизнедеятельности организма.
Если у бактерии на включение гена в работу требуется несколько минут, то у эукариот от нескольких часов до нескольких дней.
Включение и работа генов и оперонов зависят от ряда факторов:
1. Специализации клетки
2. Физиологического состояния
3. Возраста клетки
4. Условий внешней среды
5. Пространственной структуры ДНК (изгибы, петли, сверхспирали и т.д.)
6. Степени метилирования генов.
Показано, что гены материнских и отцовских хромомсом могут быть метилированы по – разному и это регулирует активность разных генов. Например, ген – индуцирующий образование опухолей. Если он передается потомству от отца, то транскрибируется только в сердце, а если от матери, то он вообще не экспрессируется. Исследования показали, что у самок этот ген метилирован, а у самцов – деметилирован.
Любой из этих факторов может оказать существенное влияние на процесс считывания генетической информации.
Читайте также: