
Болезни репарации днк реферат
Обновлено: 04.07.2024
Репарация — клеточный механизм коррекции повреждённой последовательности ДНК (точечные мутации, делеции, структурные нарушения и др.).
Системы репарации
• Темновая (фотореактивация): удаляются УФ-индуцированные ковалентные связи между смежными основаниями ДНК.
• Рекомбинационная: для восстановления одной хромосомы используется материал гомолога. Дефекты рекомбинационной репарации регистрируют при синдроме Блума.
Гены, участвующие в репарации ДНК • 126380, ген ERCC1, 19q13.2–19q13.3. До настоящего времени его клиническое значение неясно; совместно с белками XPA и XPF (продукт гена ERCC4) участвует в процессах распознавания и вырезания дефектного участка ДНК • 126340, ген ERCC2, 19q13.2–19q13. 3. Кодирует геликазу с неясными функциями; мутации вызывают пигментную ксеродерму типа D • 133510, ген ERCC3, 2q21. Кодирует геликазу репарации ДНК; различные мутации вызывают пигментную ксеродерму типа B, комплекс пигментной ксеродермы и синдрома Коккейна, трихотиодистрофию • 133530, ген ERCC5, 13q32–13q32, 13q33–13q33. Мутации вызывают комплекс пигментной ксеродермы и синдрома Коккейна типа А • 133540, ген ERCC6 (CKN2), 10q11–10q21 . Мутации вызывают синдром Коккейна типа В.
Код вставки на сайт
Репарация — клеточный механизм коррекции повреждённой последовательности ДНК (точечные мутации, делеции, структурные нарушения и др.).
Системы репарации
• Темновая (фотореактивация): удаляются УФ-индуцированные ковалентные связи между смежными основаниями ДНК.
• Рекомбинационная: для восстановления одной хромосомы используется материал гомолога. Дефекты рекомбинационной репарации регистрируют при синдроме Блума.
Гены, участвующие в репарации ДНК • 126380, ген ERCC1, 19q13.2–19q13.3. До настоящего времени его клиническое значение неясно; совместно с белками XPA и XPF (продукт гена ERCC4) участвует в процессах распознавания и вырезания дефектного участка ДНК • 126340, ген ERCC2, 19q13.2–19q13. 3. Кодирует геликазу с неясными функциями; мутации вызывают пигментную ксеродерму типа D • 133510, ген ERCC3, 2q21. Кодирует геликазу репарации ДНК; различные мутации вызывают пигментную ксеродерму типа B, комплекс пигментной ксеродермы и синдрома Коккейна, трихотиодистрофию • 133530, ген ERCC5, 13q32–13q32, 13q33–13q33. Мутации вызывают комплекс пигментной ксеродермы и синдрома Коккейна типа А • 133540, ген ERCC6 (CKN2), 10q11–10q21 . Мутации вызывают синдром Коккейна типа В.
студент группы ОМ-109 Б
Брюшинина Евгения
Проверила:
Устройство системы репарации……………………………………………………… …………. 4
Принципы репарации ДНК у различных организмов сходны, поэтому эти принципы рассматриваются на примере E. coli, у которой они хорошо изучены.
Репарация — особая функция клеток, заключающаяся в способности исправлять химические повреждения и разрывы в молекулах ДНК, повреждённой при нормальном биосинтезе ДНК в клетке или в результате воздействия физическими или химическими агентами. Осуществляется специальными ферментными системами клетки.
Начало изучению репарации было положено работами А. Келнера (США), который в 1948 обнаружил явление фотореактивации (ФР) — уменьшение повреждения биологических объектов, вызываемого ультрафиолетовыми (УФ) лучами, при последующем воздействии ярким видимым светом (световая репарация).
Р. Сетлоу, К. Руперт (США) и др. вскоре установили, что фотореактивация — фотохимический процесс, протекающий с участием специального фермента и приводящий к расщеплению димеров тимина, образовавшихся в ДНК при поглощении УФ-кванта.
Позднее при изучении генетического контроля чувствительности бактерий к УФ-свету и ионизирующим излучениям была обнаружена темновая репарация — свойство клеток ликвидировать повреждения в ДНК без участия видимого света. Механизм темновой репарации облученных УФ-светом бактериальных клетокбыл предсказан А. П. Говард-Фландерсом и экспериментально подтвержден в 1964 Ф. Ханавальтом и Д. Петиджоном (США). Было показано, что у бактерий после облучения происходит вырезание поврежденных участков ДНК с измененными нуклеотидами и ресинтез ДНК в образовавшихся пробелах.
Системы репарации существуют не только у микроорганизмов, но также в клетках животных и человека, у которых они изучаются на культурах тканей. Известен наследственный недуг человека — пигментная ксеродерма, при котором нарушена репарация.
Источники повреждения ДНК
* Ошибки репликации ДНК
* Апуринизация — отщепление азотистых оснований от сахарофосфатного остова
* Дезаминирование — отщепление аминогруппы от азотистого основания
Основные типы повреждений ДНК
* Ошибки репликации ДНК
* Апуринизация — отщепление азотистых оснований от сахарофосфатного остова
* Дезаминирование — отщепление аминогруппы от азотистого основания
Устройство системы репарации
Каждая из систем репарации включает следующие компоненты:
* фермент, "узнающий" химически изменённые участки в цепи ДНК и осуществляющий разрыв цепи вблизи от повреждения;
* фермент, удаляющий повреждённый участок
* фермент (ДНК-полимераза), синтезирующий соответствующий участок цепи ДНК взамен удалённого;
* фермент (ДНК-лигаза), замыкающий последнюю связь в полимерной цепи и тем самым восстанавливающий её непрерывность (см. рис. справа).
По отношению к процессу репликации различают два типа репарации ДНК:
1. Дорепликативную (включающую фотореактивационную и эксцизионную формы, направленные на вырезание поврежденных участков ДНК);
2. Пострепликативную (осуществляемую с помощью механизмов, участвующих в процессах рекомбинации и репликации ДНК).
Репарация может осуществляться как конститутивно с помощью специфического набора ферментов, постоянно присутствующих в нормально функционирующих клетках (фотореактивационная, эксцизионная и пострепликативная), так и в ответ на повреждение ДНК или прекращение ее синтеза (путем активации группы генов, контролирующих различные клеточные функции, так называемая SOS-репарация).
У бактерий имеются по крайней мере 2 ферментные системы, ведущие репарацию — прямая и эксцизионная.
Типы репарации ДНК:
* Прямое исправление повреждений
Наиболее частая причина точечных мутаций у человека: это спонтанное добавление метильной группы — один из типов алкилирования. Такие модификации исправляются без разрушения цепи ДНК ферментами — гликозилазами. Фермент О-6-метилгуанин-ДНК- метилтрансфераз (MGMT) защищает клетку от токсических эффектов, производимых алкилирующими агентами, переводя для этого метильную группу из О-6-метилгуанин-ДНК в цистеиновый остаток в MGMT.
* Починка путем вырезания основания ( эксцизионной репарации основания — BER)
Главные способы, посредством которых происходит починка оснований ДНК, включают устранение поврежденного основания, которое осуществляют ферменты нуклеазы. Возникающая лакуна может быть заполнена ДНК-полимеразой, что сопровождается лигацией с родительской ДНК. Окислительные повреждения, как основные, так и индуцированные, являются важными причинами для починки оснований ДНК.
* Починка путем вырезания нуклеотида (эксцизионной репарации нуклеотида — NER)
Система NER обеспечивает способность клетки устранять объемные повреждения в ДНК. NER удаляет содержащий повреждение олигонуклеотид из ДНК посредством распознавания повреждения, разреза, вырезания, нового повторного синтеза и лигации. ДНК-полимеразы (у эукариот их известно более 15) различаются по ряду признаков, в том числе по количеству нуклеотидов, которые они могут встроить в растущую цепь за один акт связывания с дуплексом ДНК. присоединяют один нуклеотид, а при участии белка l и bДНК-полимеразы —e иdPCNA (proliferating cell nuclear antigen) ДНК-полимеразы способны вставить фрагмент необходимой длины. Повреждения ДНК в активно транскрибирующих генах, особенно в транскрибирующей спирали, чинятся в первую очередь и потому быстрее, чем повреждения ДНК в остальной части генома. Так клетка защищает целостность процесса транскрипции.
* Репарация ошибок спаривания — мисмэтч репарация (mismatch — MMR)
Этот метод исправляет ошибочно встроенные неповрежденные основания, которые не образуют нормальное Уотсон-Криковское спаривание (A • T, C • G). Такие ошибки иногда происходят при репликации с помощью ДНК- полимеразы. В MMR участвуют ферменты, вовлеченные в BER и NER, а также специализированные ферменты. Синтез ДНК при MMR осуществляется ДНК-полимеразами
У эукариот существует два основных способа устранить двухцепочечные разрывы: гомологичная рекомбинация (рекомбинационная репарация) и соединение негомологичных концов. Прямое соединение сломанных концов требует специальных ферментов, которые узнают и связывают разорванные концы с последующим их сшиванием. Починка разрывов двойной спирали (и разрывов одной спирали) обычно включает в себя продуцирование трехконечного односпирального хвоста с помощью экзонуклеаз или геликаз. Посредством инвазии спирали, при которой односпиральный хвост вторгается в неповрежденную гомологичную молекулу ДНК, синтезируется ДНК. Одновременно образуется так называемое холлидеевское сочленение в комплексе ДНК. Через промежуток этого сочленения проводятся две молекулы ДНК (как со структуральным перекрестом, так и без него), каждая из которых более не содержит разрывов.
* Негомологичное соединение концов—NHEJ (Nonhomologous End-Joining)
Если разорванная ДНК имеет тупые концы, и соединение двух фрагментов ДНК происходит случайно, то такая репарация называется NHEJ. Главным компонентом NHEJ восстановительного комплекса является зависящий от ДНК белок киназа (DNA-PK), или белок Ku — гетеродимерная субъединица, состоящая из двух белков Ku70 и Ku80. Этот белок служит для выравнивания концов разорванной ДНК, чтобы упростить процесс их склеивания или выступает в качестве сигнальной молекулы для мобилизации других восстанавливающих белков.
Рассмотрим подробнее некоторые из вышеперечисленных типов.
Прямая репарация ДНК
Прямая репарация ДНК обеспечивает прямое восстановление исходной структуры ДНК или удаление повреждения. Широко распространенная система репарации такого рода — фотореактивация пиримидиновых димеров. Кроме нее, к этому типу относятся: репарация ДНК за счет 3'-5'-экзонуклеазной активности ДНК-полимеразы, репарация одноцепочечных разрывов ДНК с помощью полинуклеотиллигазы, а также генетическая репарация повреждений, вызванных алкильными или метильными группами, путем удаления этих групп специфическими ферментами.
В 1949 г. А. Кельнер и в 1950 г. Р. Дульбекко установили, что жизнеспособность актиномицетов и бактерий, подвергнутых УФ-облучению в летальных дозах, восстанавливается, если затем воздействовать на них видимым светом. Явление было названо фотореактивацией. Эффективность ее зависит от уровня рН, температуры и физиологического состояния клетки. Восстановительный эффект при фотореактивации (рис.) связан с действием фермента — дезоксирибозидпиримидинфотолиа зы, представляющего собой полипептид, ассоциированный для его активности с небольшой молекулой РНК (10-15 нуклеотидов).
Этот фермент расщепляет димеры двух соседних пиримидинов циклобутанового типа в одной цепи ДНК, образующиеся под влиянием УФ-лучей, действие которых подробнее рассмотрено в нашей статье. Каждый из димеров задерживает репликацию примерно на 10 секунд. Фермент присоединяется к ним и в темноте, и на свету, но реакция расщепления связей, объединяющих две молекулы пиримидинов, энергетически зависит от действия видимого света с большей длиной волны. На свету пиримидиновые димеры расщепляются, за счет разрыва ковалентных связей происходит мономеризация и таким образом восстанавливается нативная структура ДНК. К эффективному диапазону (365-490 нм) относятся наиболее длинноволновые УФ-лучи (365-390 нм) и примыкающие к ним видимые синие лучи (435—495 нм). Наибольшая эффективность фотореактивации отмечена для голубой части видимого спектра. Если же необходимо исключить возможность реактивации, то опыты следует проводить в более длинноволновой части спектра, начиная с желтого света (570-590 нм).
За 1 минуту молекула фотолиазы может расщепить 2,4 димера. У Е. coli система фотореактивации удаляет до 90% пиримидиновых димеров и контролируется одним геном - phr. Штаммы, несущие мутацию по этому гену, не способны к репарации ДНК.
Содержание
Введение
1. Понятие репарации
2.1История открытия
2.2 Источники повреждения ДНК
2.3 Основные типы повреждений
2.4 Устройство системы репарации
2. Типы репарации
3.5 Прямая репарация
2.1.1 Фотореактивация
3.6 Эксцизионная репарация
3.7 Мисмэтч-репарация
3.8 SOS-репарация
3. Интересные факты
Использованные источники
1. ПОНЯТИЕРЕПАРАЦИИ
Репарация — особая функция клеток, заключающаяся в способности исправлять химические повреждения и разрывы в молекулах ДНК, повреждённой при нормальном биосинтезе ДНК в клетке или в результате воздействия физическими или химическими агентами. Осуществляется специальными ферментными системами клетки.
1.1 История открытия
Начало изучению репарации было положено работами А. Келнера (США),который в 1948 обнаружил явление фотореактивации (ФР) — уменьшение повреждения биологических объектов, вызываемого ультрафиолетовыми (УФ) лучами, при последующем воздействии ярким видимым светом (световая репарация).
Р. Сетлоу, К. Руперт (США) и др. вскоре установили, что фотореактивация — фотохимический процесс, протекающий с участием специального фермента и приводящий к расщеплению димеров тимина,образовавшихся в ДНК при поглощении УФ-кванта.
Позднее при изучении генетического контроля чувствительности бактерий к УФ-свету и ионизирующим излучениям была обнаружена темновая репарация — свойство клеток ликвидировать повреждения в ДНК без участия видимого света. Механизм темновой репарации облученных УФ-светом бактериальных клетокбыл предсказан А. П. Говард-Фландерсом иэкспериментально подтвержден в 1964 Ф. Ханавальтом и Д. Петиджоном (США). Было показано, что у бактерий после облучения происходит вырезание поврежденных участков ДНК с измененными нуклеотидами и ресинтез ДНК в образовавшихся пробелах.
Системы репарации существуют не только у микроорганизмов, но также в клетках животных и человека, у которых они изучаются на культурах тканей. Известен наследственный недугчеловека — пигментная ксеродерма, при котором нарушена репарация.
2.2 Источники повреждения ДНК
* УФ излучение
* Радиация
* Химические вещества
* Ошибки репликации ДНК
* Апуринизация — отщепление азотистых оснований от сахарофосфатного остова
* Дезаминирование — отщепление аминогруппы от азотистого основания.
Основные положения:
• Под влиянием факторов внешней среды или в результате ошибок в работе различных систем клетки в генетическом материале постоянно возникают повреждения
• Для сохранения жизнеспособности во всех клетках должны быть системы репарации, снижающие количество повреждений в ДНК
Наряду с безошибочным воспроизведением генетической информации важную роль играет поддержание ее информационной целостности. Фактически в геноме человека присутствует больше генов, ответственных за репарацию повреждений ДНК, чем кодирующих ферменты репликации.
Ошибки в последовательностях ДНК могут возникать по двум причинам. Во-первых, при репликации, во вновь образующуюся цепь ДНК может включиться неправильное основание. Для предотвращения таких ошибок в системе репликации существует корректорский механизм, который снижает число ошибочно включенных нуклеотидных остатков до минимума.
Во-вторых, при воздействии таких факторов внешней среды, как ионизирующие излучения и химические агенты, нарушающие структуру нуклеотидов, в ДНК могут возникнуть повреждения. В клетке существует много репаративных систем, которые устраняют повреждения в последовательностях ДНК, восстанавливая их правильную структуру.
Рисунок иллюстрирует действие репаративной системы, которая узнает повреждения в ДНК, удаляет их, и восстанавливает исходную структуру.
Несмотря на функционирование систем репарации, в генетическом материале возникают мутации, однако они не нарушают жизнедеятельность клетки. В действительности, определенная частота мутаций необходима для обеспечения вариабельности организмов в процессе эволюции.
Мутации во всех организмах, от бактерий до высших эукариот, возникают с частотой порядка 10 -6 в пересчете на один ген (или 10 -9 -10 -10 в пересчете на нуклеотидную пару) за одно поколение. Примерно такое же количество мутаций возникает даже у организмов, живущих в экстремальных условиях. Это позволяет предполагать, что общая частота возникновения мутаций определяется балансом между неблагоприятным эффектом большинства вредных мутаций и некоторыми полезными мутациями.
Ни одна клетка не может существовать в отсутствие систем репарации. Если, например, у Е. coli прекратить действие всех систем репарации, то однократное облучение бактерий УФ может оказаться летальным. В то же время бактерии с функционирующими системами репарации выносят огромное количество повреждений.
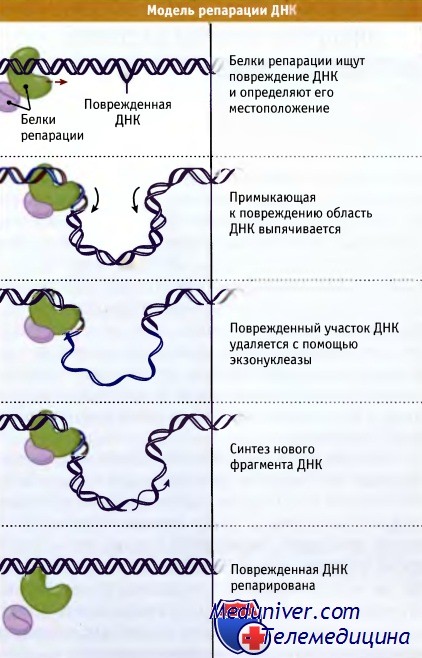
Система репарации узнает повреждение в ДНК,
удаляет поврежденный участок и заполняет образующуюся брешь.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Читайте также: