
Антибиотики пептидные антибиотики реферат
Обновлено: 05.07.2024
ПЕПТИДНЫЕ АНТИБИОТИКИ, антимикробные соед , в молекулах к-рых имеются пептидные связи. В хим. отношении это весьма разнородная группа в-в, большинство из к-рых-циклич. или линейные олиго- и полипептиды, содержащие заместители непептидной природы (остатки жирных к-т, алифатич. аминов и спиртов, гидроксикислот, а также Сахаров и гетероциклов).
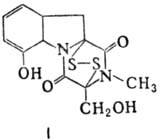
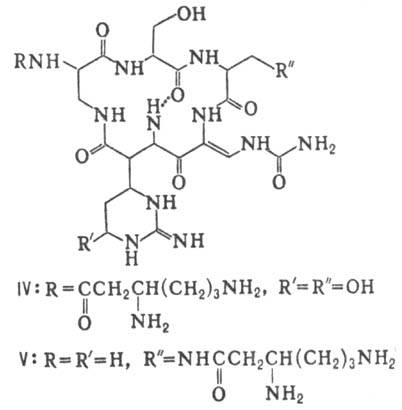
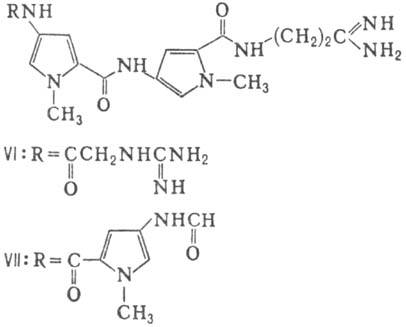
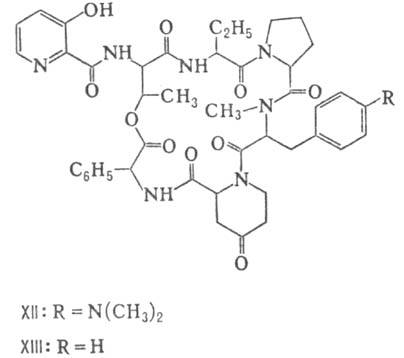
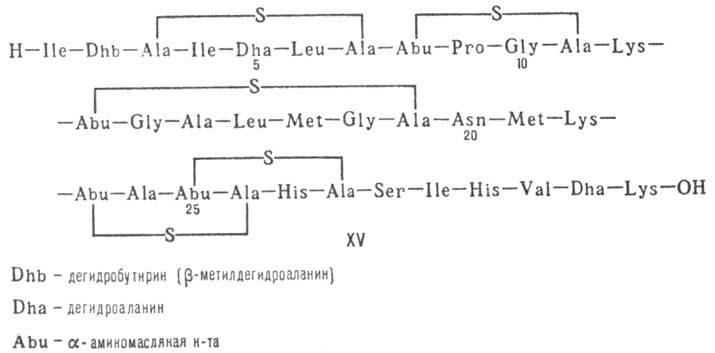
Различают пять осн. видов пептидных антибиотиков: 1) производные аминокислот (напр., циклосерин, b -лактамные антибиотики)и дикетопиперазина (глиотоксин, ф-ла I); 2) гомомерные пеп-тиды-линейные (грамицидин А, II) и циклические [бацит-рацин А, III (здесь и ниже буквы греч. алфавита показывают положение аминогрупп, к-рые участвуют в образовании связей); виомицин,, IV; капреомицин 1-А, V], а также олиго-пептиды (нетропсин, VI; дистамицин, VII); 3) гетеромерные пептиды [напр., полимиксины В, E и M, ф-лы соотв. VIII, IX и X; R = 6-метилоктаноил (B 1 , E 1 и M 1 ) или изооктаноил (B 2 , E 2 и M 2 ); Dab -2,4-диаминомасляная к-та], в т.ч. хелато-образующие (блеомицины); 4) пептолиды-хромопептолиды (актиномицины), липопептолиды (стендомицин, XI; здесь и ниже буквы Me перед лат. обозначениями аминокислот, кроме Pro, указывают на наличие в них метильной группы у атома N; МеРго 4-метилпролин), гетеропептолиды (мика-мицин В, XII; стафиломицин S, XIII), простые пептолиды (гризелимицин A, XIV) и депсипептиды (валиномицин; см. Ионофоры); 5) макромолекулярные пептиды полипептиды (низин, XV; сульфидные мостики связывают b -С-атомы Ala и Abu), белки (неокарциноста-тин, содержащий 109 аминокислотных остатков), протеиды (аспарагиназа, лизостафнин с мол. м. 32000).
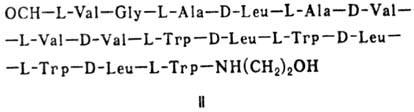
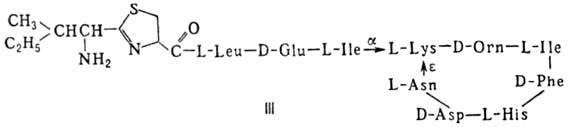
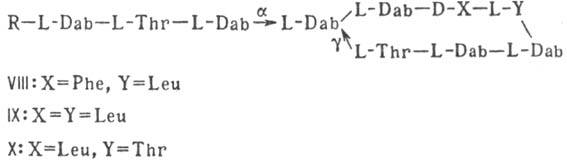


Гомо- и гетеромерные пептиды, пептолиды имеют ряд характерных особенностей, к-рые отличают их от обычных полипептидов и белков: а) низкое содержание нек-рых простых аминокислот (аргинин, гистидин, метионин), наличие аминокислот D-конфигурации и аминокислот необычной структуры (серосодержа-щих, сложных гетероциклич., ненасыщенных, N-метилированных, имино-, b - и g -аминокис-лот, производных пролина); б) наличие в составе молекул заместителей непептидной природы; в) преим. циклич. или линейно-циклич. структура без своб. карбокси- и аминогрупп; циклизация между собств. аминокислотными радикалами с образованием тиазолинов, окса-золинов и др. гетероциклич. структур. Кроме того, пептидные антибиотики, как правило, устойчивы к действию гидролаз, хотя нек-рые из них (полимиксины, блеомицины) чувствительны к аминоацилазам и пептидазам микробного и растит. происхождения.
П ептидные антибиотики продуцируются в виде смеси родственных соед., отличающихся друг от друга одним или неск. аминокислотными остатками или вариациями в строении компонентов непептидной природы. Продуцентами являются разл. виды актиномицетов, бактерий и грибов. Биосинтез пептидов и депсипептидов осуществляется без участия рибосом и РНК с помощью специфич. ферментных комплексов-синтетаз антибиотиков, содержащих всю необходимую информацию. Для ряда пептидных антибиотиков выяснен мол. механизм биосинтеза или установлен состав синтетаз. В процессе полимеризации или после образования пептидной цепи происходит циклизация молекулы и модификация отдельных аминокислот. Биосинтез макромолекулярных пептидных антибиотиков (в частности, низина) происходит на рибосомах с послед. модификацией белка-предшественника.
П ептидные антибиотики имеют разнообразные биол. св-ва. Среди.них встречаются ингибиторы синтеза клеточной стенки (бацитрацин А) и синтеза липопротеидов наружной мембраны грамотри-цат, бактерий (бицикломицин), ингибиторы репликации и транскрипции (актиномицин D, блеомицины) и синтеза белка (виомицин), ингибиторы функционирования клеточной мембраны (полимиксины, грамицидин, валиномицин), антиметаболиты (аланозин, циклосерин). Пептидные антибиотики обладают высокой антибиотич. активностью в отношении грамполо-жит. (бацитрацин А) и грамотрицат. (полимиксины) бактерий, а также микобактерий (капреомицин 1-А, виомицин). Ряд антибиотиков проявляют противоопухолевую (актино-мицины, аспарагиназа) и противогрибковую активность; дистамицин весьма активен в отношении вирусов.
П ептидные антибиотики широко применяют в ветеринарии (микамицин В, нетропсин), в качестве кормовых добавок (бацитрацин А, стафиломицины), как консерванты (низин), в биохим. исследованиях (валиномицин, грамицидины, актиномицины). Использование пептидных антибиотиков в терапии довольно ограниченно из-за нежелат. побочных эффектов, в частности нефроток-сичности. Широко применяют лишь полимиксины В, E и M, нек-рые противоопухолевые (блеомицин A 2 , актиномицин D, аспарагиназа) и противотуберкулезные препараты (циклосерин, виомицин, капреомицин 1-А, лизостафнин). Пептидные антибиотики, однако, вытесняются из мед. практики менее токсичными антибиотиками.
Лит. Ланчини Д.. Паренти Ф., Антибиотики, пер. с англ., M., 1985; Антибиотики-полипептиды (структура, функция, биосинтез), под ред. H. С. Егорова, M., 1987; Bioactive peptides produced by microorganisms, ed. by H. Umezawa, T. Takita, T. Shiba, Kodansha, Tokyo, 1978; Berdy J., CRC Handbook of antibiotic compounds, v. 4, pt 1 2, Boca Raton, 1980. А. А. Зинчепко.
Пептиды, осуществляющие защиту животных от инфекции. Микроорганизмы как биотический фактор среды, ответственный за биосинтез антимикробных пептидов. Влияние дисульфидных связей на функционирование молекул полипептидов в качестве антибиотических агентов.
| Рубрика | Биология и естествознание |
| Вид | реферат |
| Язык | русский |
| Дата добавления | 06.09.2009 |
| Размер файла | 57,0 K |
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
"Пептидные антибиотики животных как биохимические факторы противоинфекционной защиты"
Пептиды, осуществляющие защиту животных от инфекции, широко представлены в природе от простейших до человека. Наиболее распространенным в эволюции типом антимикробных молекул являются катионные цистинсодержащие полипептиды. Наименьшим по размерам молекулы представителем этой группы антибиотических соединений является додекапептид с 1 дисульфидной связью, выделенный из нейтрофилов коров. Открытые нами протегрины содержат уже 2 дисульфидных мостика, так же как и подобные им соединения из гемоцитов подковообразного краба - тахиплезины и гемолимфы скорпиона - андроктонин. Дефенсины человека, кролика, крысы, мыши и морской свинки, как и р-дефенсины коров и галлинацины кур, содержат в своем составе 6 цистеиновых остатков, образующих 3 внутримолекулярные S-S-связи. Замыкают рассматриваемый ряд антимикробные полипептиды eNAP-1 и eNAP-2 из нейтрофилов лошади, для структуры которых характерно наличие уже 4 дисульфидных групп. Есть основания предполагать, что дисульфидные связи придают молекулам полипептидов повышенную устойчивость к переваривающему действию многочисленных протеиназ лейкоцитарного и микробного происхождения, обеспечивая их пролонгированное функционирование в качестве антибиотических агентов при фагоцитозе и воспалении.
Следует подчеркнуть, что фагоциты и гемоциты являются не единственными клетками организма, содержащими дефенсины и структурно-родственные им полипептиды. Выявлены дефенсины в клетках эпителия тонкой кишки мышей, кролика и человека, в эпителии трахеи и языка коров, в структурах мочеполового и репродуктивного трактов человека.
Присутствие дефенсинов в клетках слизистой тонкого кишечника интересно в связи с представлениями И.И. Мечникова о морфофизиологическкх предпосылках формирования фагоцитарной функции клеток в эволюции. Он рассматривал фагоцитоз как антимикробную функцию специализированных клеток организма, возникшую и развившуюся на основе пищеварительной активности цитоплазмы. Антимикробная функция фагоцитов эволюционно возникла на основе способности простейших поглощать и инактивировать бактерии, которые являются основным объектом их питания. Поэтому уже на заре развития животных перед ними стояла задача инактивации потенциально патогенных микроорганизмов, являющихся для них источником пластических веществ и энергии. Есть основания допускать, что продукция простейшим Entamoeba histolytica пептидов, структурно гомологичных NK-лизинам лимфоцитов, отражает генеральную линию развития механизмов противоинфекционной резистентности у животных. В последние годы дефенсины описаны и у высших растений, причем, несмотря на наличие у них 4 дисульфидных связей, они имеют вторично-третичную структуру, сходную с таковой дефенсинов насекомых. Данные о единстве ряда молекулярных механизмов защиты от инфекции у животных и растений могут служить свидетельством общности их происхождения в эволюции.
Локализация дефенсинов в фагоцитах и клеточно-тканевых структурах организма, "пограничных к инфекции", свидетельствует в пользу их участия в формировании неспецифической антимикробной резистентности в качестве универсальных антибиотиков эндогенного происхождения. Более того, рассматриваемый молекулярный механизм защиты от микробов является, по-видимому, одним из древнейших в эволюции животных, поскольку полипептиды дефенсиновой природы встречаются уже в гемолимфе имаго медоносной пчелы, личинок мясной мухи, жука Zophobas atratus и стрекозы.
Другой распространенной в эволюции матрицей антибиотических пептидов являются цекропины, выявленные первоначально в гемолимфе ряда насекомых, а позже в слизистой тонкого кишечника свиньи. Гены структурно-родственных им пептидов были клонированы и секвеированы из асцидии Styela clava в лаборатории проф. Лерера. Эта гомологическая группа пептидов представляет собой линейные, не содержащие цистеина молекулы оснбвной природы.
При всем структурном разнообразии большинства описанных в настоящее время антибиотических пептидов все они являются, как правило, одновременно основными и амфипатическими молекулами, проявляющими сродство как к липофобным, так и липофильным средам и соединениям. Положительный заряд и амфифильность молекул антибиотических пептидов лежат в основе их функциональных проявлений,' в частности антимикробного действия. Благодаря им антибиотические пептиды вступают в электростатическое и гидрофобное взаимодействия с анионными фосфолипидами и липополисахаридами мембран микробных клеток-мишеней, которые приводят сначала к их адсорбции на поверхности мембран, а потом к внедрению в двойной липидный слой, что нарушает организацию и целостность оболо-чечных структур микроорганизмов. Подобное воздействие АП на ци-топлазматическую мембрану имеет следствием необратимые повреждения ее структуры и нарушения ее разнообразных функций, результирующим эффектом которых является гибель клеток-мишеней. Частным случаем цитотоксичности АП является их действие на микроорганизмы. В силу мембранотропного механизма антибиотического действия этих пептидов они способны в определенных условиях проявлять повреждающие эффекты в отношении клеток животного организма, который их продуцирует. В связи с этим встает вопрос о том, какие клеточно-молекулярные механизмы обеспечивают прицельность именно антимикробного действия АП, сводя к минимуму их возможные аутоповреждающие эффекты в процессах фагоцитоза и воспаления, а также на поверхности барьерных эпителиев макроорганизма. Как теперь установлено, подобная избирательность действия АП определяется рядом структурных и морфологических факторов. К последним относится, в частности, компартментализация дефенсинов в лизосомоподобных гранулах лейкоцитов и клеток Панета, где они связаны с кислыми мукополисахаридами, которые обеспечивают нейтрализацию цитотоксических молекул. Другие пептиды упакованы в гранулах в форме функционально неактивных предшественников, которые подвергаются ограниченному протеолизу с освобождением антимикробного пептида только в случае активации нейтрофильных гранулоцитов в процессе фагоцитоза. В случае освобождения содержимого лизосомоподобных гранул нейтрофильных гранулоцитов во внеклеточное пространство цитотоксичность дефенсинов нейтрализуется вследствие их взаимодействия с плазменными белками, являющимися в своей основной массе анионными соединениями. В настоящее время установлено, что хорошо известные ингибиторы сериновых протеиназ - а2-макроглобулин, а 1-ингибитор протеиназ, а 1 - антихимотрипсин, антитромбин Ш - обладают повышенным сродством к дефенси-нам, благодаря чему образуются нецитотоксичные комплексы серпины-дефенсины.
Существенно важным фактором, определяющим избирательность действия АП на микроорганизмы, являются особенности поверхностных структур бактерий, низших грибов, оболочечных вирусов и, в меньшей степени, простейших. В клеточной стенке и цитоплазматической мембране бактерий и грибов локализованы молекулы, к которым АП проявляют повышенное сродство в ходе межмолекулярных электростатических и гидрофобных взаимодействий. Так, в частности, в состав липидов цитоплазматической мембраны большинства бактерий входят кислые фосфолипиды в существенно более высокой концентрации, чем в плазмалемму эукариотических клеток. Вследствие повышенного электростатического взаимодействия АП с мембранными структурами, обогащенными кислыми фосфолипидами, наблюдается их преимущественная сорбция на бактериальных клетках как в фаголизосомах фагоцитов, так и во внеклеточной среде в очагах воспаления и на поверхности слизистых и кожных покровов. Благодаря ионному взаимодействию АП концентрируются в областях цитоплазматических мембран бактерий, обогащенных кислыми фосфолипидами. Подобное свойство АП было неоднократно продемонстрировано в модельных условиях с использованием искусственных плоских мембран и липосом. Повреждающее действие АП на мембраны-мишени также зависит и от интенсивности гидрофобных взаимодействий пептида с углеводородными хвостами жирных кислот липидов. Амфипатический характер строения молекул большинства АП обеспечивает их внедрение в двойной липидный слой мембран и нарушение структурной целостности последних. Известно также, что присутствие холестерина в составе липидов мембран эукариотических клеток повышает резистентность последних к повреждающему действию таких пептидов, как магейнин и цекропин.
При анализе рассматриваемых взаимодействий важно также учитывать то обстоятельство, что цитоплазматическая мембрана бактерий непосредственно недоступна для контакта с АП, поскольку покрыта клеточной стенкой, в состав которой у грамположительных бактерий входит пептидогликан, а грамотрицательных - пептидогликан и наружная мембрана, являющаяся дополнительным защитным барьером микробной клетки. Поэтому дефенсины, за исключением некоторых криптдинов, и мягейнины более эффективно поражают in vitro грамположительную микрофлору, нежели грамотрицательную. Однако это правило распространяется не на все группы антибиотических пептидов. Например, цекропины и некоторые бактенецины более активны как цитотоксические молекулы в отношении грамотрицательных бактерий. Это свойство указанных соединений обусловлено их повышенной способностью к взаимодействию с одним из типичных представителей структуры наружной мембраны грамотрицательных бактерий, каковым является липополисахарид. Наличие липополисахарида во внешнем слое наружной мембраны грамотрицательных бактерий является одним из условий их опознания антибиотическими пептидами и белками.
Пенетрирующая липофильный бислой активность молекул антибиотических пептидов в ряде случаев заметно зависит от трансмембранного электрического потенциала клеток-мишеней, который, как известно, у плазмалеммы бактерий обычно в 1.5-2 раза более высокий, чем у мембран эукариотических клеток. Это свойство мембран является одним из условий, облегчающих проникновение АП через липидный бислой, особенно в тех случаях, когда эффекторные молекулы умеренно - или слабоосновные. Проникающая способность высокоосновных пептидов, таких как дефенсины кролика NP-1 и NP-2, практически не зависит от мембранного потенциала клеток-мишеней. Направление электрического поля поперек мембран от плюса на внешней поверхности мембраны к минусу - на внутренней обеспечивает электрофорез положительно заряженных молекул антибиотических пептидов через мембрану внутрь клетки. При этом часть молекул самостоятельно или в ассоциации друг с другом внедряется в мембраны, образуя в них пороподобные отверстия различной молекулярной организации. Перфорация мембран, которая в ряде случаев носит транзиторный характер, приводит к утечке из клеток ионов и нарушению ионной асимметрии между средой и клетками. Следствием этого является диссипация мембранного потенциала клеток-мишеней и стремление молекул воды войти в клетки. Результатами подобного движения воды могут быть разбухание клеток-мишеней и их осмотический лизис. Рассеивание мембранного потенциала лишает клетки возможности осуществлять активный транспорт ионов и веществ против градиента концентраций, что в итоге резко снижает их жизнеспособность.
При типичной ориентации электрического поля мембран катионные АП могут переноситься через этот барьер путем электрофореза, подобно тому как это описано для микробных пептидных антибиотиков полимиксина В и грамицидинов., что дополняет картину деструктурирующих воздействий наv клетки-мишени, результатом которых является гибель последних. Обобщенная схема предполагаемого механизма антимикробного действия одного из антибиотических пептидов - цекропина - представлена на рис. 20. Первая фаза действия пептида связана с адсорбцией димера цекропина на поверхности мембраны за счет электростатических взаимодействий с ее отрицательно заряженными соединениями. На следующей стадии происходит диссоциация димера и внедрение в липофильную фазу липидного бислоя отдельных молекул цекропина за счет гидрофобных взаимодействий. На заключительной стадии внедрившиеся и ориентированные поперек мембран молекулы цекропинов формируют каналоподобные поры с умеренной селективной проницаемостью для анионов.
Схема, отражающая основные этапы взаимодействия цекропинов с липидной мембраной.
Таким образом, отрицательный заряд поверхности микробных клеток наряду с набором специфических структур их оболочек и высоким трансмембранным электрическим потенциалом их цитоплазматических мембран обеспечивает избирательность антимикробного действия антибиотических пептидов. Для рассматриваемого механизма врожденного иммунитета животных характерен относительно низкий по сравнению с молекулярными факторами приобретенного иммунитета уровень распознавания "своего" и "чужого", а также отсутствие памяти. Однако именно антибиотические пептиды клеток и жидких сред в значительной степени обеспечивают выживаемость большинства видов беспозвоночных животных в среде, изобилующей микробами, поскольку у них отсутствуют механизмы приобретенного иммунитета.
Необходимостью обеспечения надежной антимикробной защиты животных объясняется факт удивительного структурного разнообразия молекулярных матриц АП, сосуществующих в клетках и жидких средах одного и того же организма. Так, в нейтрофилах коров представлены Р-дефенсины, бактенецины и индолицидин. В гемолимфе мясных мух встречаются дефенсины, цекропины и другие АП. Один из ведущих идеологов рассматриваемой области исследований - проф. X. Боман из Стокгольмского университета - видит основную причину столь широкого разнообразия молекулярных матриц антимикробных пептидов в особенностях, очень часто ускользающих из поля зрения и анализа ученых, сопряженной эволюции экосистемы животный организм-микробная флора. Другими словами, именно микроорганизмы являются одним из ведущих природных биотических факторов среды, определяющих как поддержание стабильности молекулярно-генетических структур, ответственных за биосинтез антимикробных пептидов, так и направление возможных их изменений в эволюции животного мира. По мнению X. Бомана, именно в реальном многообразии видов микроорганизмов, с которыми в природе взаимодействует конкретный вид животных, кроется одна из основных причин видового спектра антимикробных пептидов, которые, с одной стороны, обеспечивают стерильность внутренней среды макроорганизма, а с другой - контролируют профиль его аутофлоры.
Понимание механизмов антимикробного действия АП является основанием для использования этих соединений в качестве антибиотиков нового поколения в медицине и ветеринарии. Поразительные генетическая изменчивость и скорость размножения микроорганизмов являются факторами, обеспечивающими быстрое формирование у них антибиотикорезистентности. Это обстоятельство создает для человека серьезные проблемы в борьбе с потенциально патогенными возбудителями инфекционных заболеваний. Последний новый класс антибиотиков - налидиксовая кислота - был внедрен в практику более 30 лет тому назад. В связи с этим поиск и создание новых антибиотических веществ пептидной природы являются насущными задачами современной медико-биологической науки.
Традиционно применяемые антибиотики микробного происхождения, решая основную задачу, связанную с инактивацией микроорганизмов, вызывают, однако, у человека и животных ряд нежелательных побочных эффектов: во-первых, многие из них индуцируют в организме состояние иммунодефицита вследствие иммуносупрессивной активности, во-вторых, вызывают в организме эндотоксемию, в-третьих, к ним вырабатывается резистентность со стороны микроорганизмов.
Антимикробные пептиды лишены этих недостатков. Более того, многие катионные АП обладают эндотоксиннейтрализующей и иммунорегулиру-ющей активностью. К ним практически не вырабатывается резистентность со стороны микробов. Будучи факторами проницаемости, катионные АП усиливают действие традиционно используемых антибиотиков, для них пока не описаны анафилактоидоподобные реакции, поскольку они неантигенны. Все это является предпосылкой для создания антимикробных препаратов на основе природных пептидных антибиотиков, совокупность которых представляет собой неотложную систему неспецифической антимикробной защиты, включающуюся в действие без существенно заметного латентного периода, с высокой эффективностью и относительной избирательностью распознавания "свое"-"чужое".
С целью оценки терапевтических свойств дефенсинов в условиях организма нами впервые было изучено влияние суммарных дефенсинов кролика и человека на течение экспериментальной герпетической инфекции на мышах линии СВА. С этой целью мышей заражали интрацеребрально вирусом герпеса в дозе 2 ЛД50/0.02 мл. Дефенсины испытывали по лечебно-профилактической схеме с одноразовой концентрацией белка, равной 100 мкг на животное, что оказалось оптимальным в результате предварительно проведенных экспериментов. У мышей вследствие инфицирования в мозг развивается энцефаломиелит разной степени тяжести. Причем по данным, полученным в нашей лаборатории, максимальная продукция вируса в клетках организма-хозяина имеет место на 3-4-е сут инфекционного процесса. Поэтому именно в эти сроки нами внутрибрюшинно вводились дефенсины инфицированным животным. В результате проведенных опытов получены доказательства защитного действия дефенсинов при экспериментальной герпетической инфекции у мышей, количественно более выраженного в случае использования препаратов кролика по сравнению с пептидами нейтрофилов человека. Испытание активности дефенсинов в этих условиях выявило в принципе те же тенденции, которые были установлены при оценке их прямого антивирусного действия. Эффективнее в обоих случаях были дефенсины кролика, нежели человека, что в некоторой степени можно связать с более основным характером молекул первых, определяющим в значительной степени интенсивность вирусоцидной активности препаратов. Необходимо подчеркнуть, что суммарная лечебная доза препаратов составляла в наших экспериментах 10 мг полипептидов на 1 кг веса животного, в то время как ЛД50 дефенсинов кролика, но нашим данным, находится в пределах 500-700 мг/кг, а дефенсинов человека - 700-800 мг/кг. Соотношение токсической и терапевтической доз дефенсинов позволяет считать нетоксичными эти препараты в используемых концентрациях. Что касается механизмов защитного действия этих полипептидов в условиях организма, то наряду с их прямым антивирусным действием наиболее вероятно их стимулирующее воздействие на клетки мононуклеарной фагоцитирующей системы, которые являются одним из определяющих факторов резистентности к герпетической инфекции. Проверка этих предположений является предметом наших продолжающихся работ.
Результаты данных экспериментов позволяют рассматривать дефенсины в качестве эффективных препаратов, повышающих резистентность организма к вирусной инфекции. Принципиальная возможность их использования в качестве профилактическо-терапевтических средств при инфекционной патологии с неизбежностью ставит вопрос о необходимости изучения влияния дефенсинов на гомеостатические механизмы организма.
Подобные документы
Опиоидные пептиды и физиолого-биохимические аспекты их действия. Обмен регуляторных пептидов. Ферменты обмена нейропептидов при стрессе. Схема введения предшественника лей-энкефалина. Тканевое распределение КПН, ФМСФ-ингибируемой КП и АПФ у самцов крыс.
Пептиды, осуществляющие защиту животных от инфекции, широко представлены в природе от простейших до человека. Наиболее распространенным в эволюции типом антимикробных молекул являются катионные цистинсодержащие полипептиды. Наименьшим по размерам молекулы представителем этой группы антибиотических соединений является додекапептид с 1 дисульфидной связью, выделенный из нейтрофилов коров. Открытые нами протегрины содержат уже 2 дисульфидных мостика, так же как и подобные им соединения из гемоцитов подковообразного краба - тахиплезины и гемолимфы скорпиона - андроктонин. Дефенсины человека, кролика, крысы, мыши и морской свинки, как и р-дефенсины коров и галлинацины кур, содержат в своем составе 6 цистеиновых остатков, образующих 3 внутримолекулярные S-S-связи. Замыкают рассматриваемый ряд антимикробные полипептиды eNAP-1 и eNAP-2 из нейтрофилов лошади, для структуры которых характерно наличие уже 4 дисульфидных групп. Есть основания предполагать, что дисульфидные связи придают молекулам полипептидов повышенную устойчивость к переваривающему действию многочисленных протеиназ лейкоцитарного и микробного происхождения, обеспечивая их пролонгированное функционирование в качестве антибиотических агентов при фагоцитозе и воспалении.
Следует подчеркнуть, что фагоциты и гемоциты являются не единственными клетками организма, содержащими дефенсины и структурно-родственные им полипептиды. Выявлены дефенсины в клетках эпителия тонкой кишки мышей, кролика и человека, в эпителии трахеи и языка коров, в структурах мочеполового и репродуктивного трактов человека.
Присутствие дефенсинов в клетках слизистой тонкого кишечника интересно в связи с представлениями И.И. Мечникова о морфофизиологическкх предпосылках формирования фагоцитарной функции клеток в эволюции. Он рассматривал фагоцитоз как антимикробную функцию специализированных клеток организма, возникшую и развившуюся на основе пищеварительной активности цитоплазмы. Антимикробная функция фагоцитов эволюционно возникла на основе способности простейших поглощать и инактивировать бактерии, которые являются основным объектом их питания. Поэтому уже на заре развития животных перед ними стояла задача инактивации потенциально патогенных микроорганизмов, являющихся для них источником пластических веществ и энергии. Есть основания допускать, что продукция простейшим Entamoebahistolytica пептидов, структурно гомологичных NK-лизинам лимфоцитов, отражает генеральную линию развития механизмов противоинфекционной резистентности у животных. В последние годы дефенсины описаны и у высших растений, причем, несмотря на наличие у них 4 дисульфидных связей, они имеют вторично-третичную структуру, сходную с таковой дефенсинов насекомых. Данные о единстве ряда молекулярных механизмов защиты от инфекции у животных и растений могут служить свидетельством общности их происхождения в эволюции.
Локализация дефенсинов в фагоцитах и клеточно-тканевых структурах организма, "пограничных к инфекции", свидетельствует в пользу их участия в формировании неспецифической антимикробной резистентности в качестве универсальных антибиотиков эндогенного происхождения. Более того, рассматриваемый молекулярный механизм защиты от микробов является, по-видимому, одним из древнейших в эволюции животных, поскольку полипептиды дефенсиновой природы встречаются уже в гемолимфе имаго медоносной пчелы, личинок мясной мухи, жука Zophobasatratus и стрекозы.
Другой распространенной в эволюции матрицей антибиотических пептидов являются цекропины, выявленные первоначально в гемолимфе ряда насекомых, а позже в слизистой тонкого кишечника свиньи. Гены структурно-родственных им пептидов были клонированы и секвеированы из асцидии Styelaclava в лаборатории проф. Лерера. Эта гомологическая группа пептидов представляет собой линейные, не содержащие цистеина молекулы оснбвной природы.
При всем структурном разнообразии большинства описанных в настоящее время антибиотических пептидов все они являются, как правило, одновременно основными и амфипатическими молекулами, проявляющими сродство как к липофобным, так и липофильным средам и соединениям. Положительный заряд и амфифильность молекул антибиотических пептидов лежат в основе их функциональных проявлений,' в частности антимикробного действия. Благодаря им антибиотические пептиды вступают в электростатическое и гидрофобное взаимодействия с анионными фосфолипидами и липополисахаридами мембран микробных клеток-мишеней, которые приводят сначала к их адсорбции на поверхности мембран, а потом к внедрению в двойной липидный слой, что нарушает организацию и целостность оболо-чечных структур микроорганизмов. Подобное воздействие АП на ци-топлазматическую мембрану имеет следствием необратимые повреждения ее структуры и нарушения ее разнообразных функций, результирующим эффектом которых является гибель клеток-мишеней. Частным случаем цитотоксичности АП является их действие на микроорганизмы. В силу мембранотропного механизма антибиотического действия этих пептидов они способны в определенных условиях проявлять повреждающие эффекты в отношении клеток животного организма, который их продуцирует. В связи с этим встает вопрос о том, какие клеточно-молекулярные механизмы обеспечивают прицельность именно антимикробного действия АП, сводя к минимуму их возможные аутоповреждающие эффекты в процессах фагоцитоза и воспаления, а также на поверхности барьерных эпителиев макроорганизма. Как теперь установлено, подобная избирательность действия АП определяется рядом структурных и морфологических факторов. К последним относится, в частности, компартментализация дефенсинов в лизосомоподобных гранулах лейкоцитов и клеток Панета, где они связаны с кислыми мукополисахаридами, которые обеспечивают нейтрализацию цитотоксических молекул. Другие пептиды упакованы в гранулах в форме функционально неактивных предшественников, которые подвергаются ограниченному протеолизу с освобождением антимикробного пептида только в случае активации нейтрофильных гранулоцитов в процессе фагоцитоза. В случае освобождения содержимого лизосомоподобных гранул нейтрофильных гранулоцитов во внеклеточное пространство цитотоксичность дефенсинов нейтрализуется вследствие их взаимодействия с плазменными белками, являющимися в своей основной массе анионными соединениями. В настоящее время установлено, что хорошо известные ингибиторы сериновых протеиназ - а2-макроглобулин, а 1-ингибитор протеиназ, а 1 - антихимотрипсин, антитромбин Ш - обладают повышенным сродством к дефенси-нам, благодаря чему образуются нецитотоксичные комплексы серпины-дефенсины.
Существенно важным фактором, определяющим избирательность действия АП на микроорганизмы, являются особенности поверхностных структур бактерий, низших грибов, оболочечных вирусов и, в меньшей степени, простейших. В клеточной стенке и цитоплазматической мембране бактерий и грибов локализованы молекулы, к которым АП проявляют повышенное сродство в ходе межмолекулярных электростатических и гидрофобных взаимодействий. Так, в частности, в состав липидов цитоплазматической мембраны большинства бактерий входят кислые фосфолипиды в существенно более высокой концентрации, чем в плазмалемму эукариотических клеток. Вследствие повышенного электростатического взаимодействия АП с мембранными структурами, обогащенными кислыми фосфолипидами, наблюдается их преимущественная сорбция на бактериальных клетках как в фаголизосомах фагоцитов, так и во внеклеточной среде в очагах воспаления и на поверхности слизистых и кожных покровов. Благодаря ионному взаимодействию АП концентрируются в областях цитоплазматических мембран бактерий, обогащенных кислыми фосфолипидами. Подобное свойство АП было неоднократно продемонстрировано в модельных условиях с использованием искусственных плоских мембран и липосом. Повреждающее действие АП на мембраны-мишени также зависит и от интенсивности гидрофобных взаимодействий пептида с углеводородными хвостами жирных кислот липидов. Амфипатический характер строения молекул большинства АП обеспечивает их внедрение в двойной липидный слой мембран и нарушение структурной целостности последних. Известно также, что присутствие холестерина в составе липидов мембран эукариотических клеток повышает резистентность последних к повреждающему действию таких пептидов, как магейнин и цекропин.
При анализе рассматриваемых взаимодействий важно также учитывать то обстоятельство, что цитоплазматическая мембрана бактерий непосредственно недоступна для контакта с АП, поскольку покрыта клеточной стенкой, в состав которой у грамположительных бактерий входит пептидогликан, а грамотрицательных - пептидогликан и наружная мембрана, являющаяся дополнительным защитным барьером микробной клетки. Поэтому дефенсины, за исключением некоторых криптдинов, и мягейнины более эффективно поражают invitro грамположительную микрофлору, нежели грамотрицательную. Однако это правило распространяется не на все группы антибиотических пептидов. Например, цекропины и некоторые бактенецины более активны как цитотоксические молекулы в отношении грамотрицательных бактерий. Это свойство указанных соединений обусловлено их повышенной способностью к взаимодействию с одним из типичных представителей структуры наружной мембраны грамотрицательных бактерий, каковым является липополисахарид. Наличие липополисахарида во внешнем слое наружной мембраны грамотрицательных бактерий является одним из условий их опознания антибиотическими пептидами и белками.
Пенетрирующая липофильный бислой активность молекул антибиотических пептидов в ряде случаев заметно зависит от трансмембранного электрического потенциала клеток-мишеней, который, как известно, у плазмалеммы бактерий обычно в 1.5-2 раза более высокий, чем у мембран эукариотических клеток. Это свойство мембран является одним из условий, облегчающих проникновение АП через липидный бислой, особенно в тех случаях, когда эффекторные молекулы умеренно - или слабоосновные. Проникающая способность высокоосновных пептидов, таких как дефенсины кролика NP-1 и NP-2, практически не зависит от мембранного потенциала клеток-мишеней. Направление электрического поля поперек мембран от плюса на внешней поверхности мембраны к минусу - на внутренней обеспечивает электрофорез положительно заряженных молекул антибиотических пептидов через мембрану внутрь клетки. При этом часть молекул самостоятельно или в ассоциации друг с другом внедряется в мембраны, образуя в них пороподобные отверстия различной молекулярной организации. Перфорация мембран, которая в ряде случаев носит транзиторный характер, приводит к утечке из клеток ионов и нарушению ионной асимметрии между средой и клетками. Следствием этого является диссипация мембранного потенциала клеток-мишеней и стремление молекул воды войти в клетки. Результатами подобного движения воды могут быть разбухание клеток-мишеней и их осмотический лизис. Рассеивание мембранного потенциала лишает клетки возможности осуществлять активный транспорт ионов и веществ против градиента концентраций, что в итоге резко снижает их жизнеспособность.
При типичной ориентации электрического поля мембран катионные АП могут переноситься через этот барьер путем электрофореза, подобно тому как это описано для микробных пептидных антибиотиков полимиксина В и грамицидинов., что дополняет картину деструктурирующих воздействий наv клетки-мишени, результатом которых является гибель последних. Обобщенная схема предполагаемого механизма антимикробного действия одного из антибиотических пептидов - цекропина - представлена на рис. 20. Первая фаза действия пептида связана с адсорбцией димера цекропина на поверхности мембраны за счет электростатических взаимодействий с ее отрицательно заряженными соединениями. На следующей стадии происходит диссоциация димера и внедрение в липофильную фазу липидного бислоя отдельных молекул цекропина за счет гидрофобных взаимодействий. На заключительной стадии внедрившиеся и ориентированные поперек мембран молекулы цекропинов формируют каналоподобные поры с умеренной селективной проницаемостью для анионов.
Схема, отражающая основные этапы взаимодействия цекропинов с липидной мембраной.

Таким образом, отрицательный заряд поверхности микробных клеток наряду с набором специфических структур их оболочек и высоким трансмембранным электрическим потенциалом их цитоплазматических мембран обеспечивает избирательность антимикробного действия антибиотических пептидов. Для рассматриваемого механизма врожденного иммунитета животных характерен относительно низкий по сравнению с молекулярными факторами приобретенного иммунитета уровень распознавания "своего" и "чужого", а также отсутствие памяти. Однако именно антибиотические пептиды клеток и жидких сред в значительной степени обеспечивают выживаемость большинства видов беспозвоночных животных в среде, изобилующей микробами, поскольку у них отсутствуют механизмы приобретенного иммунитета.
Необходимостью обеспечения надежной антимикробной защиты животных объясняется факт удивительного структурного разнообразия молекулярных матриц АП, сосуществующих в клетках и жидких средах одного и того же организма. Так, в нейтрофилах коров представлены Р-дефенсины, бактенецины и индолицидин. В гемолимфе мясных мух встречаются дефенсины, цекропины и другие АП. Один из ведущих идеологов рассматриваемой области исследований - проф. X. Боман из Стокгольмского университета - видит основную причину столь широкого разнообразия молекулярных матриц антимикробных пептидов в особенностях, очень часто ускользающих из поля зрения и анализа ученых, сопряженной эволюции экосистемы животный организм-микробная флора. Другими словами, именно микроорганизмы являются одним из ведущих природных биотических факторов среды, определяющих как поддержание стабильности молекулярно-генетических структур, ответственных за биосинтез антимикробных пептидов, так и направление возможных их изменений в эволюции животного мира. По мнению X. Бомана, именно в реальном многообразии видов микроорганизмов, с которыми в природе взаимодействует конкретный вид животных, кроется одна из основных причин видового спектра антимикробных пептидов, которые, с одной стороны, обеспечивают стерильность внутренней среды макроорганизма, а с другой - контролируют профиль его аутофлоры.
Понимание механизмов антимикробного действия АП является основанием для использования этих соединений в качестве антибиотиков нового поколения в медицине и ветеринарии. Поразительные генетическая изменчивость и скорость размножения микроорганизмов являются факторами, обеспечивающими быстрое формирование у них антибиотикорезистентности. Это обстоятельство создает для человека серьезные проблемы в борьбе с потенциально патогенными возбудителями инфекционных заболеваний. Последний новый класс антибиотиков - налидиксовая кислота - был внедрен в практику более 30 лет тому назад. В связи с этим поиск и создание новых антибиотических веществ пептидной природы являются насущными задачами современной медико-биологической науки.
Традиционно применяемые антибиотики микробного происхождения, решая основную задачу, связанную с инактивацией микроорганизмов, вызывают, однако, у человека и животных ряд нежелательных побочных эффектов: во-первых, многие из них индуцируют в организме состояние иммунодефицита вследствие иммуносупрессивной активности, во-вторых, вызывают в организме эндотоксемию, в-третьих, к ним вырабатывается резистентность со стороны микроорганизмов.
Антимикробные пептиды лишены этих недостатков. Более того, многие катионные АП обладают эндотоксиннейтрализующей и иммунорегулиру-ющей активностью. К ним практически не вырабатывается резистентность со стороны микробов. Будучи факторами проницаемости, катионные АП усиливают действие традиционно используемых антибиотиков, для них пока не описаны анафилактоидоподобные реакции, поскольку они неантигенны. Все это является предпосылкой для создания антимикробных препаратов на основе природных пептидных антибиотиков, совокупность которых представляет собой неотложную систему неспецифической антимикробной защиты, включающуюся в действие без существенно заметного латентного периода, с высокой эффективностью и относительной избирательностью распознавания "свое"-"чужое".
С целью оценки терапевтических свойств дефенсинов в условиях организма нами впервые было изучено влияние суммарных дефенсинов кролика и человека на течение экспериментальной герпетической инфекции на мышах линии СВА. С этой целью мышей заражали интрацеребрально вирусом герпеса в дозе 2 ЛД50 /0.02 мл. Дефенсины испытывали по лечебно-профилактической схеме с одноразовой концентрацией белка, равной 100 мкг на животное, что оказалось оптимальным в результате предварительно проведенных экспериментов. У мышей вследствие инфицирования в мозг развивается энцефаломиелит разной степени тяжести. Причем по данным, полученным в нашей лаборатории, максимальная продукция вируса в клетках организма-хозяина имеет место на 3-4-е сут инфекционного процесса. Поэтому именно в эти сроки нами внутрибрюшинно вводились дефенсины инфицированным животным. В результате проведенных опытов получены доказательства защитного действия дефенсинов при экспериментальной герпетической инфекции у мышей, количественно более выраженного в случае использования препаратов кролика по сравнению с пептидами нейтрофилов человека. Испытание активности дефенсинов в этих условиях выявило в принципе те же тенденции, которые были установлены при оценке их прямого антивирусного действия. Эффективнее в обоих случаях были дефенсины кролика, нежели человека, что в некоторой степени можно связать с более основным характером молекул первых, определяющим в значительной степени интенсивность вирусоцидной активности препаратов. Необходимо подчеркнуть, что суммарная лечебная доза препаратов составляла в наших экспериментах 10 мг полипептидов на 1 кг веса животного, в то время как ЛД50 дефенсинов кролика, но нашим данным, находится в пределах 500-700 мг/кг, а дефенсинов человека - 700-800 мг/кг. Соотношение токсической и терапевтической доз дефенсинов позволяет считать нетоксичными эти препараты в используемых концентрациях. Что касается механизмов защитного действия этих полипептидов в условиях организма, то наряду с их прямым антивирусным действием наиболее вероятно их стимулирующее воздействие на клетки мононуклеарной фагоцитирующей системы, которые являются одним из определяющих факторов резистентности к герпетической инфекции. Проверка этих предположений является предметом наших продолжающихся работ.
Результаты данных экспериментов позволяют рассматривать дефенсины в качестве эффективных препаратов, повышающих резистентность организма к вирусной инфекции. Принципиальная возможность их использования в качестве профилактическо-терапевтических средств при инфекционной патологии с неизбежностью ставит вопрос о необходимости изучения влияния дефенсинов на гомеостатические механизмы организма.
Антибиотики этой группы широко распространены в природе, они продуцируются различными видами актиномицетов, бактерий, грибов и весьма разнообразны по своему химическому строению — столь разнообразны, что их химическая классификация становится размытой, т.е. можно выделить слишком много различных подгрупп. Поэтому мы ограничимся лишь анализом наиболее типичных структур, ярких примеров и тех структурных особенностей, которые являются достаточно общими.
В первую очередь, мы должны, конечно же, отметить, что антибиотики этого типа обязательно содержат аминокислоты: количество аминокислотных остатков может варьироваться от нескольких единиц (антибиотик цикло-серин образован только одной аминокислотой) до нескольких сотен (неокар-циностатин содержит 109 аминокислот, аспарагиназы и того более: их молекулярная масса колеблется от 30 до 130 тысяч Дальтон).
Во вторую очередь следует обратить внимание на “качество” аминокислот, участвующих в образовании пептидных антибиотиков: это могут быть как обычные протеиногенные a-L-аминокислоты, так и непротеиногенные аминокислоты, кроме того — здесь сплошь и рядом участвуют а-аминокислоты D-конфигурации. А если учесть различные оксикислоты, вклинивающиеся в полипептидные цепочки, то можно увидеть, насколько разнообразно “качество" субъединиц, участвующих в построении молекулярных структур пептидных антибиотиков.
Третий, достаточно общий структурный фактор пептидных антибиотиков — это вид полипептидной (или депсипеп-тидной) цепочки: либо это открытая классическая цепь, либо циклополипептидная структура, либо комбинированная система из цикла и открытой цепочки, либо система нескольких небольших циклов.
Пептидным антибиотиком минимальной структуры можно считать циклосерин, который и назвать то пептидным мы можем условно (поскольку в нем нет классических пептидных связей), на том лишь основании, что он образован из серина D-конфигурации с фрагментом -CO-NH-. По химической структуре он относится к гетероциклам оксазолам, проявляет противотуберкулезную активность, продуцируется Streptomyces gariphcilus, Str. Or-chidciceits и др., но в настоящее время его получают синтетическим путем.
Представителями достаточно простых полипептидных антибиотиков являются грамицидины А, В, С и D (линейные) и грамицидин S (циклический), продуцируемые Bacillus brevis (схема 11.4.1). Все они образованы протеиногенными аминокислотами, с той лишь особенностью, что некоторые из них имеют D-конфигурацию — D-Leu, D-Val, D-Phe, а в общем балансе их преобладают гидрофобные аминокислоты. Их биологический эффект связан с нарушением ионного транспорта через мембраны бактерий: линейные грамицидины (А) встраиваются в мембрану в виде димерной спирали с осевой полостью, которая выполняет роль ионного канала; циклически грамицидины (S) выполняют ту же роль, но только по другому механизму — по механизму “гость-хозяин”, когда ион включается внутрь макроцикла и в таком виде транспортируется через мембрану клетки. Этими же микробами продуцируются весьма
близкие по структуре циклополипептидные антибиотики туроцидины.
Очень разнообразны пептидные антибиотики, образованные непротеиногенными аминокислотами: как правило, это циклополипептиды (схема 11.4.2).
Они очень разнообразны и по биологической активности — от росторегулирующей (бацитрацин А), что используется в ветеринарии, до туберкулостатической (туберактиномицин, капреомицин).
К этой же подгруппе полипептидных антибиотиков относятся полимиксины, продуцируемые различными штаммами Bacillus polymyxa и В. Circulaus (схема 11.4.3), важной биологической особенностью которых является высокая активность по отношению к грам-отрицательным бактериям, особенно к Pseudomonas aeruginosa, что и используется в медицинской практике.
Из культуры молочнокислого стрептококка Streptococcus lactis выделена группа полипептидных антибиотиков низинов, представленных полициклической полипептидной структурой, все циклы которой сформированы специфическими сульфидными мостиками на базе непротеиногенной аминокислоты лантионина и ее 3-метилзамещенного производного. В построении этого антибиотика принимают участие также ненасыщенные аминокислоты (схема 11.4.4).
Характерными структурными элементами этого антибиотика являются непредельные аминокислоты дегидроаланин и дегидроаминомасляная кислота, которые способны взаимодействовать с сульфгидрильными (-SH) группами некоторых ферментов, на чем и основано (как предполагают) их биологическое действие. Он активен против
многих грам-положительных бактерий, некоторых кислотоустойчивых бактерий, в том числе, спорообразующих бактерий и используется как пищевой консервант сыра, овощей, фруктов.
К пептидным антибиотикам можно отнести и биологически активные депсипептиды валиномицин (Streptomices fulvissimus) и энниатины (грибы рода Fusarium), которые являются природными ионофорами (схема 11.4.5). Они весьма близки по химической структуре, но валиномицин более специфический агент по отношению к ионам на чем и основана его активность. В его присутствии проницаемость мембран чувствительных бактериальных клеток увеличивается в раз, а это неминуемо приводит к их гибели.

Обзор
Антимикробный пептид на поверхности модели бактериальной мембраны. Связывание пептида приводит к дестабилизации мембраны и проникновению воды внутрь бислоя.
Автор
Редакторы
После появления пенициллина более чем 80 лет человечество ведет ожесточенную борьбу с патогенными микроорганизмами. Широкое использование антибиотиков в терапии инфекционных заболеваний породило новые устойчивые к ним формы бактерий. На пороге XXI века медики и фармацевты столкнулись с проблемой поиска альтернативы традиционным антибиотикам. Для этой цели использование антимикробных пептидов может оказаться весьма эффективным.

Рисунок 1. Представители различных групп антимикробных пептидов.
1 — Линейные α-спиральные; 2 — не имеющие характерной структуры, напр. богатые определенной аминокислотой; 3 — дефензины; 4 — лантибиотики (низин).
Основная цепь пептидов изображена в виде ленты (1, 2). Фрагменты β-слоя представлены стрелками (3). Для низина (4) показаны также боковые цепи остатков.
Антимикробные пептиды
Несмотря на огромное разнообразие, на основании структурной организации антимикробные пептиды можно разделить на несколько групп:
- линейные α-спиральные пептиды (напр., магаинин, меллитин, цекропин);
- пептиды, богатые определенной аминокислотой, напр., гистидином или пролином;
- пептиды, имеющие сложную пространственную организацию, содержащие дисульфидные мостики, тиоэфирные циклы и др. К ним относятся дефензины, протегрины и лантибиотики — высокоактивные бактериальные пептиды (напр., низин — см. выше).
Тем не менее, для большинства таких пептидов характерно несколько общих черт: высокий положительный заряд (+2 — +9), пространственное разделение гидрофобных и гидрофильных участков молекулы (амфифильность). Амфифильность является важной особенностью антимикробных пептидов, которая обеспечивает возможность одновременно выгодно взаимодействовать с гидрофобным ядром липидной мембраны и полярным окружением (например, с водой).
Механизмы действия
Выделяют два основных типа воздействия антимикробных пептидов на клетки: ингибирование метаболитических процессов или нарушение целостности клеточной мембраны [5]. Большинство антимикробных пептидов вызывают гибель клетки по второму механизму. Лишь для некоторых пептидов показано действие по первому типу, среди них лантибиотики, которые селективно связываются с предшественником бактериальной стенки [6]. Для того, чтобы достигнуть плазматической бактериальной мембраны, пептиды предварительно должны преодолеть несколько слоев клеточной стенки. В случае грам-отрицательных бактерии она состоит из внешней липополисахаридной мембраны и пептидогликанового слоя. У грам-положительных бактерий внешняя мембрана отсутствует, однако пептидогликановый слой развит гораздо сильнее. Пептиды обычно беспрепятственно достигают плазматической мембраны, и именно ее разрушение приводит гибели клетки.
Перспективы использования антимикробных пептидов в медицине

Рисунок 2. Антимикробный пептид рамопланин (Ramoplanin).
Продукт нерибосомального синтеза в микроорганизмах рода Actinomycetes spp. Механизм действия основан на связывании с предшественником бактериальной стенки липидом II. На данный момент находится на клинических испытаниях фазы III в качестве антибиотика против инфекций дыхательных путей (в первую очередь, стафилококков).
Несмотря на то, что для антимикробных пептидов характерно довольно высокие действующие концентрации (~10 −7 —10 −6 моль, 10 −9 моль у лантибиотиков) и низкая селективность, они обладают некоторыми преимуществами: способность быстро убивать клетки-мишени, широкий спектр действия, активность в отношении штаммов, резистентных к другим антибиотикам, а также относительная трудность в развитии устойчивости. Поскольку некоторые антимикробные пептиды обладают цитотоксическим эффектом (действуют на эукариотические клетки), наиболее эффективно они могут быть использованы при лечении заболеваний наружных покровов, слизистых — без введения в кровь пациента. На сегодняшний день такие пептиды активно используются для создания новых лекарственных препаратов. Наиболее успешным примером, является циклический пептидный антибиотик рамопланин, который уже находится на стадии клинических испытаний для лечения инфекционных заболеваний дыхательных путей. Также показано, что ингаляции смеси, содержащей антимикробные пептиды, являются эффективными в терапии туберкулеза.
Все это позволяет рассматривать описанные выше молекулы в качестве основы для создания эффективных лекарств, особенно на фоне снижения потенциала обычных антибиотиков.
Читайте также: