
Ядра спинного мозга гистология кратко
Обновлено: 03.07.2024
Строение спинного мозга. Структура спинного мозга.
В спинном мозге различают серое и белое вещество. На поперечном срезе спинного мозга серое вещество имеет вид буквы Н. Выделяют передние (вентральные), боковые, или латеральные (нижние шейные, грудные, два поясничных), и задние (дорсальные) рога серого вещества спинного мозга.
Серое вещество представлено телами нейронов и их отростками, нервными окончаниями с синаптическим аппаратом, макро- и микроглией и сосудами.
Белое вещество окружает снаружи серое вещество и образовано пучками мякотных нервных волокон, которые формируют проводящие пути на протяжении всего спинного мозга. Эти пути направляются в сторону головного мозга или нисходят из него. Сюда же относятся волокна, направляющиеся в выше- или нижележащие сегменты спинного мозга. Кроме того, в белом веществе находятся астроциты, отдельные нейроны, гемокапилляры.
В белом веществе каждой половины спинного мозга (на поперечном срезе) различают три пары столбов (канатиков): задний (между задней срединной перегородкой и медиальной поверхностью заднего рога), боковой (между передним и задним рогами) и передний (между медиальной поверхностью переднего рога и передней срединной щелью).
В центре спинного мозга проходит канал, выстланный эпендимоцитами, среди которых различают малодифференцированные формы, способные, по данным некоторых авторов, к миграции и дифференцировке в нейроны. В нижних сегментах спинного мозга (поясничном и сакральном) после полового созревания происходит пролиферация глиоцитов и зарастание канала, образование интраспиналъного органа. В составе последнего находятся глиоциты и секреторные клетки, вырабатывающие вазоактивный нейропептид. Орган подвергается инволюции после 36 лет.
Нейроны серого вещества спинного мозга являются мультиполярными. Среди них различают нейроны с немногочисленными слабоветвящимися дендритами, нейроны с ветвящимися дендритами, а также переходные формы.
В зависимости от того, куда идут отростки нейронов, выделяют: внутренние нейроны, отростки которых заканчиваются синапсами в пределах спинного мозга; пучковые нейроны, нейрит которых идет в составе пучков (проводящих путей) в другие отделы спинного мозга или в головной мозг; корешковые нейроны, аксоны которых покидают спинной мозг в составе передних корешков.
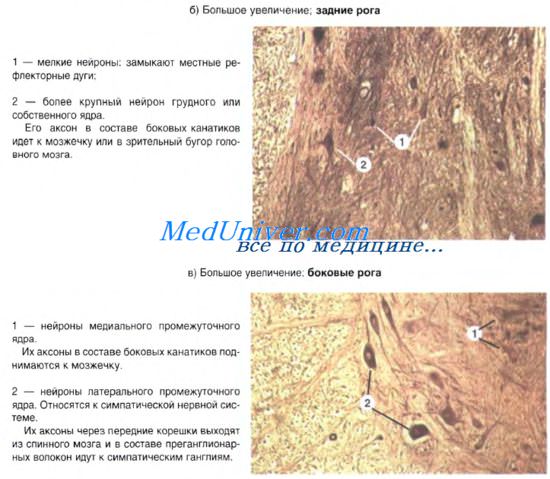
На поперечном срезе нейроны группируются в ядра, в составе которых находятся сходные по строению и функции нейроны. На продольном срезе эти нейроны располагаются послойно в виде колонки, что отчетливо видно в области заднего рога. Нейроны каждой колонки иннервируют строго определенные области тела. О закономерностях группировки нейронов и их функциях можно судить по пластинам Рекседа (1-Х). В центре заднего рога располагается собственное ядро заднего рога, у основания заднего рога — грудное ядро (Кларка), латеральнее и несколько глубже располагаются базилярные ядра, в промежуточной зоне — медиальное промежуточное ядро. В дорсальной части заднего рога из глубины кнаружи последовательно располагаются мелкие нейроны студневидного вещества (роландова), далее — мелкие нейроны губчатой зоны и, наконец, — пограничная зона, содержащая мелкие нейроны.
Аксоны чувствительных нейронов из спинальных ганглиев входят в спинной мозг через задние корешки и далее в краевую зону, где делятся на две ветви: короткую нисходящую и длинную восходящую. По боковым ответвлениям от этих ветвей аксона импульсы передаются на ассоциативные нейроны серого вещества. Болевая, температурная и тактильная чувствительность проецируется на нейроны студневидного вещества и собственного ядра заднего рога. В составе студневидного вещества находятся интернейроны, продуцирующие опиоидные пептиды, которые влияют на болевые ощущения (так называемые "ворота боли"). Импульсы от внутренних органов передаются на нейроны ядер промежуточной зоны. Сигналы от мышц, сухожилий, суставных капсул и др. (проприорецепция) направляются на ядро Кларка и другие ядра. Аксоны нейронов этих ядер формируют восходящие проводящие пути.
В задних рогах спинного мозга много диффузно расположенных нейронов, аксоны которых заканчиваются в пределах спинного мозга той же или противоположной стороны серого вещества. Аксоны этих нейронов выходят в белое вещество и тут же делятся на нисходящую и восходящую ветви. Распространяясь на уровне 4-5 спинномозговых сегментов, эти ветви образуют в совокупности собственные пучки белого вещества, непосредственно прилежащие к серому веществу. При этом различают задний, боковой и передний собственные пучки. Все эти пучки белого вещества относятся к собственному аппарату спинного мозга. От аксонов, входящих в состав собственных пучков, отходят коллатерали, заканчивающиеся синапсами на двигательных нейронах. Благодаря этому создаются условия для лавинообразного нарастания числа нейронов, передающих импульсы по рефлекторным дугам собственного аппарата спинного мозга.



Нейробласты — клетки с большим округлым ядром, плотным ядрышком и бледной цитоплазмой — дают начало всем нейронам ЦНС. Нейроны — классический пример клеток, относящихся к статической популяции. Ни при каких условиях они in vivo не способны к пролиферации и обновлению. Обонятельные нейроны (происходят из обонятельных плакод) эпителиальной выстилки носовых ходов — единственное известное исключение.
Глиобласты — предшественники макроглии [астроциты и олигодендро(глио)циты]. Все типы макроглии способны к пролиферации.
Генез клеток микроглии спорен. Согласно наиболее распространённой точке зрения, клетки микроглии относят к системе мононуклеарных фагоцитов. Достаточно вероятным представляется их нейроэктодермальный генез. В этом случае подразумевается гетерогенность популяции клеток микроглии.
Раннее развитие мозга



Морфогенетические процессы — индукция, адресная миграция клеток, направленный рост аксонов, гибель клеток, так называемые нейротрофические взаимодействия — хорошо выражены в развивающейся нервной системе. Жёсткость организации мозга определяют два момента: адресная миграция клеток и направленный рост их отростков.
Адресная миграция клеток


Направленный рост аксонов
Направленный рост аксонов, как и адресная миграция клеток, осуществляется в рамках концепции “сигнал–ответ”. Эта концепция объясняет, как нейрон узнаёт свою область иннервации и находит своего клеточного партнёра, и как в развивающемся мозге многочисленные переплетающиеся отростки нейронов устанавливают связи с высокой точностью. Отросток нейрона — аксон — сразу и без ошибок находит свои мишени. Направленный рост аксонов осуществляет конус роста.
Конус роста — мобильная терминаль отростка нейрона, специализированная на удлинение и навигацию этого отростка, впервые детально описан Сантъяго Рамон-и-Кахалем. Конус имеет на конце булавовидное утолщение (ламеллоподия), от которого отходят тонкие пальцевидные отростки — филоподии. Они растут в различных направлениях и исследуют потенциальное пространство роста аксона. Согласно представлению С. Рамон-и-Кахаля о хемотропизме, рост аксонов происходит по градиенту концентрации специфических химических факторов, вырабатываемых в мишенях. Действительно, in vitro градиенты фактора роста нервов (NGF) и других веществ (например, ацетилхолина) влияют на направление роста аксонов. Наиболее разработано представление о меченых путях, которые образованы молекулярными метками (своего рода знаками навигационной обстановки), закономерно распределёнными в потенциальном пространстве роста аксонов. По мере роста пионерский аксон последовательно считывает одну за другой метки, расположенные в межклеточном пространстве или на поверхности клеток, и растёт в нужном направлении. Вслед за ним мигрируют отростки других аксонов, совокупность которых формирует тракты в ЦНС и нервы на периферии. Примером клеток, направляющих рост аксонов, могут служить временно живущие нейроны Кахаля–Ретциуса. Ключевой момент представления о заранее размеченных путях — узнавание — обеспечивают молекулы адгезии, которые встроены в плазмолемму ламеллоподии и филоподий и взаимодействуют с комплементарными молекулами (ламинин, фибронектин, коллаген, тенасцин и др.) во внеклеточном матриксе. Это обеспечивают фиксацию конуса роста на поверхности мишени в нужном месте и в нужное время.


Характеристика
Молекул внеклеточного матрикса
Ламинин, тенасцины, коллаген, фибронектин, иммуноглобулины, аносмин‑1 ( KAL ), хондроитин/гепарансульфат протеогликаны, тромбоспондин‑1
HomA , протеогликаны, интегрины
Многие из них важны для нейроонтогенеза. Все продолжают экспрессироваться в зрелом мозге
Sema 3A‑F, Sema 4A‑G, Sema 5A и B, Sema 6A‑C, Sema 7A
Плексин A, нейропилин
Хеморепелленты, вызывают коллапс конуса роста сенсорных аксонов in vivo . Некоторые разновидности могут выступать в роли аттрактантов. Могут быть интегральными мембранными белками или секретироваться
N – CAM , PSA - N – CAM , L 1, TAG -1/аксонин‑1, DM - Grasp , кадгерины
Интегральные и мембраносвязанные гликопротеины, участвуют в прикреплении аксонов к субстрату, фасцикуляции, направленном росте, в гомофильных и гетерофильных взаимодействиях
Миелин-ассоциированный гликопротеин ( MAG ), nogo
Интегральные мембранные белки экспрессируются в зрелом миелине, ингибиторы роста аксонов in vitro и in vivo . Могут выступать в роли рецепторов для некоторых молекул внеклеточного матрикса
Рецепторов, связанных с тирозинкиназам
Ephrin A1-A5, Ephrin B1-B3
Лиганды и рецепторы, связаны с мембраной или являются трансмембранными белками, участвуют в процессах миграции клеток и роста аксонов в нейроонтогенезе
DCC, нейрогенин, Unc ‑ 5
Важны для миграции нейробластов и направленного роста аксонов в развивающемся мозге. Связывание с рецептором DCC прослежено как при позитивном, так и негативном регулировании
NGF, NT-3, BDNF, CNTF, GDNF
Trk A,B,C, p75, Lifrb, GFRa1
Нейротропная и нейротрофичекая активность
FGF, IGF, PDGF, VEGFa
FGFr1-4, flk-1, IGFr1, PDGFr, EGFr
Нейротрофическая активность, контроль пролиферации, дифференцировки и гибели клеток
TGF b , LIF, TNF, EGF
TGF b тип I; TGF b , тип II; LIFrb, gp130, TNFr1-2
Нейротрофичекая активность, поддержание дифференцировки нейронов
Возможно, участвуют в синаптогенезе. Изменяют направление роста аксонов чувствительных нейронов
Репелленты для различных аксонов и мигрирующих клеток, но и стимуляторы ветвления и удлинения аксонов
Механизм роста аксона
Перемещение конуса роста осуществляется при помощи молекулярного мотора. В конусе роста различают центральный и периферический домен. Периферический домен объединяет филоподии и часть ламеллоподии. Здесь актин полимеризуется на ведущем конце нити, что поддерживает рост аксона в ортоградном направлении, но тянется в обратном (ретроградном) направлении за счет работы миозинового мотора. Ретроградное перемещение актина сдерживает проникновение микротрубочек в периферический домен. Сигналы взаимодействия поверхностных рецепторов с лигандами угнетают ретроградное перемещение актиновых нитей и равновесие между сборкой и перемещением актиновых нитей в ортоградном направлении, с одной стороны, и ретроградным перемещением актиновых нитей за счет работы мотора, с другой стороны, нарушается в пользу первого и в итоге обеспечивает перемещение (рост) всей структуры конуса роста в ортоградном направлении. Остановка ретроградного перемещения актина позволяет микротрубочкам проникать в периферический домен. Этот рост микротрубочек осуществляется путём полимеризации тубулина. В результате аксон удлиняется.
Реакции актина на ведущем конце ламеллоподии . Сеть цитоскелета на ведущем конце ламеллоподии содержит интенсивно ветвящиеся актиновые нити с (+)–концом, направленным в сторону роста, и (–)–концом, образующим угол ~70º с другими нитями. В состоянии покоя (+)-конец актиновой нити обычно покрыт кэппинг–белком. Под действием стимула происходит полимеризация актина на свободном (+)–конце нити, расположенной в ведущей части конуса роста. Нить удлиняется и возникает усилие в направлении поверхностной мембраны. В диссоциации кэппинг–белка и освобождении (+)–конца актиновой нити имеют значение увеличение уровня инозитолдифосфата, разрезание нити ADF/кофилином и нуклеация актина, зависимая от комплекса Arp2/3. Этот комплекс активируется при связывании с белком N – WASP ( W iskott - A ldrich S yndrome P rotein , тяжелая врожденная нейтропения, тромбоцитопения), одна из форм которого, в свою очередь, активируется при участии Cdc42.
Физиологическая гибель клеток
Запрограммированная массовая смерть нейронов происходит на строго определённых этапах онтогенеза. Естественная гибель нейронов прослежена как в ЦНС, так и в периферической нервной системе. Объём субпопуляции гибнущих нейронов оценивают в широком интервале, от 25 до 75%. Иногда в популяции погибают все нейроны (например, несущие метку для направленного роста аксонов). Выраженная гибель нейронов в сформированной нервной ткани наблюдается при дегенеративных заболеваниях нервной системы, таких как болезнь Альцхаймера, Паркинсона, Хантингтона, Кройтцфельдта–Якоба, боковой амиотрофический склероз и др.
Эта концепция предусматривает информационный обмен между нейронами, отличающийся от химической передачи возбуждения в синапсах. Согласно концепции, такой обмен поддерживает фенотип взаимодействующих клеток на уровне, адекватном выполнению их функции. Нейротрофические взаимодействия реализуются через нейротрофические факторы. Эти химические факторы вырабатываются в одних нейронах или в ненервных клетках и воздействуют на другие нейроны. Нейротрофические факторы поддерживают выживание и дифференцировку нейронов в нейроонтогенезе и при регенерации. Наиболее изученным нейротрофическим фактором является фактор роста нервов ( NGF ).
Нейроны относят к статической клеточной популяции. Это означает, что в постнатальном онтогенезе не происходит образования новых нейронов. Следовательно, погибающие нейроны не восстанавливаются. Но из этого не следует, что в нервной системе отсутствует регенерация. Она осуществляется за счёт восстановления целостности повреждённых нейронов, роста их отростков, размножения глиальных и шванновских клеток. Всё это, а также изменение связей между регенерирующими и интактными клетками при благоприятных условиях, может привести к восстановлению функции в нервной системе.
Нейроны (термин предложил Вильгельм фон Вальдейер) — главные клеточные типы нервной ткани. Эти возбудимые клетки осуществляют передачу электрических сигналов (между собой при помощи нейромедиаторов в синапсах) и обеспечивают способность мозга к переработке информации.



Перикарион содержит ядро, комплекс Гольджи, гранулярную эндоплазматическую сеть, митохондрии, лизосомы, элементы цитоскелета.
Ядро нейрона имеет мелкодисперсный хроматин и ядрышко. В силу относительно большого диаметра ядро в световом микроскопе выглядит (особенно в крупных нейронах) как оптически пустое. Ядрышко крупное и резко базофильное.
Комплекс Гольджи хорошо развит, особенно в крупных нейронах. Его особенность — расположение между ядром и местом отхождения аксона, что отражает мощный транспорт белков, синтезированных в гранулярной эндоплазматической сети перикариона, в аксон.
Аксонный холмик — свободная от гранулярной эндоплазматической сети и рибосом область перикариона, содержащая много микротрубочек и нейрофиламентов, место, где начинается аксон и генерируется потенциал действия.
Гранулярная эндоплазматическая сеть. В перикарионе и дендритах развита гранулярная эндоплазматическая сеть. Она соответствует глыбкам хроматофильного вещества, которые впервые обнаружил Франц Ниссль при окраске метиленовым синим, почему её в нейронах иногда называют веществом Ниссля (тигроид). Тигролиз — распыление хроматофильного (тигроидного) вещества, отражающее глубокие дистрофические изменения при нарушении целостности нейрона ( например , при сдавлении или перерезке аксона).
Митохондрии многочисленны. Значительные энергетические потребности нервных клеток обеспечива ет преимущественно аэробный метаболизм, почему нейроны крайне чувствительны к гипоксии.
Пигменты . В нейронах (особенно с возрастом) накапливается липофусцин. Нейроны некоторых ядер мозга нормально содержат иные пигменты, почему эти образования и получили своё название (substantia nigra, locus coeruleus).
Отростки, отходящие от перикариона, — аксон и дендриты (рис. 8–13). Отростки нейрона участвуют в образовании синапсов.
Аксон (нейрит) — длинный отросток, как правило, не ветвящийся, но образующий концевые разветвления, содержащие синаптические пузырьки; проводит пачки импульсов (спайки) от перикариона. Объём аксона может достигать 99% суммарного объёма нейрона. Длина аксона может быть весьма значительной — десятки сантиметров.
Синтез белка в аксоне . Считается общепринятым, что синтез белка в нейроне происходит только в перикарионе и дендритах. Однако в последнее время появились достаточно убедительные доказательства синтеза белка в аксоне. Многочисленные белки аксоплазмы синтезируются в перикарионе и перемещаются в составе медленного аксонного транспорта. Длина аксона варьирует от нескольких микрон до метра, и транспортировка белковых молекул по аксону может занять дни, недели и даже месяцы. Следовательно, при срочном ответе нейрона на изменившиеся условия функционирования локальный синтез белка экономит время и энергию, необходимые для транспортировки вновь синтезированного белка в нужный компартмент аксона. Учитывая тот факт, что белки имеют определённый период полураспада и во время медленного транспорта по аксону постоянно подвергаются деградации можно предположить, что внутриаксонный синтез белка также необходим для поддержания белкового состава аксоплазмы и массы аксона в целом.
Цитоскелет нейронов состоит из микротрубочек, промежуточных филаментов (нейрофиламенты) и микрофиламентов.

Читайте также: