
Транскрипция у прокариот кратко
Обновлено: 08.07.2024
В Транскрипция ДНК это процесс, с помощью которого информация, содержащаяся в дезоксирибонуклеиновой кислоте, копируется в форме подобной молекулы, РНК, либо в качестве предварительной стадии для синтеза белка, либо для образования молекул РНК, которые участвуют во множественных клеточных процессах большое значение (регуляция экспрессии генов, передача сигналов и т. д.).
Хотя неверно, что все гены организма кодируют белки, верно то, что все белки клетки, будь то эукариотические или прокариотические, кодируются одним или несколькими генами, где каждая аминокислота представлена набор из трех оснований ДНК (кодон).
Синтез полипептидной цепи любого клеточного белка происходит благодаря двум фундаментальным процессам: транскрипции и трансляции; оба сильно регулируются, поскольку они представляют собой два процесса, очень важных для функционирования любого живого организма.
Что такое транскрипция ДНК?
Этот процесс осуществляется ферментом, называемым РНК-полимеразой, который распознает определенные места в последовательности ДНК, связывается с ними, открывает цепь ДНК и синтезирует молекулу РНК, используя одну из этих комплементарных цепей ДНК в качестве матрицы или шаблон, даже если он встречает другую специальную последовательность остановки.
Транскрипция у эукариот (процесс)
- На что похожи гены эукариот?
Ген состоит из последовательности ДНК, характеристики которой определяют его функцию, поскольку порядок нуклеотидов в указанной последовательности определяет ее транскрипцию и последующую трансляцию (в случае тех, которые кодируют белки).
Когда ген транскрибируется, то есть когда его информация копируется в форме РНК, результатом может быть некодирующая РНК (кРНК), которая выполняет прямые функции в регуляции экспрессии генов, в передаче сигналов клеток и т. Д. или это может быть информационная РНК (мРНК), которая затем будет транслироваться в аминокислотную последовательность в пептиде.
Наличие у гена функционального продукта в виде РНК или белка зависит от определенных элементов или участков, присутствующих в его последовательности.
Каждый ген имеет на концах регуляторные последовательности:
- те последовательности, которые наиболее близки к 3 'области генов, обычно являются последовательностями терминатора, которые указывают полимеразе, что она должна останавливать и прекращать транскрипцию (или репликацию, в зависимости от обстоятельств)
Промоторная область делится на дистальную и проксимальную в зависимости от ее близости к кодирующей области. Он находится на 5'-конце гена и является сайтом, который распознает фермент РНК-полимеразу и другие белки, чтобы инициировать транскрипцию ДНК в РНК.
В проксимальной части промоторной области могут связываться факторы транскрипции, которые обладают способностью изменять сродство фермента к транскрибируемой последовательности, таким образом, они несут ответственность за положительную или отрицательную регуляцию транскрипции генов.
- Кто отвечает за транскрипцию?
Каким бы ни был организм, транскрипция осуществляется группой ферментов, называемых РНК-полимеразами, которые, подобно ферментам, отвечающим за репликацию ДНК, когда клетка собирается делиться, специализируются на синтезе цепи РНК. от одной из нитей ДНК транскрибируемого гена.
РНК-полимеразы - это большие ферментные комплексы, состоящие из многих субъединиц. Есть разные виды:
- РНК-полимераза II (Pol II): транскрибирует гены, кодирующие белки, и продуцирует микро-РНК.
- РНК-полимераза III (Pol III): продуцируется транспортными РНК, используемыми во время трансляции, а также РНК, соответствующей малой субъединице рибосомы.
- РНК-полимеразы IV и V (Pol IV и Pol V): они типичны для растений и отвечают за транскрипцию малых интерферирующих РНК.
- Каков процесс?
Генетическая транскрипция - это процесс, который можно разделить на три фазы: инициация, удлинение и завершение.
Инициация
РНК-полимераза (скажем, РНК-полимераза II) связывается с последовательностью промоторной области, которая состоит из участка длиной 6-10 пар оснований на 5'-конце гена, обычно на расстоянии примерно 35 пар оснований. сайта начала транскрипции.
Удлинение
Набор, состоящий из нити ДНК, РНК-полимеразы и формирующейся нити РНК, известен как транскрипционный пузырь или комплекс.
Прекращение
Транскрипция в прокариотах (процесс)
Таким образом, повышение цитозольной концентрации данного белка у прокариот происходит значительно быстрее, чем у эукариот, поскольку процессы транскрипции и трансляции происходят в одном и том же компартменте.
- Что такое прокариотические гены?
В этом смысле важно упомянуть, что в целом прокариотические гены всегда включены по умолчанию.
Разница в транскрипции между прокариотами и эукариотами заключается в том, что обычно информационные РНК эукариот являются моноцистронными, то есть каждая из них содержит информацию для синтеза одного белка, тогда как у прокариот они могут быть моноцистронными или полицистронными, где только одна МРНК может содержать информацию о двух или более белках.
Таким образом, хорошо известно, что прокариотические гены, кодирующие белки со сходными метаболическими функциями, например, обнаруживаются в группах, известных как опероны, которые одновременно транскрибируются в единую молекулярную форму матричной РНК.
Прокариотические гены плотно упакованы, без множества некодирующих областей между ними, поэтому после транскрибирования в молекулы линейной информационной РНК они могут быть немедленно транслированы в белок (эукариотические мРНК часто нуждаются в дальнейшей обработке).
- Как работает прокариотическая РНК-полимераза?
Прокариотические организмы, такие как бактерии, например, используют один и тот же фермент РНК-полимеразу для транскрипции всех своих генов, то есть тех, которые кодируют рибосомные субъединицы, и тех, которые кодируют различные клеточные белки.
В бактериях Кишечная палочка РНК-полимераза состоит из 5 полипептидных субъединиц, две из которых идентичны. Субъединицы α, α, β, β 'составляют центральную часть фермента и собираются и разбираются во время каждого события транскрипции.
Субъединицы α - это субъединицы, которые обеспечивают соединение между ДНК и ферментом; субъединица β связывается с трифосфатными рибонуклеотидами, которые будут полимеризоваться в соответствии с ДНК-матрицей в формирующейся молекуле мРНК, а β'-субъединица связывается с указанной цепью матричной ДНК.
Пятая субъединица, известная как σ участвует в инициации транскрипции и придает полимеразе специфичность.
- Каков процесс?
Транскрипция у прокариот очень похожа на транскрипцию эукариот (она также делится на инициацию, элонгацию и терминацию) с некоторыми различиями в идентичности промоторных областей и факторов транскрипции, необходимых для РНК-полимеразы. выполнять свои функции.
Инициация
Это зависит от σ-субъединицы РНК-полимеразы, поскольку она опосредует взаимодействие между ДНК и ферментом, делая его способным распознавать промоторные последовательности. Инициация заканчивается, когда выделяются некоторые абортивные транскрипты длиной около 10 нуклеотидов.
Удлинение
Когда σ-субъединица отделяется от фермента, начинается фаза элонгации, которая состоит из синтеза молекулы мРНК в направлении 5'-3 '(примерно 40 нуклеотидов в секунду).
Прекращение
Терминация у прокариот зависит от двух разных типов сигналов, она может быть Rho-зависимой и Rho-независимой.
Rho-независимая терминация контролируется специфическими последовательностями гена, обычно богатыми гуанин-цитозиновыми (GC) повторами.
Транскрипцией называется биосинтез РНК на матрице ДНК. Это первая стадия реализации генетической информации, в процессе которой определенные участки нуклеотидной последовательности ДНК “переписываются” в комплементарные одноцепочечные молекулы РНК. В результате транскрипции образуются мРНК, кодирующие аминокислотные последовательности белков, а также тРНК, рРНК и другие виды РНК, выполняющие структурные, регуляторные и каталитические функции. В основе транскрипции лежит принцип комплементарности азотистых оснований полинуклеотидных цепей ДНК и РНК. Процесс транскрипции осуществляется с участием соответствующих ферментов – РНК-полимераз. Кроме того, для осуществления процесса необходимы белки-регуляторы.
У подавляющего большинства организмов ДНК представляет собой двухцепочечные молекулы, составленные из комплементарных (но не идентичных) цепей, транскрипции подвергается обычно одна из них, носящая название матричной цепи:
ДНК 5´-АTGATTGGGGCTCTA-3´ смысловая цепь
3´-TACTAACCCCGAGAT-5´ матричная цепь
Различные РНК обычно транскрибируются с одной из цепей ДНК, однако у некторых геномов (например бактериофаг Т4) матрицей могут служить разные цепи ДНК.
Синтез РНК (транскрипция) осуществляется ферментами ДНК-зависимыми РНК-полимеразами. РНК-полимеразы, так же как и ДНК-полимеразы используют в качестве субстратов нуклеозидтрифосфаты и активны только в присутствии ионов Mg 2+ . В отличие от ДНК-полимераз РНК-полимеразы не нуждаются в праймере. Рост цепи РНК происходит путем последовательного присоединения рибонуклеотид-5´-монофосфатов к 3´-гидроксильной группе рибозы предшествующего нуклеотида (т.е. в направлении 5´-3´). Последовательность нуклеотидов в синтезируемом РНК-транскрипте определяется комплементарными взаимодействиями азотистых оснований, используемых для синтеза рибонуклеотидов, с азотистыми основаниями матричной цепи ДНК.
У всех без исключения организмов процессу транскрипции подвергается единовременно не вся молекула, а только определенные ее участки – транскриптоны. Они ограничены двумя последовательностями, которые называются промотором (зона начала транскрипции)и терминатором (зона остановки транскрипции). Транскриптоны бактерий называют оперонами. Опероны обычно включают в себя нуклеотидные последовательности, кодирующие структуру нескольких белков, называемые цистронами или структурными генами. Поэтому синтезируемая на оперонах бактерий мРНК является полицистронной и используется для синтеза нескольких белков (обычно это ферменты одного метаболитического пути).
Бактериальные РНК-полимеразы – сложные белки, состоящие из нескольких субъединиц. Наиболее изученный фермент кишечной палочки содержит 5 субъдиниц: две субъединицы α и по одной β, β´, ώ (основа фермента – кор). После присоединения к кору еще одной субъединицы – σ-фактора – образуется холофермент РНК-полимераза. Установлено, что при присоединении к ДНК фермент “закрывает” 60 пар нуклеотидов.
Процесс транскрипции состоит их 3 этапов: инициации, элонгации и терминации.
Функцией холофермента РНК-полимеразы является связывание с промотором и инициация транскрипции. Кор-фермент (α,α ,β,β´) осуществляет элонгацию, т.е. собственно синтез РНК. Для терминации транскрипции в некоторых случаях требуется присоединение к кор-ферменту фактора терминации – ρ-фактора.
Инициация транскрипции происходит в зоне промотора – особой нуклеотидной последовательности ДНК, которая содержит сигнал начала транскрипции. В большинстве промоторов E.coli присутствуют две последовательности: последовательность 5´-ТАТАТТ-3´(расположена на расстоянии -10 нуклеотидов от точки начала транскрипции, носит название Прибнов-бокс) и последовательность 5´-TTGACA-3´(расположена на расстоянии -35 нуклеотидов от точки старта транскрипции).
Для инициации транскрипции необходимо присоединение к кор-ферменту σ-фактора с образованием холофермента. Основная функция σ-фактора состоит в том, чтобы обеспечить стабильное связывание РНК-полимеразы с промотором, а не с другими участками ДНК. Минимальный фермент имеет сродство к ДНК; в основе этого сродства лежит электростатическое взаимодействие между положительно заряженным белком и отрицательно заряженной ДНК. Случайная последовательность ДНК, которая связывается РНК-полимеразой, называется слабым участком связывания, а комплекс ДНК-фермент закрытым, так как ДНК находится в таком комплексе в двухцепочечной форме.
Сигма-фактор принципиально изменяет характер взаимодействия РНК-полимеразы с ДНК. У холофермента сродство к любым последовательностям ДНК, кроме промотора, резко снижено. Сигма-фактор сообщает ферменту способность к узнавать специфические участки связывания (промоторы), путем присоединения к последовательности -35.
Взаимодействие холофермента с промотором начинается с образования двойного закрытого комплекса (включает 2 компонента -ДНК и фермент, ДНК имеет двухцепочечную форму). Двойной закрытый комплекс затем переходит в двойной открытый (в нем небольшой фрагмент ДНК расплетается в области Прибнов-бокса) комплекс. Следующим шагом является включение двух первых нуклеотидов РНК и образование фосфодиэфирной связи между ними. Это приводит к формированию тройного открытого комплекса между ферментом, ДНК и синтезирующейся РНК. После образования тройного комплекса инициация заканчивается и сигма-фактор отделяется (он используется клеткой многократно- т.н. цикл сигма-фактора).
Элонгацию осуществляет минимальный фермент, в котором сигма-фактор замещается фактором элонгации (например белок NusA E.coli). Перемещаясь вдоль двойной спирали ДНК РНК-полимераза непрерывно раскручивает спираль впереди того участка, где происходит синтез РНК. Затем фермент вновь закручивает ДНК позади участка полимеризации и готовая часть транскрипта выводится из комплекса через особый канал, свойственный РНК-полимеразе. Скорость синтеза РНК у бактерий составляет примерно 30 нуклеотидов в секунду.
Терминация транскрипции определяется особой нуклеотидной последовательностью в ДНК, расположенной в зоне терминатора (стоп-сигнал). В стоп-сигнале имеется GC-богатый участок, обладающий центральной симметрией и представляющий собой самокомплементарную нуклеотидную последовательность (инвертированные повторы).Такая последовательность называется палиндромом:
Копии повтора, находящиеся в одной и той же цепи ДНК взаимно комплементарны, если их прочитывать в противоположных направлениях. При образовании комплементаных пар возможно возникновение структуры шпильки в РНК или структуры креста в ДНК.
За палиндромом часто распологается последовательностьиз 4-8 адениловых нуклеотидов (ро-независимые терминаторы). Транскрипция в GC-богатой области приводит к образованию в РНК-транскрипте устойчивого элемента вторичной структуры – шпильки. Такая шпилька нарушает прочность связи в комплексе ДНК-РНК, кроме того, транскрипция олиго(А)- последовательности приводит к образованию дуплекса из очень непрочных пар A-U, что также приодит к разрушению комплекса ДНК-РНК. Таким образом, сразу после образования шпильки происходит высвобождение новосинтезированной РНК, отделение РНК-полимеразы и восстановлению двуспиральной структуры ДНК.
Часть терминаторов узнается РНК-полимеразой только с помощью белкового фактора терминации ρ(ро)-фактора (ро-зависимые терминаторы). Он присоединяется к продукту непосредственно перед терминатором. Ро-фактор обладает АТФазной активностью, во время его работы высвобождается энергия, которая изменяет его конформацию, что, вероятно, способствует вытеснению РНК из транскрипционного комплекса.
Регуляция транскрипции
Бактериям необходимо уметь быстро отвечать на изменения в окружающей среде. Их выживаемость зависит от способности переключать метаболизм с одного субстрата на другой, поскольку в условиях их обитания поступление питательных веществ может постоянно меняться. Одноклеточные эукариоты способны проявлять такую же зависимость от непрерывно меняющихся условий среды. Более сложно организованные многоклеточные организмы имеют более постоянный набор метаболитических путей и могут не отвечать на изменение внешних условий перестройкой своего метаболизма.
Легкая приспособляемость – большое преимущество бактерий. Однако при этом им необходима экономичность, поскольку бактерия, которая в ответ на изменение условий окружающей среды использует пути, требующие больших энергетических затрат, может оказаться в невыгодном положении. Очевидно, что энергетически невыгодно производить все ферменты метаболитических путей, если они не используются в данный момент из-за отсутствия субстрата. Действительно доказано, что бактерии не синтезируют ферментов того или иного пути в отсутствие соответствующего субстрата, но способны в любое время начать их синтез при появлении такового.
Этим объясняются главные особенности организации бактериальных генов – их объединение в кластеры таким образом, что все ферменты, необходимые для осуществления определенного пути биосинтеза, детерминируются генами, сцепленными друг с другом. Вся группа генов транскрибируется в одну полицистронную мРНК, которая последовательно транслируется рибосомами с образованием каждого их белков.
Транскрипция у прокариот регулируется преимущественно на стадии инициации и связана с деятельностью регуляторных белков – активаторов и репрессоров трансткрипции. Наиболее изученными оперонами бактерий являются лактозный и триптофановый опероны.
Существует ряд экспериментальных данных, свидетельствующих о том, что транскрипция осуществляется в так называемых транскрипционных фабриках: огромных, по некоторым оценкам, до 10 МДа комплексах, которые содержат около 8 РНК-полимераз II и компоненты последующего процессинга и сплайсинга, а также пруф-ридинга новосинтезированного транскрипта. В ядре клетки происходит постоянный обмен между пулами растворимой и задействованной РНК-полимеразы. Активная РНК-полимераза задействована в таком комплексе, который в свою очередь является структурной организовывающей компактизацию хроматина единицей. Последние данные. свидетельствуют о том, что транскрипционные фабрики существуют и в отсутствие транскрипции, они фиксированы в клетке (пока не ясно, взаимодействуют ли они с матриксом клетки или нет) и представляют собой независимый ядерный субкомпартмент. Попытки выделить белковый функциональный комплекс транскрипционной фабрики пока не привели к успеху ввиду его огромных размеров и низкой растворимости.
РНК-полимеразы эукариот
У эукариот имеется 3 типа РНК-полимераз (не считая митохондриальной и хлоропластной):
РНК полимеразаI - синтезирует в ядрышках рибосомные RNA (18S и 28S рРНК, кроме 5S);
РНК-полимеразаII - синтезирует mRNA и некоторых sRNA;
РНК-полимеразаIII - синтезирует tRNA, sRNA, 5S rRNA.
RNA-полимеразы эукариот отличаются: количеством субъединиц – 2 большие (120-220кДа) и до 8 малых (10-100кДа), потребностью в ионах Mg и Mn, чувствительностью к – амонитину - токсину бледной поганки - пептиду включающему D-аминокислоты: polI - устойчива, polII - ингибируется при концентрации 10-8М, polIII - при 10-6М амонитина. РНК-полимеразы I,II,III кодируются в ядре. Большие субъединицы гомологичны β и β`-субъединицам эубактерий.
РНК-полимераза I
РНК-полимераза II
PolII Человека содержит более 10 субъединиц, слабо ассоциирующих друг с другом. Некоторые из них принадлежат к основным факторам транскрипции (GTF).
Белки holo-фермента PolII дрожжей [Патрушев, 2000].
Pol II - РНК-Полимеразная активность, взаимодействует с множеством общих и тканеспецифических факторов транскрипции, участвует в выборе точки инициации транскрипции.
TFIIB - Связывает Pol II и TBP на промоторе, участвует в выборе точки инициации транскрипции
TFIIF - Взаимодействует с Pol II, стимулирует элонгацию транскрипции Pol II, компонент субкомплекса SRB/медиатор
TFIIH - Активность ДНК-зависимой ATPазы, ДНК-геликазная активность, обладает активностью CTD-киназы
SRB2, SRB5 - Участвуют в образовании инициационного комплекса, стимулируют базальный и индуцированный синтез РНК,
взаимодействуют с TBP, компоненты субкомплекса SRB/медиатор
GAL11/SPT13 - Участвуют в образовании инициационного комплекса, стимулируют базальный и индуцированный синтез РНК,
компоненты субкомплекса SRB/медиатор, предположительно взаимодействуют с активаторами транскрипции
SUG1 - Компонент субкомплекса SRB/медиатор, предположительно взаимодействует с активаторами транскрипции
SRB4, SRB6, SRB7, SRB8, SRB9, SRB10, SRB11 - Компоненты субкомплекса SRB/медиатор, предположительно
взаимодействуют с CTD-доменом Pol II
РНК-полимераза III
Факторы транскрипции
Инициация
Инициация транскрипции происходит на кэп-сайте кодирующем первый нуклеодид первого экзона мРНК.
ТАТА-бокс локализуется в 25-30 пн выше кэп-сайта связывая РНК-полимеразу перед кэп-сайтом. Промотор - примерно 200 пн выше кэп-сайта. Энхансеры обычно имеют длину 100-200 пн.
Элонгация
Терминация
Терминация на сайте полиаденилирования.
Вновь синтезированная РНК генов связывается с ядерными белками - информомерами, подвергается различным посттранскрипционным модификациям и транспортируется из ядра (см. обзор Процессинг) для последующей трансляции (см. обзор Трансляция).
РНК-полимераза E.coli
РНК-полимераза E.coli осуществляет транскрипцию всех бактериальных генов и состоит из нескольких субъединиц: α-35кДа, β‘-165кДа, β-155кДа, σ-чаще 70кДа (σ70). РНК-полимераза состава ααββ’σ70 называется holo-фермент (Еσ70), состава ααββ’- core-фермент (E).
σ - сменный фактор специфичности, который диссоциирует после инициации транскрипции. Элонгация и терминация осуществляется core-ферментом. У Е.coli ~10 видов σ-субъединиц. Транскрипция генов теплового шока, оперонов gln или nif осуществляется σ54 в составе holo-фермента Eσ54 (54 кДа).
Все субъединицы заряжены отрицательно: σ>α>β>β’ – расположены по убыванию заряда. В каждой субъединице имеется кластер (+)-заряженных участков, которыми они связываются с ДНК. Наибольшее число кластеров у – β’, который участвует в связывании фермента с ДНК, β-субъединица содержит активные центры - инициации и элонгации, α-субъединицы обеспечивают правильное взаимодействие фермента с промоторами. Рифампицин – блокирует инициацию, стрептолидигин – блокирует элонгацию, что говорит о разнесении активных центров в РНК-полимеразе.
Узнавание и связывание RNA-pol с промотором осуществляется holo-ферментом
Одновременно в клетке присутствует около 7000 молекул РНК-полимеразы. Только holo-фермент обладает высоким сродством к специфической последовательности нуклеотидов - промотору, сродство к остальным случайным последовательностям ДНК у него снижено в 10000 раз. У core-фермента одинаковое сродство к любой последовательности нуклеотидов.
Сам по себе сигма - фактор обладает наименьшим сродством к ДНК по сравнению с другими субьединицами РНК-полимеразы, однако он придает holo-ферменту такую конформацию, которая обладает повышенным сродством к промотору.
Стадии узнавания и связывания, а также инициации осуществляются holo-ферментом. Элонгация и терминация осуществляются core-ферментом.
Две α субъединицы - каркас РНК-полимеразы. К ним крепятся остальные субъединицы.
β' - субъединица отвечает за прочное связывание с ДНК за счет кластера положительно заряженных аминокислот.
В β - субъединице находятся два каталитических центра. Один отвечает за инициацию, а другой - за элонгацию. Один центр работает в holo-, а другой - в core- ферменте.
Инициация транскрипции
РНК-полимераза Ecoli узнает два 6н разделенных 25н
Элонгация транскрипции
Терминация транскрипции
Регуляция транскрипции
Схема негативной индукции Жакоба и Моно
Lac-оперон E. coli содержит 3 гена, отвечающие за образование белков, участвующих в переносе в клетку дисахарида лактозы и в ее расщеплении.
Z-β - галактозидаза (расщепляет лактозу на глюкозу и галактозу).
Y-β- галактозидпермеаза (переносит лактозу через мембрану клетки).
А - тиогалактозидтрансацетилаза (ацетилирует галактозу).
В отсутствие в клетке лактозы lac-оперон выключен. Активный белок - репрессор, кодируемый в моноцистронном опероне (LacI) , не имеющем оператора, связан с оператором lac-оперона. Поскольку оператор перекрывается с промотором, даже посадка РНК-полимеразы на промотор невозможна.
Как только некоторое количество лактозы попадает в клетку, две молекулы субстрата (лактозы) взаимодействуют с белком - репрессором, изменяют его конформацию - и он теряеет сродство к оператору.
Тут же начинается транскрипция lac-оперона и трансляция образующейся mРНК; три синтезируемых белка участвуют в утилизации лактозы.
Когда вся лактоза переработана, очередная порция репрессора, свободного от лактозы, выключает lac-оперон.
Схема позитивной индукции
В Аra-опероне E. сoli 3 цистрона, которые кодируют ферменты, расщепляющие сахар арабинозу. В норме оперон закрыт. Белок - репрессор связан с оператором.
Когда в клетку попадает арабиноза, она взаимодействует с белком - репрессором. Белок - репрессор меняет конформацию и превращается из репрессора в активатор, взаимодейсивующий с промотором и облегчающий посадку РНК-полимеразы на промотор.
Эта схема регуляции называется позитивной индукцией, поскольку контролирующий элемент - белок - активатор "включает" работу оперона.
Схема позитивной репрессии
В опероне синтеза рибофлавина у Вacilus subtilis располагаются цистроны ферментов синтеза рибофлавина. Есть белок-активатор, обеспечивающий посадку РНК-полимеразы на промотор. В норме оперон открыт. Образуется N молекул рибофлавина.
N+1-ая молекула (лишняя) взаимодействует с активатором и он теряет способность активировать посадку РНК-полимеразы на промотор.
Позитивная репрессия, поскольку в регуляции участвует белок - активатор, а сама регуляция заключается в выключении транскрипции.
Схема негативной репрессии
В опероне синтеза триптофана у E. сoli имеется 5 цистронов, которые кодируют ферменты последовательной цепи реакций синтеза триптофана. В норме оперон включен. Белок - репрессор неактивен (в форме апо-репрессора), он не способен садиться на оператор.
Клетке нужно N молекул триптофана. N+1-ая молекула взаимодействует с апо-репрессором. Он меняет конформацию, садится на оператор и синтез РНК прекращается.
Схема регуляции - негативная репрессия, потому что белок репрессор "выключает" оперон.
Позитивный контроль работы lac-оперона
Lac-оперон, подчиняющийся схеме негативной индукции, имеет и позитивный контроль. цАМФ образуется из АТФ ферментом аденилатциклазой. Фосфодиэстераза превращает цАМФ в АМФ. Глюкоза активирует второй и инактивирует первый фермент. Чем больше в клетке глюкозы, тем меньше цАМФ.
Если нет глюкозы, то цАМФ соединяется с белком катаболической репрессии (САР) и образуется комплекс САР•цАМФ, активирующий посадку РНК-полимеразы на промотор. В присутствии лактозы lac-оперон включается и работает. Если же в клетке есть еще и глюкоза (более экономичный источнок энергии), то нет цАМФ - и активатор не образуется, lac-оперон работает слабо, без дополнительной индукции.
И транскрипция, и трансляция относятся к матричным биосинтезам. Матричным биосинтезом называется синтез биополимеров (нуклеиновых кислот, белков) на матрице - нуклеиновой кислоте ДНК или РНК. Процессы матричного биосинтеза относятся к пластическому обмену: клетка расходует энергию АТФ.
Матричный синтез можно представить как создание копии исходной информации на несколько другом или новом "генетическом языке". Скоро вы все поймете - мы научимся достраивать по одной цепи ДНК другую, переводить РНК в ДНК и наоборот, синтезировать белок с иРНК на рибосоме. В данной статье вас ждут подробные примеры решения задач, генетический словарик пригодится - перерисуйте его себе :)

Возьмем 3 абстрактных нуклеотида ДНК (триплет) - АТЦ. На иРНК этим нуклеотидам будут соответствовать - УАГ (кодон иРНК). тРНК, комплементарная иРНК, будет иметь запись - АУЦ (антикодон тРНК). Три нуклеотида в зависимости от своего расположения будут называться по-разному: триплет, кодон и антикодон. Обратите на это особое внимание.
Репликация ДНК - удвоение, дупликация (лат. replicatio — возобновление, лат. duplicatio - удвоение)
Процесс синтеза дочерней молекулы ДНК по матрице родительской ДНК. Нуклеотиды достраивает фермент ДНК-полимераза по принципу комплементарности. Переводя действия данного фермента на наш язык, он следует следующему правилу: А (аденин) переводит в Т (тимин), Г (гуанин) - в Ц (цитозин).
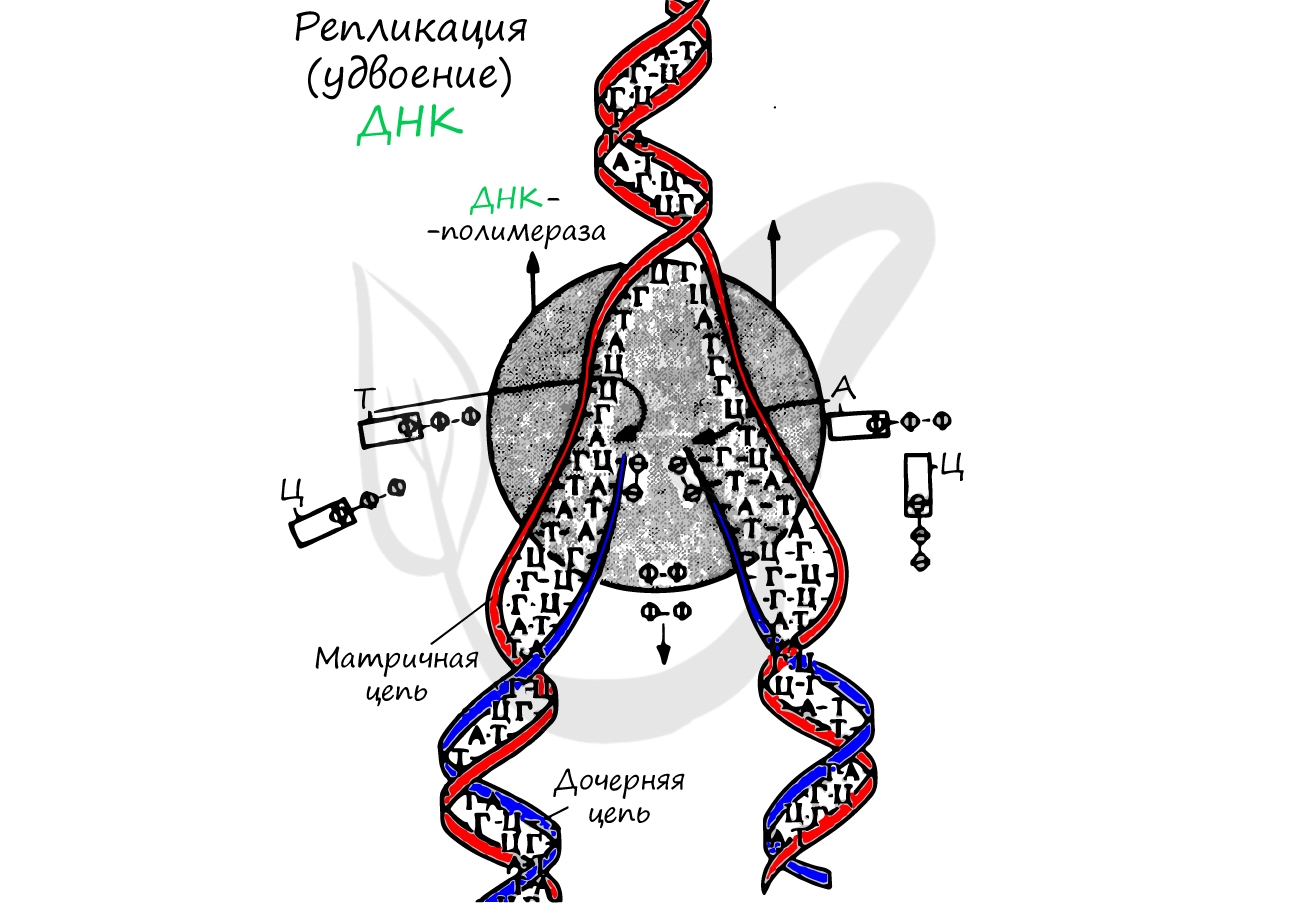
Удвоение ДНК происходит в синтетическом периоде интерфазы. При этом общее число хромосом не меняется, однако каждая из них содержит к началу деления две молекулы ДНК: это необходимо для равномерного распределения генетического материала между дочерними клетками.
Транскрипция (лат. transcriptio — переписывание)
Транскрипция представляет собой синтез информационной РНК (иРНК) по матрице ДНК. Несомненно, транскрипция происходит в соответствии с принципом комплементарности азотистых оснований: А - У, Т - А, Г - Ц, Ц - Г (загляните в "генетический словарик" выше).
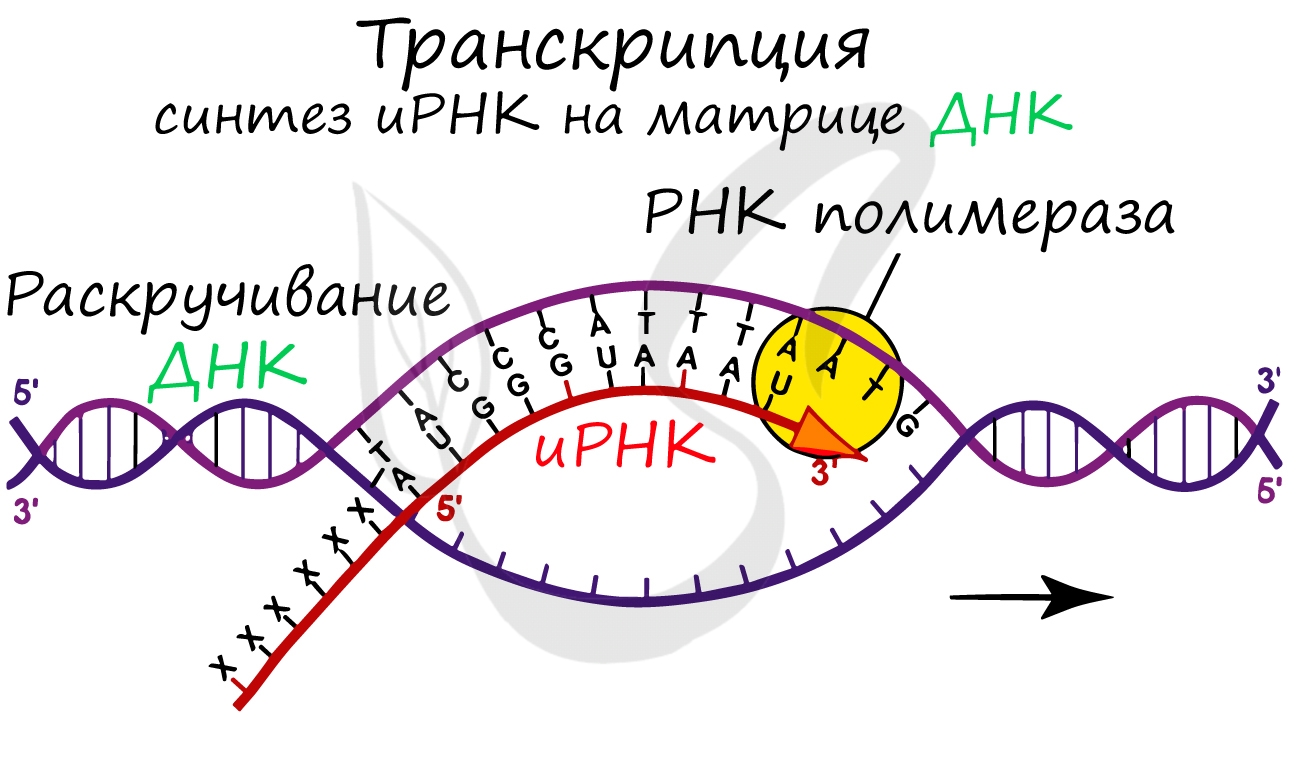
До начала непосредственно транскрипции происходит подготовительный этап: фермент РНК-полимераза узнает особый участок молекулы ДНК - промотор и связывается с ним. После связывания с промотором происходит раскручивание молекулы ДНК, состоящей из двух цепей: транскрибируемой и смысловой. В процессе транскрипции принимает участие только транскрибируемая цепь ДНК.
-
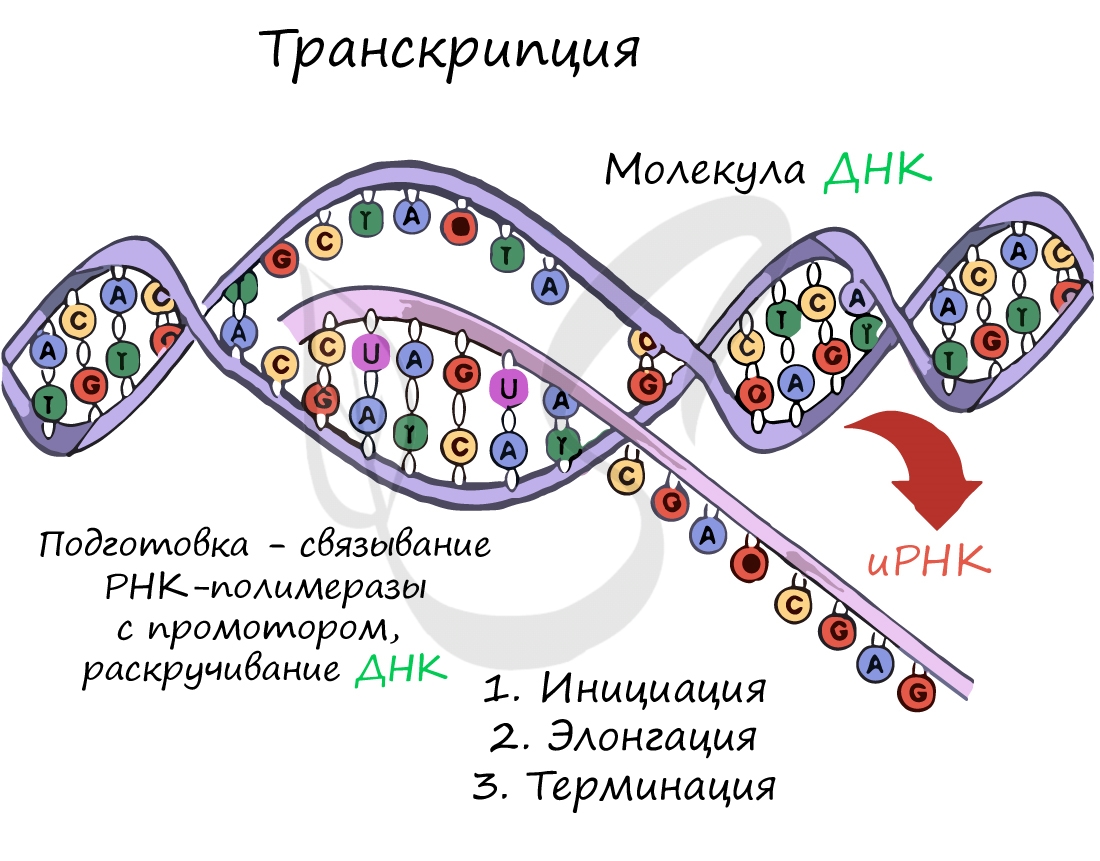
Инициация (лат. injicere — вызывать)
Образуется несколько начальных кодонов иРНК.
Нити ДНК последовательно расплетаются, освобождая место для передвигающейся РНК-полимеразы. Молекула иРНК быстро растет.
Достигая особого участка цепи ДНК - терминатора, РНК-полимераза получает сигнал к прекращению синтеза иРНК. Транскрипция завершается. Синтезированная иРНК направляется из ядра в цитоплазму.
Трансляция (от лат. translatio — перенос, перемещение)
Куда же отправляется новосинтезированная иРНК в процессе транскрипции? На следующую ступень - в процесс трансляции. Он заключается в синтезе белка на рибосоме по матрице иРНК. Последовательность кодонов иРНК переводится в последовательность аминокислот.
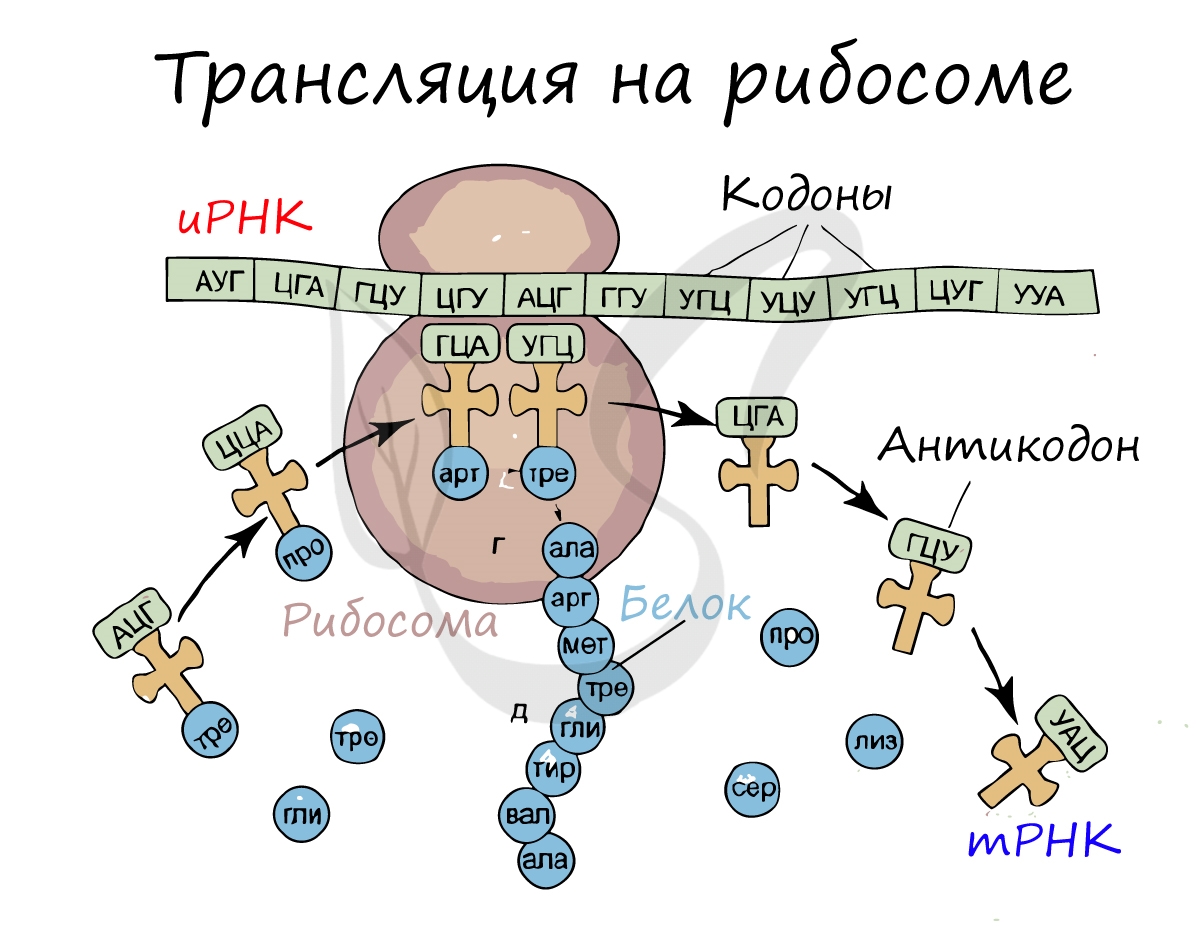
Информационная РНК (иРНК, синоним - мРНК (матричная РНК)) присоединяется к рибосоме, состоящей из двух субъединиц. Замечу, что вне процесса трансляции субъединицы рибосом находятся в разобранном состоянии.
Первый кодон иРНК, старт-кодон, АУГ оказывается в центре рибосомы, после чего тРНК приносит аминокислоту, соответствующую кодону АУГ - метионин.
Рибосома делает шаг, и иРНК продвигается на один кодон: такое в фазу элонгации происходит десятки тысяч раз. Молекулы тРНК приносят новые аминокислоты, соответствующие кодонам иРНК. Аминокислоты соединяются друг с другом: между ними образуются пептидные связи, молекула белка растет.
Доставка нужных аминокислот осуществляется благодаря точному соответствию 3 нуклеотидов (кодона) иРНК 3 нуклеотидам (антикодону) тРНК. Язык перевода между иРНК и тРНК выглядит как: А (аденин) - У (урацил), Г (гуанин) - Ц (цитозин). В основе этого также лежит принцип комплементарности.
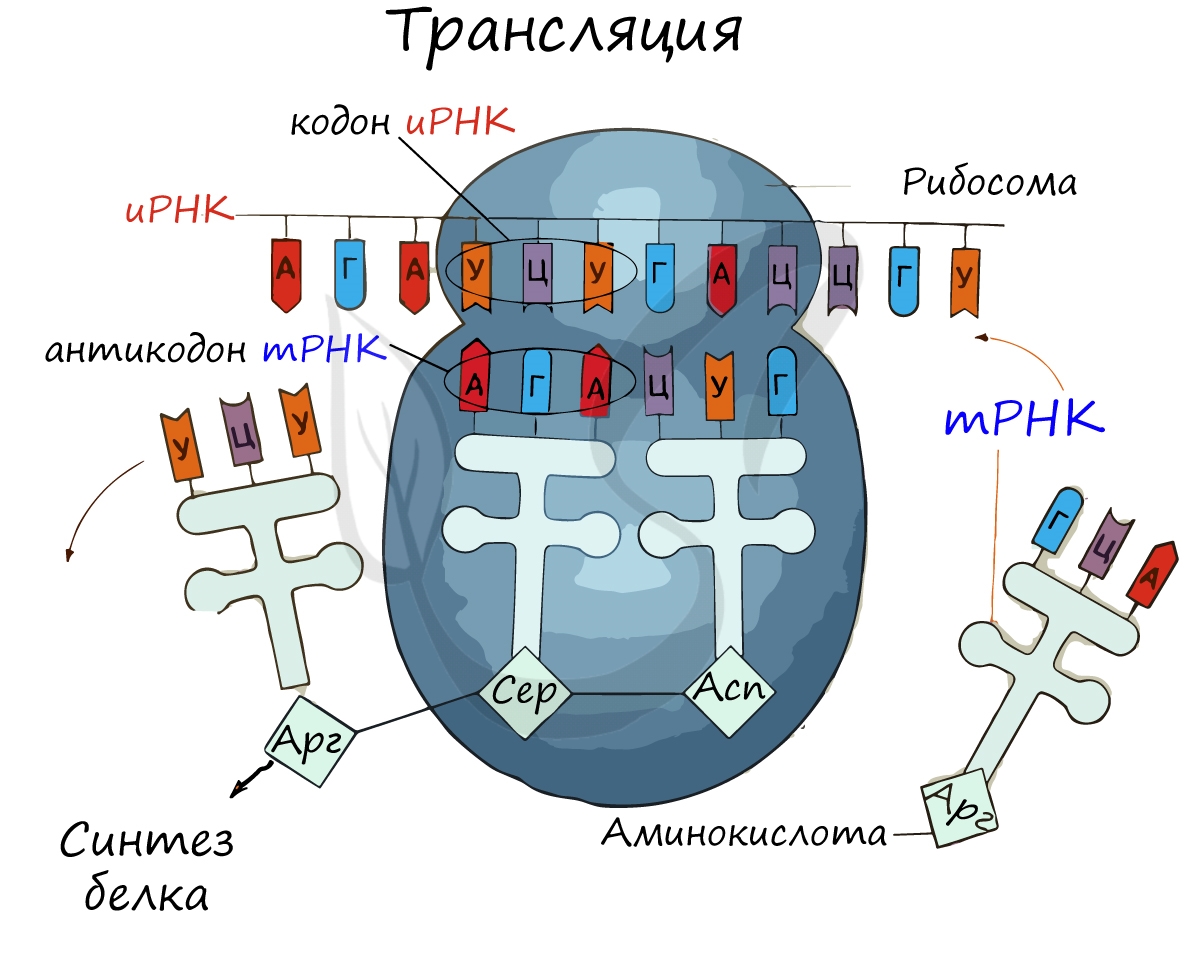
Движение рибосомы вдоль молекулы иРНК называется транслокация. Нередко в клетке множество рибосом садятся на одну молекулу иРНК одновременно - образующаяся при этом структура называется полирибосома (полисома). В результате происходит одновременный синтез множества одинаковых белков.
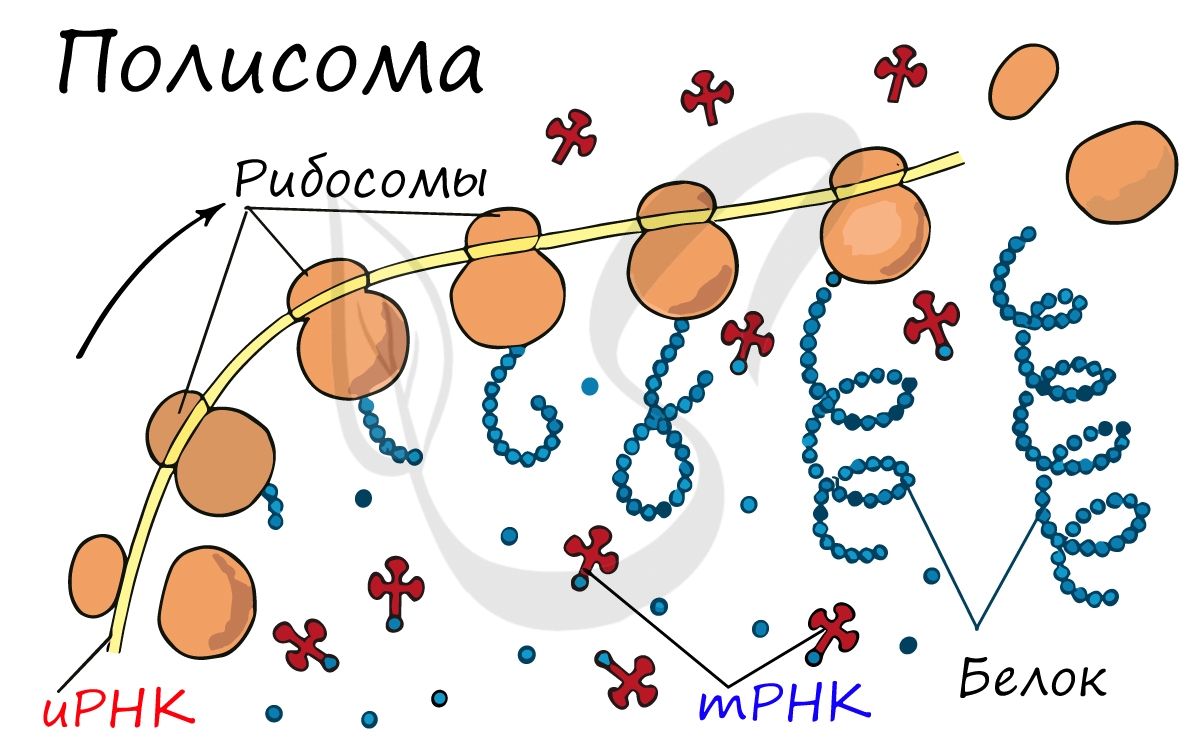
Синтез белка - полипептидной цепи из аминокислот - в определенный момент завершатся. Сигналом к этому служит попадание в центр рибосомы одного из так называемых стоп-кодонов: УАГ, УГА, УАА. Они относятся к нонсенс-кодонам (бессмысленным), которые не кодируют ни одну аминокислоту. Их функция - завершить синтез белка.
Существует специальная таблица для перевода кодонов иРНК в аминокислоты. Пользоваться ей очень просто, если вы запомните, что кодон состоит из 3 нуклеотидов. Первый нуклеотид берется из левого вертикального столбика, второй - из верхнего горизонтального, третий - из правого вертикального столбика. На пересечении всех линий, идущих от них, и находится нужная вам аминокислота :)
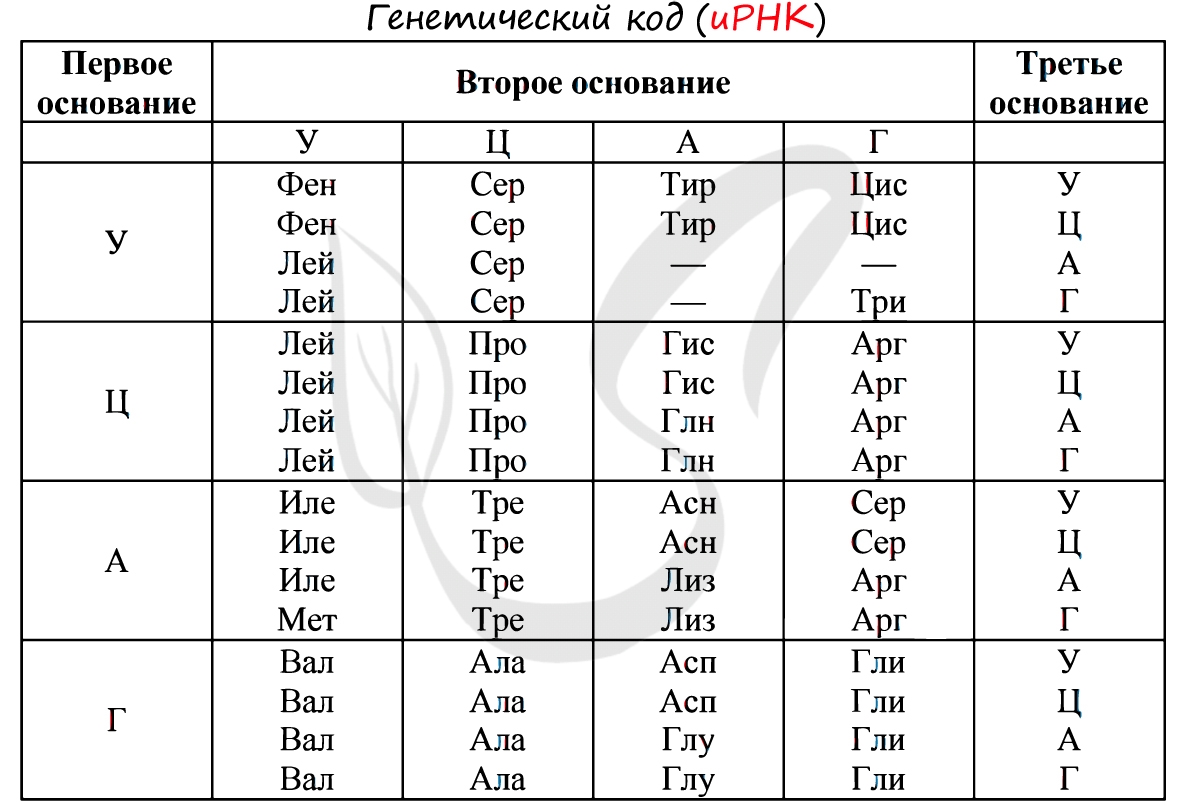
Давайте потренируемся: кодону ЦАЦ соответствует аминокислота Гис, кодону ЦАА - Глн. Попробуйте самостоятельно найти аминокислоты, которые кодируют кодоны ГЦУ, ААА, УАА.
Кодону ГЦУ соответствует аминокислота - Ала, ААА - Лиз. Напротив кодона УАА в таблице вы должны были обнаружить прочерк: это один из трех нонсенс-кодонов, завершающих синтез белка.
Примеры решения задачи №1
Без практики теория мертва, так что скорее решим задачи! В первых двух задачах будем пользоваться таблицей генетического кода (по иРНК), приведенной вверху.
"Фрагмент цепи ДНК имеет следующую последовательность нуклеотидов: ЦГА-ТГГ-ТЦЦ-ГАЦ. Определите последовательность нуклеотидов во второй цепочке ДНК, последовательность нуклеотидов на иРНК, антикодоны соответствующих тРНК и аминокислотную последовательность соответствующего фрагмента молекулы белка, используя таблицу генетического кода"

По принципу комплементарности мы нашли вторую цепочку ДНК: ГЦТ-АЦЦ-АГГ-ЦТГ. Мы использовали следующие правила при нахождении второй нити ДНК: А-Т, Т-А, Г-Ц, Ц-Г.
Вернемся к первой цепочке, и именно от нее пойдем к иРНК: ГЦУ-АЦЦ-АГГ-ЦУГ. Мы использовали следующие правила при переводе ДНК в иРНК: А-У, Т-А, Г-Ц, Ц-Г.
Зная последовательность нуклеотидов иРНК, легко найдем тРНК: ЦГА, УГГ, УЦЦ, ГАЦ. Мы использовали следующие правила перевода иРНК в тРНК: А-У, У-А, Г-Ц, Ц-Г. Обратите внимание, что антикодоны тРНК мы разделяем запятыми, в отличие кодонов иРНК. Это связано с тем, что тРНК представляют собой отдельные молекулы (в виде клеверного листа), а не линейную структуру (как ДНК, иРНК).
Пример решения задачи №2
"Известно, что все виды РНК синтезируются на ДНК-матрице. Фрагмент цепи ДНК, на которой синтезируется участок центральной петли тРНК, имеет следующую последовательность нуклеотидов: ТАГ-ЦАА-АЦГ-ГЦТ-АЦЦ. Установите нуклеотидную последовательность участка тРНК, который синтезируется на данном фрагменте, и аминокислоту, которую будет переносить эта тРНК в процессе биосинтеза белка, если третий триплет соответствует антикодону тРНК"

Обратите свое пристальное внимание на слова "Известно, что все виды РНК синтезируются на ДНК-матрице. Фрагмент цепи ДНК, на которой синтезируется участок центральной петли тРНК ". Эта фраза кардинально меняет ход решения задачи: мы получаем право напрямую и сразу синтезировать с ДНК фрагмент тРНК - другой подход здесь будет считаться ошибкой.
Итак, синтезируем напрямую с ДНК фрагмент молекулы тРНК: АУЦ-ГУУ-УГЦ-ЦГА-УГГ. Это не отдельные молекулы тРНК (как было в предыдущей задаче), поэтому не следует разделять их запятой - мы записываем их линейно через тире.
Третий триплет ДНК - АЦГ соответствует антикодону тРНК - УГЦ. Однако мы пользуемся таблицей генетического кода по иРНК, так что переведем антикодон тРНК - УГЦ в кодон иРНК - АЦГ. Теперь очевидно, что аминокислота кодируемая АЦГ - Тре.
Пример решения задачи №3
Длина фрагмента молекулы ДНК составляет 150 нуклеотидов. Найдите число триплетов ДНК, кодонов иРНК, антикодонов тРНК и аминокислот, соответствующих данному фрагменту. Известно, что аденин составляет 20% в данном фрагменте (двухцепочечной молекуле ДНК), найдите содержание в процентах остальных нуклеотидов.

Один триплет ДНК состоит из 3 нуклеотидов, следовательно, 150 нуклеотидов составляют 50 триплетов ДНК (150 / 3). Каждый триплет ДНК соответствует одному кодону иРНК, который в свою очередь соответствует одному антикодону тРНК - так что их тоже по 50.
По правилу Чаргаффа: количество аденина = количеству тимина, цитозина = гуанина. Аденина 20%, значит и тимина также 20%. 100% - (20%+20%) = 60% - столько приходится на оставшиеся цитозин и гуанин. Поскольку их процент содержания равен, то на каждый приходится по 30%.
Теперь мы украсили теорию практикой. Что может быть лучше при изучении новой темы? :)
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Читайте также: