
Транскрипция генетический код кратко
Обновлено: 02.07.2024
Биосинтез белка – ферментативный процесс синтеза белков в клетке. В нём участвуют три структурные элемента клетки – ядро, цитоплазма, рибосомы.
В ядре клетки в молекулах ДНК сохраняется информация о всех белках, которые в ней синтезируются, зашифрованная с помощью четырёхбуквенного кода.
Генетический код – это последовательность расположения нуклеотидов в молекуле ДНК, которая определяет последовательность аминокислот в молекуле белка.
Свойства генетического кода таковы:
Генетический код триплетный, то есть каждой аминокислоте соответствует свой кодовый триплет (кодон), состоящий из трёх расположенных рядом нуклеотидов.
Аминокислота цистеин кодируется триплетом А-Ц-А, валин – триплетом Ц-А-А.
Код не перекрывается, то есть нуклеотид не может входить в состав двух соседних триплетов.
Код вырожден, то есть одна аминокислота может кодироваться несколькими триплетами.
Аминокислота тирозин кодируется двумя триплетами.
Код не имеет запятых (разделительных знаков), считывание информации происходит тройками нуклеотидов.
Ген – участок молекулы ДНК, который характеризуется определённой последовательностью нуклеотидов и определяет синтез одногой полипептидной цепи.
Код является универсальным, то есть единым для всех живых организмов – от бактерий до человека. У всех организмов есть одни и те же 20 аминокислот, которые кодируются одними и теми же триплетами.
Этапы биосинтеза белка: транскрипция и трансляция
Структура любой белковой молекулы закодирована в ДНК, которая не участвует непосредственно в её синтезе. Она служит лишь матрицей для синтеза РНК.
Процесс биосинтеза белка происходит на рибосомах, которые расположены преимущественно в цитоплазме. Значит, для осуществления передачи к месту синтеза белка генетической информации из ДНК нужен посредник. Эту функцию выполняет иРНК.
Процесс синтеза молекулы иРНК на одной цепи молекулы ДНК на основании принципа комплементарности называется транскрипцией, или переписыванием.
Транскрипция происходит в ядре клетки.
Готовые работы на аналогичную тему
Процесс транскрипции осуществляется одновременно не на всей молекуле ДНК, а лишь на её небольшом участке, который отвечает определённому гену. При этом происходит раскручивание части двойной спирали ДНК и короткий участок одной из цепей оголяется – теперь он будет выполнять роль матрицы для синтеза иРНК.
Потом вдоль этой цепи двигается фермент РНК-полимераза, соединяющий нуклеотиды в цепь иРНК, которая удлиняется.
Транскрипция может одновременно происходить и на нескольких генах одной хромосомы и на генах разных хромосомах.
Образованная в результате иРНК содержит последовательность нуклеотидов, которая является точной копией последовательности нуклеотидов на матрице.
Если в молекуле ДНК есть азотистое основание цитозин, то в иРНК – гуанин и наоборот. Комплементарной парой в ДНК является аденин – тимин, а РНК вместо тимина содержит урацил.
На специальных генах синтезируются и два другие типа РНК – тРНК и рРНК.
Соединение тРНК с аминокислотами происходит в цитоплазме. Молекула тРНК формой напоминает листик клевера, на его верхушке расположен антикодон – триплет нуклеотидов, который кодирует аминокислоту, которую переносит данная тРНК.
Сколько видов аминокислот, столько существует и тРНК.
Поскольку много аминокислот могут кодироваться несколькими триплетами, то количество тРНК больше 20 (известно около 60 тРНК).
Соединение тРНК с аминокислотами происходит с участием ферментов. Молекулы тРНК транспортируют аминокислоты к рибосомам.
Трансляция – это процесс, в результате которого информация о структуре белка, записанная в иРНК в виде последовательности нуклеотидов, реализуется в виде последовательности аминокислот в молекуле белка, которая синтезируется.
Этот процесс осуществляется в рибосомах.

Каждая рибосома состоит из двух субъединиц – малой и большой. Молекула иРНК присоединяется к малой субъединице. В месте контакта рибосомы и иРН находятся 6 нуклеотидов (2 триплета). К одному из них всё время подходят из цитоплазмы тРНК с разными аминокислотами и касаются антикодоном кодона иРНК. Если триплеты кодона и антикодона оказываются комплементарными, между аминокислотой уже синтезированной части белка и аминокислотой, которая доставляется тРНК, возникает пептидная связь. Соединение аминокислот в молекулу белка осуществляется с участием фермента синтетазы. Молекула тРНК отдаёт аминокислоту и переходит в цитоплазму, а рибосома передвигается на один триплет нуклеотидов. Так последовательно синтезируется полипептидная цепь. Продолжается всё это до тех пор, пока рибосома не дойдёт к одному из трёх терминирующих кодонов: УАА, УАГ или УГА. После этого синтез белка прекращается.
Таким образом, последовательность кодонов иРНК определяет последовательность включения аминокислот в цепь белка. Синтезированные белки поступают в каналы эндоплазматического ретикулюма. Одна молекула белка в клетке синтезируется за 1 - 2 минуты.
Биосинтез белка — это ферментативный процесс синтеза белков в клетке, в котором принимают участие три структурных элемента клетки: ядро, цитоплазма и рибосомы.
Молекулы ДНК в ядре клетки сохраняют информацию обо всех белках, синтезирующихся в этой клетке. Эта информация находится в зашифрованном виде — шифруется 4-буквенным кодом.
Генетический код представляет собой последовательность расположения нуклеотидов в молекуле ДНК, определяющей последовательность аминокислот в молекуле белка.
Генетический код обладает следующими свойствами:
- он триплетный. У каждой аминокислоты есть свой кодовый триплет или кодон, в состав которого входят три нуклеотида, расположенные рядом.
К примеру, такая кислота как цистеин кодируется при помощи триплета А-Ц-А. В отношении валина — это Ц-А-А.
- код не перекрывается. Это значит, что в составе двух соседних триплетов нет того же нуклеотида.
- вырожденный код. Имеется в виду, что какая-либо аминокислота кодируется при помощи нескольких триплетов.
Если взять аминокислоту тирозин, то она кодируется при помощи двух триплетов.
- в коде не содержатся запятые, выполняющие функцию разделительных знаков, поэтому информация считывается тройками нуклеотидов.
УАГ, УАА, УГА — три несодержательных кодона, не кодирующие аминокислоты. Предполагается, что они выступают в качестве стоп-сигналов, благодаря которым происходит разделение генов в молекуле ДНК.
Ген — участок молекулы ДНК, для которого свойственна определенная последовательность нуклеотидов. Ген определяет синтез одной полипептидной цепи.
- универсальность кода. Он един для всех живых организмов, включая бактерий и человека. Все организмы содержат одинаковые 20 аминокислот, кодируемые одними и теми же триплетами.
Этапы биосинтеза белка: транскрипция и трансляция
Транскрипция белка
Этапы биосинтеза белка основаны на двух процессах: транскрипции и трансляции.
Самый популярный вопрос в рамках этой темы — где происходит синтез белка. И только потом разбираются с этапами синтеза белка (и схемой биосинтеза белка).
Любая белковая молекула имеет структуру, закодированную в ДНК. В ее синтезе эта ДНК не принимает непосредственного участия. Роль белковой молекулы — роль матрицы для синтеза РНК.
Далее охарактеризуем функции различных видов РНК в биосинтезе белка.
Где и как происходит биосинтез белка? Синтез белка происходит в, а точнее, синтез белка происходит на рибосомах — в основном они размещаются в цитоплазме. Поэтому, чтобы генетическая информация из ДНК передалась к месту, где белок синтезируется, необходим посредник.
Роль такого посредника играет иРНК.
Первый этап биосинтеза белка — транскрипция.
Транскрипция (переписывание) — процесс синтеза молекулы иРНК на одной цепи молекулы ДНК, в основе которого лежит принцип комплементарности.
Биосинтез белка происходит в рибосомах — с этим мы разобрались.
Где происходит транскрипция? Этот процесс осуществляется в ядре клетки.
Транскрипция происходит в одно и то же время не на всей молекуле ДНК — для этого достаточно одного небольшого участка, отвечающего за определенный ген. Часть двойной спирали ДНК раскручивается, и короткий участок одной из цепей оголяется. Роль матрицы в синтезе молекул иРНК выполняет этот же участок.
Далее в дело вступает фермент РНК-полимераза, который движется вдоль этой цепи. Он соединяет нуклеотиды в цепь иРНК, тем самым удлиняя ее.
Процесс транскрипции осуществляется одновременно на нескольких генах одной хромосомы и на генах разных хромосом.
иРНК, образованная в результате, имеет последовательность нуклеотидов — точную копию последовательности нуклеотидов на матрице.
Если молекула ДНК содержит азотистое основание цитозин, то иРНК — гуанин и наоборот. Комплементарная пара ДНК — аденин-тимин, РНК — аденин-урацил.
тРНК и рРНК (другие типы РНК) синтезируются на специальных генах.
Аминокислоты соединяются с тРНК в цитоплазме. По своей форме молекула тРНК — лист клевера. Вверху этого листа находится антикодон: триплет нуклеотидов, отвечающий за кодировку аминокислоты (ее эта тРНК и переносит).
Количество тРНК определяется количеством аминокислот.
Так как много аминокислот кодируется при помощи нескольких триплетов, то количество тРНК превышает 20. Сегодня известно примерно 60 тРНК.
Ферменты — связующее звено между аминокислотами и тРНК. С помощью молекул тРНК осуществляется транспортировка аминокислот к рибосомам.
Кратко о трансляции в биологии
Что такое трансляция в биологии и как связан с трансляцией биосинтез белка?
В биологии трансляция — это процесс реализации информации о структуре белка, представленной в иРНК последовательностью нуклеотидов, как последовательности аминокислот в синтезируемой молекуле белка.
Как и где происходит биосинтез белка в рамках трансляции и какова схема синтеза белка?
Первый этап трансляции белка — присоединение иРНК к рибосоме. Далее трансляция в биологии — это нанизывание первой рибосомы, синтезирующей белок, на иРНК. Далее трансляция синтеза белка основывается на нанизывании новой рибосомы — по мере того, как предыдущая рибосома продвигается на конец иРНК, который освобождается.
Одна иРНК может одновременно вмещать свыше 80 рибосом, синтезирующих один и тот же белок.
Полирибосома или полисома — группа рибосом, соединенных с одной иРНК,
Информация, записанная на иРНК (а не рибосома), определяет вид синтезируемого белка. Разные белки могут синтезироваться одной и той же рибосомой. Рибосома отделяется от иРНК после того, как синтез белка завершается. Заключительный этап трансляции — это синтез белка или его поступление в эндоплазматическую сеть.
Рибосома включает две субъединицы: малую и большую. Присоединение молекулы иРНК происходит к малой субъединице. Место, в котором рибосома и иРНК контактируют, содержит 6 нуклеотидов (2 триплета). Из цитоплазмы к одному из триплетов постоянно подходят тРНК с различными аминокислотами. Своим антикодоном они касаются кодона иРНК. В случае комплементарности кодона и антикодона, возникает пептидная связь: она образуется между аминокислотой уже синтезированной части белка и аминокислотой, доставляемой тРНК.
Фермент синтетазы участвует в соединении аминокислот в молекулу белка. После отдачи аминокислоты молекула тРНК переходит в цитоплазму, в результате чего рибосома перемещается на один триплет нуклеотидов. Таким образом, происходит последовательный синтез полипептидной цепи. Этот процесс длится до момента достижения рибосомой одного из трех терминирующих кодонов: УАА, УАГ или УГА. Как только это происходит, синтез белка останавливается.
Последовательность того, как аминокислоты включаются в цепь белка, определяется последовательностью кодонов иРНК. В каналы эндоплазматического ретикулюма поступают синтезированные белки. Синтез одной молекулы белка в клетке происходит в течение 1-2 минут.
Схема синтеза белка выглядит следующим образом:
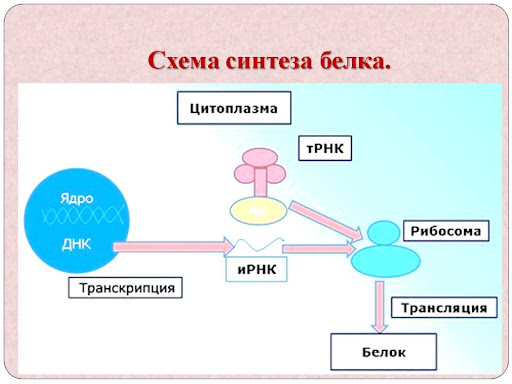
Из схемы биосинтеза белка выше вы можете понять, на чем осуществляется синтез белков, как происходит биосинтез белка, и что кроется за трансляцией и транскрипцией.
Также предлагаем изучить таблицу биосинтеза белка. Здесь описано, как осуществляется синтез белков в клетке, описываются кратко транскрипция и трансляция (этапы синтеза белка).
Таблица биосинтеза белка:
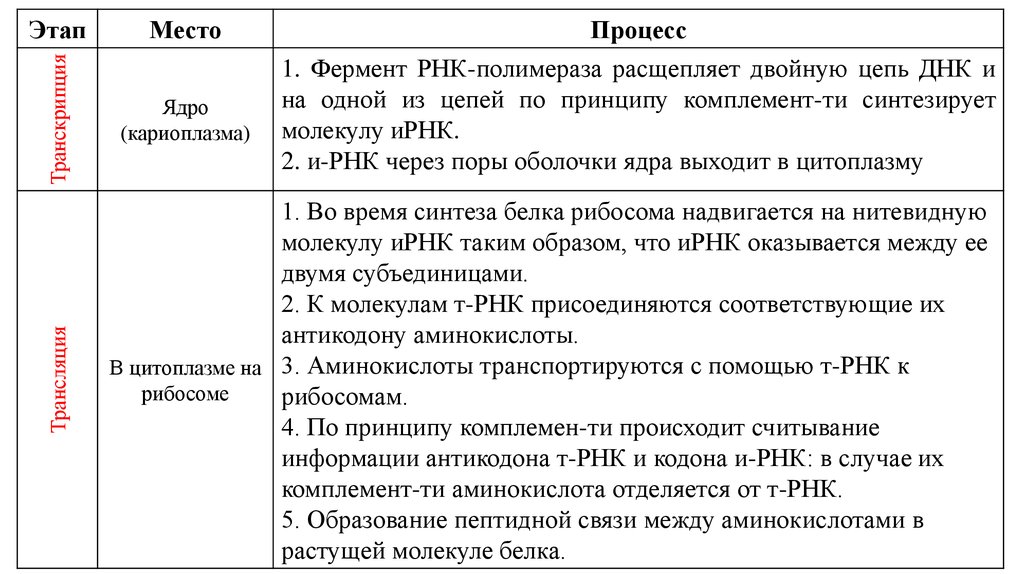
Из таблицы становится ясно, как проходит синтез белка, какие основные этапы синтеза белка, какова роль транскрипции в биосинтезе белка, где происходит синтез белков (место), а также кратко описаны стадии биосинтеза белка.
Таким образом мы охарактеризовали функции различных видов РНК в биосинтезе белков. На примере трансляции и транскрипции мы рассмотрели основные этапы биосинтеза белка.

Основная роль в определении структуры синтезируемого белка принадлежит ДНК.
ДНК – это полимер из нуклеотидов, а белок из аминокислот. Для того, чтобы 4 нуклеотида могли кодировать 20 аминокислот, они должны быть в определенных сочетаниях. Экспериментальным путем было выяснено, что это последовательность из трех нуклеотидов – триплет (или кодон). Разных триплетов из четырех по три будет 64, а аминокислот 20, следовательно, одна и та же аминокислота кодируется несколькими триплетами. И только метионин и триптофан кодируется одним триплетом. Из 64 возможных триплетов 61 кодируют 20 аминокислот, а 3 (нонсенс триплеты) кодируют окончание биосинтеза белка.
Генетический код имеет следующие основные свойства:
Универсальность - код един для всех живых организмов;
Вырожденность (множественность) - одну аминокислоту кодируют от 2 до 6 триплетов;
Триплетность - одну аминокислоту кодируют 3 нуклеотида;
Неперекрываемость - нуклеотид одного триплета не может входить в состав соседнего триплета;
Специфичность - один триплет кодирует строго определенную аминокислоту.
РЕАКЦИИ МАТРИЧНОГО СИНТЕЗА
Реакции транскрипции, трансляции, а также репликации (удвоение ДНК) носят название матричных реакций. Роль матрицы в клетках живых организмов выполняют ДНК или иРНК. Мономерные молекулы (нуклеотиды или аминокислоты), из которых синтезируется полимер, в соответствии с принципом комплементарности располагаются и фиксируются на матрице в строго определенном порядке. После соединения мономерных звеньев в полимерную цепь, новый полимер сходит с матрицы.
Таким образом, матричные реакции:
ДНК материнской клетки репликация ДНК (дочерних клеток – все клетки многоклеточного организма) транскрипция иРНК трансляция белок.
Реакции матричного синтеза обнаружены только в клетках живых организмов и вне организма, в естественных условиях, неизвестны. Поэтому матричный синтез является одним из специфичных свойств живого.
БИОСИНТЕЗ БЕЛКОВ
Структура белка определяется ДНК, которая находятся в ядре. Ген – участок двойной молекулы ДНК – строго определенная последовательность нуклеотидов – содержащая информацию о первичной структуре одной молекулы белка и определяющая развитие какого-либо признака или свойства организма.
В процессе биосинтеза белка выделяют два последовательных этапа транскрипцию и трансляцию.
I этап – транскрипция (протекает в ядре) – процесс переписывания информации с ДНК на молекулу про-иРНК, согласно принципу комплементарности. Предшественница иРНК (про-иРНК) содержит в себе ряд инертных, бессмысленных участков-интронов. В результате процессинга – созревания иРНК, интроны с помощью фермента рестриктазы вырезаются, а оставшиеся экзоны – смысловые участки, несущие информацию о белке, сшиваются ферментом лигазой в цепочку. Процесс сшивания иРНК в одну нить называется сплайсингом. Сплайсинг идет в ядрышках.
II этап – трансляция (протекает в цитоплазме )– синтез полипептидных цепей белков на матрице иРНК, выполняемый рибосомами..
Во время трансляции выделяют такие стадии:
1. Стадия активизации аминокислот – аминокислоты присоединяются к ножке тРНК, образуя комплекс аминоацил-тРНК.
2. Стадия инициации. Синтез белка начинается с образования комплекса иРНК с рибосомой. Сначала поступившая из ядра в цитоплазму иРНК соединяется с малой субъединицей рибосомы. Первый кодон у всех иРНК несет информацию об аминокислоте метионине, т.к. имеет стартовый кодон АУГ, к которому присоединяется антикодон тРНК-метионин, именуемая инициаторной тРНК, т.к. обеспечивает связь малой субъединицы рибосомы с большой.
Дальнейшее удлинение пептидной цепи происходит путем повторения предыдущих фаз.
4. Стадия терминации – окончание биосинтеза белка. иРНК имеет участок, содержащий один из стоп-кодонов при контакте рибосомы с этими кодонами биосинтез белка прекращается, а рибосома распадается на субъединицы.
Каждый вид имеет свой собственный, отличный от других видов, набор белков. Интересно то, что белки , выполняющие идентичные функции у разных видов могут быть похожими или даже абсолютно идентичными.
У белков есть несколько состояний их структур:

Именно первичная структура является определяющей свойства белка . Эта структура – цепь из аминокислот . Аминокислоты, в свою очередь, представляют собой ряд триплетов из нуклеотидов. Решая генетические задачи, обращаются как раз-таки к знакомой таблице:
Каждая аминокислота кодируется тремя нуклеотидами, которые составляют триплет или иначе кодон . Именно последовательность нуклеотидов называется генетической информацией , а участок последовательности, в котором хранится информация о первичной структуре белка это и есть ген .
Нуклеотиды, составляющие ДНК и РНК различаются:
В состав ДНК входят:
В состав РНК входят:
Кроме того, в составе РНК ( рибонуклеиновой кислоты ) сахар рибоза, а ДНК ( дезоксирибонуклеиновой кислоты ) – дезоксирибоза. РНК – одноцепочечная, а ДНК – двухцепочечная.
Свойства генетического кода
- Генетические код триплетен, то есть состоит из аминокислот, которые состоят из триплетов, а триплеты – 3 нуклеотида.
- Генетический код специфичен, один триплет кодирует одну аминокислоту. Посмотрите на таблицу аминокислот. При пересечении всех трех нуклеотидов у нас нет выбора между аминокислотами, таблица указывает лишь на одну определенную аминокислоту.
- Генетический код избыточен, одна аминокислота может быть закодирована более чем одним триплетом нуклеотидов. Здесь важно не запутаться. Опять смотрим на таблицу. Несмотря на то, что пересечение трех нуклеотидов дает 1 аминокислоту, мы видим повторы аминокислот в таблице. Например, аминокислота фенилаланин (сокращенно Фен) кодируется как триплетом УУУ, так и УУЦ. Есть аминокислоты и с большим количеством вариантов.
- Неперекрываемость генетического кода. Один и тот же нуклеотид не может входить в состав разных триплетов. Это не значит, что если у нас есть триплет УУУ, то рядом с ним не может быть триплета УЦГ. Это значит, что урацил в этих триплетах – не одна и та же молекула.
- Генетический код универсален, то есть, несмотря на все различия между живыми организмами, их генетическая информация кодируется одинаковыми аминокислотами, но в разных последовательностях и вариациях.
- Полярность генетического кода. В цепочке аминокислот есть триплеты, которые не несут информацию, а присутствуют для разделения цепи. Так как они не некодирующие, то в таблице у этих сочетаний букв стоит прочерк: УАА, УАГ, УГА.
Транскрипция и трансляция
Из цитологии известно, что генетическая информация у эукариотических клеток заключена в ядре в виде ДНК. Однако процесс биосинтеза белка происходит в цитоплазме на рибосомах.
Спиральная цепь ДНК при раскручивается, в это время по одной из цепочек ДНК строится комплементарная цепь. Из ядра в цитоплазму информация выходит в виде информационной РНК ( иРНК ). иРНК комплементарная одной из цепей ДНК. Этот процесс переписывания называется транскрипцией. Полученная цепь практически идентичная другой цепи ДНК, за исключением того, что вместо тимина там урацил.В процессе участвует специальный фермент РНК-полимераза.

Теперь в ядре есть цепочка, которая уже начала процесс биосинтеза. Как говорилось выше, процесс ассимиляции идет на рибосомах. иРНК выходит в цитоплазму через поры ядерной мембраны
тРНК по форме напоминает лист клевера, а по принципу работы – штамп. На него, прямо как чернила, наслаиваются кодоны.
В цитоплазме начинается процесс трансляции, то есть перевод последовательности нуклеотидов информационной РНК в последовательность аминокислот белка.

По окончанию процесса биосинтеза, цепочка отсоединяется от рибосомы и принимает свою природную структуру: вторичную, третичную или четвертичную.
Задание ollbio10101120162017в1 В геном одного из растений ввели генно-инженерную конструкцию, похожую на использованную в предыдущем задании. Но промотор был заменён на другой – APETALA 3, который включается в лепестках и тычинках. В дальнейшем получили чистую линию трансгенных растений (линия №1). Другие растение трансформировали конструкцией, в которой кодирующая часть гена CRE была поставлена под промотор LEA, активирующийся на поздних стадиях формирования зародыша, а ко ди рующая часть гена Flp – под промотор CAULIFLOWER, который активен в чашелистиках и лепест ках. После этого удалось получить чистую линию №2. А. Какие органы будут светиться у растений из линии №1? Растений из линии №2? Б. Каким будет фенотип растений первого поколения гибридов между линиями №1 и №2? Для об основания ответа опишите структуру генно-инженерной конструкции с флуоресцентными белками. В. Каким будет расщепление по фенотипами и генотипам среди потомков второго поколения, полученных при самоопылении гибридов первого поколения? Считайте, что генно-инженерные конструкции наследуются независимо, а кроссинговер внутри конструкций не происходит
pазбирался: Надежда | обсудить разбор | оценить

Задание ollbio09101120162017в1 Для исследования различных процессов в живых организмах используют флуоресцентные белки. При облучении, например, ультрафиолетовым светом такой белок светится в видимой части спектра. Получены зеленый (GFP, green fluorescent protein), синий (BFP, blue fluorescent protein), желтый (YFP, yellow fluorescent protein) и даже красный (DsRed, из коралла Discosoma striata) флуоресцентные белки. В генно-инженерных конструкциях их ставят под определенные промоторы. В зависимости от этого в живом объекте светятся разные части. 35 CaMV – промотор, который работает во всех клетках растений. Генный инженер создал конcтрукцию, схематическая карта которой приведена ниже. Промотор условно изображён в форме пятиугольника, кодирующие части генов – в форме серых прямоугольников, сайты Lox P и FRT – в виде стрелок, показывающих направление асимметричной части. Для получения белкового продукта необходимо, чтобы кодирующая часть оказалась на той же цепи ДНК, что и промотор, находилась в верной ориентации (и при этом – в сторону 5´- конца нити ДНК относительно промотора). Последовательности Lox P и FRT достаточно короткие и не мешают считыванию и-РНК. Чёрными ромбами обозначены терминаторы транскрипции. Считайте, что в этом месте матричный синтез и-РНК прекращается.А. Каким цветом должны светиться клетки, в которых содержится данная генно-инженерная конструкция? Почему? Б. Нарисуйте в тех же условных обозначениях структуру приведённого участка ДНК после действия рекомбиназы CRE. (Считайте, что при этом рекомбинация произошла только один раз!) Изменится ли после этого свечение клеток? В. Нарисуйте в тех же условных обозначениях структуру приведённого участка ДНК после действия флиппазы Flp. (Считайте, что при этом рекомбинация произошла только один раз!) Изменится ли после этого свечение клеток? Г. Предположим, что на исходную последовательнось ДНК в генно-инженерной конструкции сначала подействовали рекомбиназой CRE, а после этого – флиппазой Flp. Нарисуйте схему строения ДНК для этого случая. Каким будет свечение клеток?
pазбирался: Надежда | обсудить разбор | оценить

Задание ollbio09101120172018в2 У одного из представителей семейства Колокольчиковые (Campanulaceae) – платикодона крупноцветкового (Platycodon grandiflorum) пентамерные цветки, состоящие из круга чашелистиков, круга лепестков, круга тычинок и круга плодолистиков (см. рис.). Иногда среди платикодонов можно найти махровые цветки, у которых на месте тычинок развиваются лепестки. А. Нарисуйте диаграмму махрового цветка платикодона. На диаграмме обозначьте части цветка. Предложите для него формулу. Б. Предположим, что в природной популяции платикодона крупноцветкового возникла форма с махровыми цветками (по остальным признакам форма не отличается от нормы). Образование махровых цветков определяется одной рецессивной мутацией. Ученые пересадили из природы на экспериментальный участок два мутантных и одно нормальное растение. Считая, что при опылении пыльца всех особей смешивается, пыльца из природных популяций не попадает на участок, и при этом возможно самоопыление, рассчитайте, каким может быть расщепление в потомстве первого поколения по генотипам и фенотипам. В. Далее среди потомков первого поколения выбрали только те растения, у которых цветки нормальные, а остальные убрали с участка до опыления. С оставленных растений собрали семена и посеяли. Каким может оказаться расщепление среди потомков второго поколения по генотипу и фенотипу?
А. Опираясь на рисунок, мы видим, что чашелистики изображены свободными, тогда как все лепестки срослись. Пять тычинок свободные, а плодолистиков три, и они также срослись. (У Колокольчиковых завязь нижняя, но это не принципиально для дальнейшего решения.) Можно предложить следующую формулу для типичного цветка в сем. Колокольчиковые: * Ч5 Л(5) Т5 П( — 3) или * Ca5 Co(5) A5 G( — 3) . Поскольку у махровых форм происходит замена тычинок на лепестки, в формуле вместо тычинок нужно указать дополнительный круг лепестков: * Ч5 Л(5)+(5) П( — 3) или * Ca5 Co(5)+(5) G( — 3) . При построении диаграммы должны выполняться следующие принципы: 1. Органы в круге располагаются друг относительно друга под углом 360 : 5 = 72 градуса. 2. В двух соседних кругах органы должны чередоваться, т.е. положение медианы каждого органа должно приходиться строго на промежуток между органами предыдущего круга. Для пентамерного цветка между органами в соседних кругах угол должен составлять 36 градусов. На рисунке видно, что плодолистики (поскольку из три) не могут правильно чередоваться с пятью тычинками. 3. Если рассматривать органы через круг, то их медианы должны находиться друг напротив друга (органы противолежат). 4. Центром цветка считается центр завязи. Поэтому при проверке расположения органов в цветке все линии будут проводиться через центр завязи и центральную (медианную) жилку органа. 5. На рисунке показан цветок с центрально-угловой плацентацией ( гинецей синкарпный). Между гнездами завязи находятся перегородки (септы). Для плодолистика медианой считается линия, делящая угол между септами ровно пополам. Б. Обозначим ген , отвечающий за проявление махровости как А. Поскольку мы знаем, что махровость цветков определяется рецессивной мутацией по этому гену, генотип махровых растений может быть только аа. Взятое из природы нормальное растение могло оказаться как гомозиготой АА, так и носителем рецессивного аллеля Аа. Поэтому возможно два варианта расщепления среди потомков. Из природы были взяты два махровых и одно немахровое растение, и по семенной продуктивности все три растения одинаковы, следовательно, 2/3 семян будет собрано с махровых, и 1/3 – с немахровых растений. Однако пыльцу может образовать только растение с немахровыми цветками. Вариант 1. Немахровое растение – гомозигота АА. Тогда среди потомков в данном скрещивании должно оказаться: 1/3 (≈33.3%) АА 2/3 (≈66.7%) Аа или 1 АА : 2 Аа По фенотипу все потомки окажутся немахровыми. Вариант 2. Немахровое растение – гетерозигота Аа. Среди женских гамет соотношение вклад каждого из растений останется прежним, т.е. 2/3 от всех аллелей а придут от махровых растений. Среди оставшихся 1/3 женских гамет 1/6 будет нести аллель а, и еще 1/6 – аллель А. Таким образом, соотношение среди женских гамет будет 5/6 а и 1/6 А. Среди мужских гамет 1/2 будет нести аллель А, и еще 1/2 – аллель а. Таким образом, среди потомков первого поколения возможно следующее расщепление по генотипам : 1/12 АА (≈8.3%) 6/12=1/2 Аа (50.0%) 5/12 аа (≈41.7%) 1 АА : 6 Аа : 5 аа По фенотипам: 7/12 (≈50.3%) немахровых 5/12 (≈41.7%) махровых 7 немахровых : 5 махровых В. В первом варианте скрещивания махровых растений не окажется. Рассчитаем доли потомков по генотипам и фенотипам во втором поколении. 1/3 (≈33.3%) АА дадут только гаметы А, тогда как 2/3 растений с генотипом Аа дадут половину гамет А и вторую половину гамет а. Таким образом, суммарно гамет А в популяции окажется 2/3, и 1/3 гамет, несущих аллель а. Таким образом, среди потомков второго поколения возможно следующее расщепление по генотипам: 4/9 АА (≈44.4%) 4/9 Аа (44.4%) 1/9 аа (≈11.1%) 4 АА : 4 Аа : 1 аа По фенотипам: 8/9 (≈88.9%) немахровых 1/9 (≈11.1%) махровых 8 немахровых : 1 махровых. Во втором случае (из природы было взято гетерозиготное немахровое растение) после того, как мы удалим все махровые растения, останется 1/7 АА (≈14.3%) и 6/7 Аа (≈85.7%). Последние дадут половину гамет А (3/7) и половину гамет а (3/7). Суммарная доля гамет А составит 4/7. Тогда: Во втором случае расщепление среди потомков второго поколения будет: по генотипам: 16/49 АА (≈32.6%) 24/49 Аа (≈49.0%) 9/49 аа (≈18.4%) 25 АА : 30 Аа : 9 аа По фенотипам: 40/49 (≈81.6%) немахровых 9/49 (≈18.4%) махровых 40 немахровых : 9 махровых.
pазбирался: Надежда | обсудить разбор | оценить
Сначала найдём место расщепления плазмиды рестриктазой BglII: Таких участков оказывается два. В результате расщепления из плазмиды выщепляется короткий фрагмент: Остаётся укороченная линейная ДНК , содержащая интактный ген устойчивости к ампицилину и расщеплённый ген устойчивости к эритромицину. При сшивании липких концов ДНК-лигазой наиболее часто будут соединяться концы этой молекулы и образовываться кольцо длиной 4163 нуклеотида. Такая ДНК будет сообщать клеткам устойчивость к ампицилину и не даст устойчивости к эритромицину. Второй фрагмент из-за небольшой длины не может замкнуться в кольцо. Второй вариант лигирования приводит к сшиванию липких концов двух фрагментов. Он происходит примерно в 10 раз реже, а после сшивки вторая пара липких концов скорее всего также, как и исходный фрагмент замкнётся в кольцо. Таких колец из пары фрагментов может образоваться 4 вида: димеры большого фрагмента в двух разных ориентациях (правый конец с левым концом второго фрагмента и левый конец с правым концом второго фрагмента или правый с правым и левый с левым) и соединения большого и малого фрагмента в двух разных ориентациях (вариант исходной плазмиды и инверсия малого фрагмента). Из них только в варианте исходной плазмиды восстанавливается устойчивость к эритромицину. Линейная молекула, образованная сшиванием двух фрагментов, может присоединить ещё один фрагмент с ещё в 10 раз меньшей частотой. Такие фрагменты в дальнейшем будут циклизоваться в плазмиды трёх размеров: из трёх больших фрагментов, из двух больших и одного малого и одного большого и двух малых. Три малых фрагмента дадут короткую последовательность, которая не сможет замкнуться в кольцо и существовать в клетке. В каждом размерном классе будет несколько вариантов с разной ориентацией фрагментов. Только в одном из них восстановится ген устойчивости к эритромицину: правый конец большого фрагмента соединяется с левым концом малого фрагмента, а правый конец малого фрагмента – с левым концом второго большого фрагмента, а оставшиеся концы двух больших фрагментов соединяются с образованием кольцевой плазмиды длиной 8363 пары нуклеотидов. Доля таких молекул будет менее 1% всех плазмид . Вероятность образования плазмид из 4 и более фрагментов ещё на порядок ниже и их обнаружение при данном числе полученных трансформированных клеток нереально. А. Так как расщепление рестриктазой не затрагивает ген устойчивости к ампицилину, все клетки, в результате трансформации получившие любую плазмиду, будут устойчивы к ампицилину и вырастут на среде с этим антибиотиком. Таким образом из 33506 выросших колоний плазмиду получили 578, выросших на ампицилине. Эффективность трансформации представляет долю трансформированных клеток от общего их числа, т.е. 573 : 51366 × 100% = 1.12% Б. На эритромицине могут вырасти только те клетки, в которые попали плазмиды, в которых в результате лигирования восстановится последовательность нуклеотидов в гене устойчивости к этому антибиотику, расщеплённому рестриктазой. Остальные плазмиды, полученные по приведённой методике, будут содержать либо ген с выщепленным коротким фрагментом, что приведёт либо к утрате стартового кодона (если обозначенный зелёным цветом кодон является стартовым), либо к сдвигу рамки считывания (т.к. число удалённых нуклеотидов не кратно трём), либо, при инверсии короткого фрагмента, к появлению стоп-кодонов т.е. прекращению синтеза белка . Таким образом большинство полученных плазмид не обеспечат устойчивости к эритромицину. В. Рост на эритромицине могут обеспечить только плазмиды, несущие восстановленную последовательность гена устойчивости. Такие плазмиды могли образоваться из одного большого и одного малого фрагмента (4200 пар, исходная плазмида )) или из двух больших и одного малого (8363 пары, начало и конец гена из разных копий большого фрагмента). Г. Получается 1 размер из одного большого фрагмента, два размерных класса из двух фрагментов и три размерных класса из трёх фрагментов, то есть 6 размерных классов. (В реальности различить по длине плазмиды, отличающиеся на длину малого фрагмента, т.е. менее чем на 0,5%, невозможно. Поэтому в эксперименте, например на электрофореграмме, будут видны лишь три размерных класса, соответствующие 1, 2 или 3 копиям большого фрагмента.)
pазбирался: Надежда | обсудить разбор | оценить
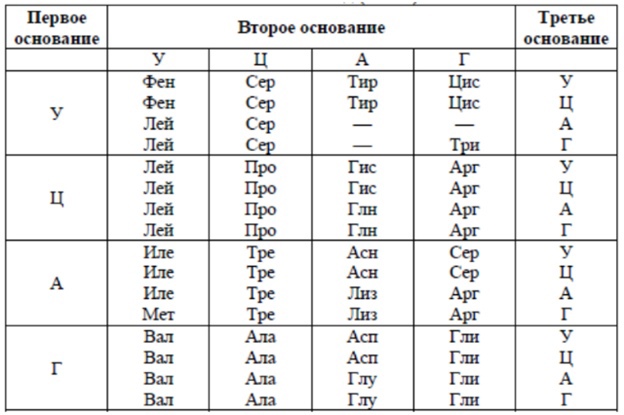
Задание EB2719t Фрагмент цепи ДНК имеет следующую последовательность нуклеотидов: АГТЦЦГАТГТГТ. Определите последовательность кодонов на иРНК, антикодоны соответствующих тРНК и аминокислотную последовательность соответствующего фрагмента молекулы белка. Ответ поясните. Для решения задания используйте таблицу генетического кода.
По принципу комплементарности строим иРНК на матрице ДНК.
Теперь, опять же по принципу комплементарности, строим тРНК.
АГУ, ЦЦГ, АУГ, УГУ
Определяем с помощью таблицы аминокислотную последовательность синтезируемого белка по иРНК.
Читайте также: