
Теория синзооспоры захваткина кратко
Обновлено: 05.07.2024
Относительно происхождения многоклеточных животных существует несколько гипотез. Гипотезы подразделяются на две группы: колониальные и полиэнергидные.
Колониальные гипотезы базируются на признании предками многоклеточных колониальных простейших. Ими являются:
5. А.В.Иванов (1967г.) за основу принимает гипотезу фагоцителлы. Предком многоклеточных является колония воротничковых жгутиконосцев с голозойным способом питания. Моделью фагоцителлы является трихоплакс. Фагоцителла дала начало двум типам: губкам и пластинчатым животным.
Согласно этой теории предком многоклеточных была гастрея - многоклеточный двуслойный организм. Она произошла от колониальных протистов с шарообразными колониями. Процесс интеграции клеток в колонии сделал возможным разделение функций между клетками: передние клетки утрачивают жгутики и превращаются в фагоциты, сидящие во впячивании на переднем конце - образуется кишечник.Остальные клетки утрачивают пищеварительную функцию и становятся чисто двигательными. Рот гастреи находился на переднем конце, и пища "сама заплывала" в кишечник. Симметрия у гастреи была радиальной. При переходе к сидячему образу жизни ее потомки эволюционировали в губок и кишечнополостных, а при переходе к ползанию по дну - в плоских червей и всех остальных многоклеточных.
Эта теория во многом сходна с предыдущей. Но предком многоклеточных считается фагоцителла. Фагоцителла не имела рта и кишечника, пищеварение было внутриклеточное. Рот сформировался,как просвет между клетками наружного слоя, ведущий во внутреннюю паренхиму. Располагался он, в отличие от гастреи на заднем конце тела. Кишечника еще не было. Но теперь возникла возможность питаться более крупной добычей: внутренние клетки могли окружать ее, образуя гигантскую пищеварительную вакуоль. Однако для хищничества нужна еще способность ловить добычу. Поэтому хищничать научились только настоящие многоклеточные - после того, как у них возникли мышцы и управляющая ими нервная система. Постепенно у потомков фагоцителлы сформировался постоянный кишечник. По мере увеличения размеров он мог усложняться: возникли боковые карманы, чтобы доставлять пищу к наружным слоям клеток. В дальнейшем у некоторых животных эти карманы могли отделиться, дав начало полости тела - целому. Фагоцителла обитала в толще воды. Нетрудно представить себе, как от нее могли произойти современные группы животных при переходе к жизни на дне. Когда рта еще не было, осевшая на дно фагоцителла "превратилась" в трихоплакса. После появления рта, но до появления кишечника при переходе к ползанию возникли бескишечные турбеллярии. Рот у них сместился на брюхо, и они стали двустороннесимметричными. После появления кишечника часть потомков фагоцителлы перешли к сидячему образу жизни на дне - они превратились в кишечнополостных.
Гаметы и зигота - единственные одноклеточные стадии в жизненном цикле животных. Многоклеточных поколений может быть в жизненном цикле несколько. Согласно данной теории многоклеточные произошли от колониальных протистов. У протистов встречаются клетки, сильно увеличенные за счет запасания питательных веществ - как яйцеклетка у животных. Часто такие клетки делятся несколько раз подряд - это похоже на дробление. Таким способом образуются у протистов одноклеточные мелкие расселительные стадии - зооспоры. У колониальных протистов зооспоры могут оставаться все вместе, образуя колонию - синзооспору. В процессе эволюции могла произойти неотения и утратиться взрослая сидячая стадия. Таким образом бластула - это синзооспора, семья зооспор.
Отличия от теорий фагоцителлы и гастреи:
Считается, что никогда не существовало однослойного шарообразного предка. Об этом свидетельствует то, что у всех многоклеточных бластулы не питаются. Не питаются и образующиеся из них паренхимулы. Поэтому и у древних многоклеточных такие стадии не были взрослыми организмами - это были всегда только личинки.
Сидячий образ жизни примитивных взрослых многоклеточных.
Наиболее примитивной из расселительных личинок считается бластула. В паренхимулу она превращается, готовясь к превращению во взрослый организм. Этот метаморфоз происходит после перехода к сидячему образу жизни. У всех остальных многоклеточных взрослая сидячая стадия утратилась. У этих животных личинки стали взрослыми - произошла неотения.
Эта теория на сегодняшнем уровне знаний имеет лишь исторический интерес. Она предполагает, что предками многоклеточных были сложно организованные протисты, такие как инфузории, а органы многоклеточных образовались в результате отделения мембранами органелл. Так, в соответствии с этой теорией кишечник образовался из глотки инфузории-туфельки, выделительная система - из ее сократительных вакуолей, покровы - из периферического слоя цитоплазмы. Эта теория полностью не соответствует взглядам современной науки и является безусловно ошибочной.
О предках многоклеточных
В рамках рассмотреннных теорий предполагается, что скорее всего предками многоклеточных была группа воротничковых жгутиконосцев - хоанофлагеллят. В пользу этого говорит то, что для примитивных многоклеточных очень характерны воротничково - жгутиковые клетки, строение которых очень слабо отличается от строения хоанофлагеллят. Подходящий у воротничковых жгутиконосцев и способ питания. Все они гетеротрофы, питающиеся за счет фагоцитоза и пиноцитоза. Им свойственна колониальность. Среди колоний есть даже такие, у которых наружные клетки имеют воротнички и жгутики, а внутренние - амебоидные. Единственный недостаток хоанофлагеллят, как предков многоклеточных, состоит в том, что до сих пор у них достоверно известен только один способ размножения - деление пополам. Ничего похожего на половое размножение или на дробление у них нет. Тем не менее, многие ученые считают эту группу протистов наиболее вероятными предками всех Metazoa
Многоклеточные организмы - это организмы , состоящие [внезапно] из более чем одной клетки , в отличие от одноклеточных организмов.
Все виды животных, наземные растения и большинство грибов являются многоклеточными, как и многие водоросли , тогда как некоторые организмы являются частично одно- и частично многоклеточными, как слизистые плесени и социальные амебы, такие как род Dictyostelium .
Рис. 1. Колония Dictyostelium в процессе агрегации (Автор Bruno in Columbus)
Многоклеточные организмы возникают различными путями, например, путем деления клеток или путем агрегации (соединения) множества отдельных клеток. Колониальные организмы являются результатом того, что отдельные индивидуальные клетки объединяются в колонию. Однако часто бывает трудно отделить колониальных простейших от настоящих многоклеточных организмов, потому что эти две концепции не различаются.
Возникновение многоклеточности
Многоклеточность развивалась независимо по меньшей мере в 46 раз у эукариот [статья на эту тему на английском языке тут ], а также у некоторых прокариот, таких как цианобактерии, миксобактерии, актиномицеты, у экстремофильной бактерии Magnetoglobus multicellularis и у группы метановых архебактерий Methanosarcina.
Однако сложные многоклеточные организмы развивались только в шести эукариотических группах: животные, грибы, бурые водоросли, красные водоросли, зеленые водоросли и наземные растения. Мноклеточность неоднократно развивалась для Chloroplastida (зеленые водоросли и наземные растения), один или два раза для животных, один раз для бурых водорослей, три раза в грибах (хитриды, аскомицеты и базидиомицеты) и, возможно, несколько раз для слизистых плесени и красные водоросли.
Первым доказательством многоклеточности являются цианобактериоподобные организмы, которые жили 3–3,5 миллиарда лет назад.
Потеря многоклеточности
Некоторые группы организмов в своем эволюционном развитии утратили многоклеточность.
Многие грибы (например, Saccharomycotina, Cryptococcus и другие дрожжи), некоторых красные водоросли (например, Porphyridium sp. ), но возможно, что они примитивно одноклеточные.
Потеря многоклеточности также считается вероятной у некоторых зеленых водорослей (например, Chlorella vulgaris и некоторые из группы Ulvophyceae). В других группах, как правило, паразитах, происходило уменьшение количества клеток или уменьшение типов клеток (например, многие представители Myxozoa, многоклеточные организмы, ранее считавшиеся одноклеточными, являются паразитами морских животных).
Гипотезы происхождения
Одна из гипотез о происхождении многоклеточности заключается в том, что группа функционально-специфических клеток объединяется в слизистую массу, которая перемещается как многоклеточная единица. Это в основном то, что делают слизевики.
Другая гипотеза состоит в том, что примитивная клетка подвергалась делению ядра, превращаясь в коеноцит (клетку с множеством ядер). Мембрана, в таком случае, сформировалась бы вокруг каждого ядра (и клеточного пространства и органелл, занятых в пространстве), таким образом приводя к группе связанных клеток в одном организме. Этот механизм наблюдается у дрозофилы.
Третья гипотеза состоит в том, что при разделении одноклеточного организма дочерние клетки не могли отделиться, что привело к конгломерации идентичных клеток в одном организме, что впоследствии могло развить специализированные ткани. Это то, что делают эмбрионы растений и животных, а также колониальные хоанофлагелляты.
Гиф. 1. Tetrabaena socialis - зеленая водоросль, самый примитивный многоклеточный организм, состоящий из четырех клеток, несущих по два жгутика
Поскольку первые многоклеточные организмы были простыми, мягкими организмами, у которых не было костей, раковин или других твердых частей тела, они плохо сохранились в ископаемом материале. Одним из исключений может быть Обыкновенная губка (Demospongiae), которая, возможно, оставила химический след в древних породах.
Самые ранние окаменелости многоклеточных организмов включают Grypania spiralis (см. рисунок 1) и окаменелости черных сланцев палеопротерозойской фервильской группы в Габоне (2,5 млрд лет назад).
Рис. 2. Grypania spiralis в отложениях
Далее я более подробно расскажу о современных теориях возникновения многоклеточности:
Теория симбиоза
Эта теория предполагает, что первые многоклеточные организмы произошли от симбиоза (кооперации) различных видов одноклеточных организмов, каждый из которых играет разные роли. Со временем эти организмы станут настолько зависимыми друг от друга, что не смогут выжить независимо, что в конечном итоге приведет к объединению их геномов в один многоклеточный организм. Каждый соответствующий организм стал бы отдельной линией дифференцированных клеток во вновь создаваемых видах.
Однако проблема этой теории заключается в том, что до сих пор неизвестно, как ДНК каждого организма может быть включена в один геном, чтобы составить их как единый вид. Хотя считается, что такой симбиоз имел место (например, митохондрии и хлоропласты в клетках животных и растений - эндосимбиоз), он случался очень редко, и даже тогда геномы эндосимбионтов сохраняли элемент различия, раздельно реплицируя свою ДНК во время митоза видов-хозяев.
Например, два или три симбиотических организма, образующих составной лишайник , хотя и зависят друг от друга в отношении выживания, должны воспроизводиться по отдельности, а затем переформироваться, чтобы создать еще один отдельный организм.
Теория синцития
Эта теория утверждает, что один одноклеточный организм с несколькими ядрами мог бы создать внутренние мембранные перегородки вокруг каждого из своих ядер.
Многие протисты, такие как инфузории или слизевики, могут иметь несколько ядер, что подтверждает эту гипотезу. Однако простого наличия нескольких ядер недостаточно для поддержки теории. Множественные ядра инфузорий различны и имеют четко дифференцированные функции. Макроядро служит потребностям организма, а микроядро используется для полового размножения с обменом генетического материала.
Синцитиальные слизевики образуются из отдельных амебоидных клеток, подобно синцитиальным тканям некоторых многоклеточных организмов, а не наоборот.
Чтобы считаться действительной, эта теория нуждается в наглядном примере и механизме образования многоклеточного организма из ранее существовавшего синцития.
Рис. 3. Слизевик Fuligo septica (Автор фото Kreis Tuttlingen)
Колониальная теория
Теория предполагает, что симбиоз многих организмов одного и того же вида (в отличие от симбиотической теории , которая предполагает симбиоз разных видов) привел к многоклеточному организму.
Преимущество колониальной теории состоит в том, что она была обнаружена независимо в 16 различных типах простейших. Например, во время нехватки пищи амеба Dictyostelium (Рис. 1) группируется в колонии, которая перемещается как единое целое в новое место. Некоторые из этих амеб затем немного отличаются друг от друга.
Другими примерами колониальной организации у простейших является, например, Volvox sp. (Рис. 4), который состоит из 500-50000 клеток (в зависимости от вида), только часть из которых размножается.
Рис. 4. Колонии вольвокса (Автор фото Frank Fox)
В отечественной литературе колониальную теорию обычно делят на теорию гастреи (Геккель, 1872), теорию фагоцителлы (Мечников, 1878) и теорию синзооспоры (Захваткин, 1949). Обе теории довольно схожи, разница в том, как, согласно эти теориям, происходила интеграция клеток в колонии.
Теория гастреи
Согласно этой теории предком многоклеточных была гастрея - многоклеточный двуслойный организм. Она произошла от колониальных протистов с шарообразными колониями. Процесс интеграции клеток в колонии сделал возможным разделение функций между клетками: передние клетки утрачивают жгутики и превращаются в фагоциты, сидящие во впячивании на переднем конце - образуется кишечник.Остальные клетки утрачивают пищеварительную функцию и становятся чисто двигательными. Рот гастреи находился на переднем конце, и пища "сама заплывала" в кишечник. Симметрия у гастреи была радиальной. При переходе к сидячему образу жизни ее потомки эволюционировали в губок и кишечнополостных, а при переходе к ползанию по дну - в плоских червей и всех остальных многоклеточных (Рис. 5).
Так развиваются ланцетники и коралловые полипы.
Теория фагоцителлы
Фагоцителла не имела рта и кишечника, пищеварение было внутриклеточное. Рот сформировался,как просвет между клетками наружного слоя, ведущий во внутреннюю паренхиму. Располагался он, в отличие от гастреи на заднем конце тела. Кишечника еще не было. Но теперь возникла возможность питаться более крупной добычей: внутренние клетки могли окружать ее, образуя гигантскую пищеварительную вакуоль. Однако для хищничества нужна еще способность ловить добычу. Поэтому хищничать научились только настоящие многоклеточные - после того, как у них возникли мышцы и управляющая ими нервная система. Постепенно у потомков фагоцителлы сформировался постоянный кишечник. По мере увеличения размеров он мог усложняться: возникли боковые карманы, чтобы доставлять пищу к наружным слоям клеток. В дальнейшем у некоторых животных эти карманы могли отделиться, дав начало полости тела - целому. Фагоцителла обитала в толще воды. Нетрудно представить себе, как от нее могли произойти современные группы животных при переходе к жизни на дне. Когда рта еще не было, осевшая на дно фагоцителла "превратилась" в трихоплакса . После появления рта, но до появления кишечника при переходе к ползанию возникли бескишечные турбеллярии. Рот у них сместился на брюхо, и они стали двустороннесимметричными. После появления кишечника часть потомков фагоцителлы перешли к сидячему образу жизни на дне - они превратились в кишечнополостных (Рис. 5).
Так развиваются обыкновенные губки, гидроидные и сцифоидные стрекающие.
Основным кандидатом на роль предка всех многоклеточных является воротничковый жгутиконосец (Choanoflagellata), их клетки снабжены жгутиком, окруженный воротничком. Жгутик создает токи жидкости, позволяющие хоанофлагеллятам плавать в толще воды. Эти же токи пригоняют в воротничковую зону бактерий, которыми хоанофлагелляты питаются. Многие их этих простейших образуют колонии, причем легко переходят из одноклеточного состояния в колониальное и обратно.
Рис. 5. Гастрея и фагоцителла
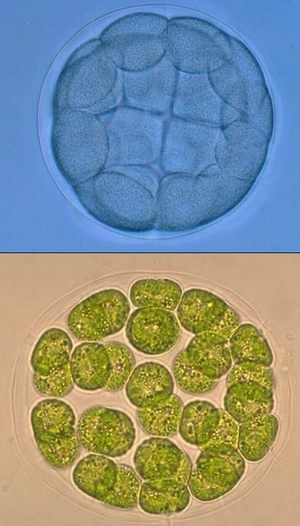
Теория синзооспоры
Согласно данной теории многоклеточные также произошли от колониальных протистов. У протистов встречаются клетки, сильно увеличенные за счет запасания питательных веществ - как яйцеклетка у животных. Часто такие клетки делятся несколько раз подряд - это похоже на дробление. Таким способом образуются одноклеточные мелкие расселительные стадии - зооспоры.
У колониальных протистов зооспоры могут оставаться все вместе, образуя колонию - синзооспору. В процессе эволюции могла произойти неотения и утратиться взрослая сидячая стадия.
Отличия от теорий фагоцителлы и гастреи:
- Считается, что никогда не существовало однослойного шарообразного предка. Об этом свидетельствует то, что у всех многоклеточных бластулы не питаются. Не питаются и образующиеся из них паренхимулы. Поэтому и у древних многоклеточных такие стадии не были взрослыми организмами - это были всегда только личинки.
- Сидячий образ жизни примитивных взрослых многоклеточных.
- Наиболее примитивной из расселительных личинок считается бластула. В паренхимулу она превращается, готовясь к превращению во взрослый организм. Этот метаморфоз происходит после перехода к сидячему образу жизни. У всех остальных многоклеточных взрослая сидячая стадия утратилась. У этих животных личинки стали взрослыми - произошла неотения.
Преимущества многоклеточности
Многоклеточность позволяет организму превышать пределы размера, обычно налагаемые диффузией: отдельные клетки с увеличенным размером имеют уменьшенное отношение поверхности к объему и испытывают трудности с поглощением достаточного количества питательных веществ и их транспортировкой по всей клетке.
Таким образом, многоклеточные организмы имеют конкурентные преимущества увеличения размера без его ограничений. Многоклеточность также позволяет увеличивать сложность, позволяя дифференцировать типы клеток в пределах одного организма.
Я попытался привести современные данные по возникновению многоклеточности. Тема спорная, вариантов куча и все они интересны. Есть, что добавить - пиши в комменты.
Свыше полувека назад выдающийся зоолог А. А. Захваткин разработал гипотезу происхождения многоклеточных животных, основанную на сопоставлении жизненных циклов одноклеточных организмов с индивидуальным развитием многоклеточных. Эти идеи продолжает творчески развивать сын А. А. Захваткина, профессор Сельскохозяйственной академии им. Тимирязева Ю. А. Захваткин.
В связи с тем, что данная статья, с одной стороны, представляет большой теоретический интерес, а с другой – довольно сложна для понимания, редакция сайта Журнала Общей Биологии обратилась к автору, Юрию Алексеевичу Захваткину, с просьбой написать для нашего сайта краткое популярное изложение основных идей статьи. Юрий Алексеевич откликнулся на эту просьбу, за что мы ему очень благодарны. Ниже публикуются два текста:
1) Авторский синопсис, написанный Ю.А.Захваткиным;
2) Обычный редакционный синопсис, написанный редактором сайта А. В. Марковым. Возможно, этот второй текст является излишним, но мы пока решили его на всякий случай оставить (комментарии читателей приветствуются).
Преемственность поколений и их интеграция
С позиций "палинтомической гипотезы" и "теории эмбрионизации развития" А.А. Захваткина (1946, 1949, 1953) онтогенез многоклеточных существ оформился за счет интеграции жизненного цикла Простейших. При этом одноклеточная материнская особь уже не делится сразу же на две дочерние особи а, продолжая избыточный, не прерываемый делениями рост (гипертрофическую дистомию), увеличивает свои размеры в сотни и тысячи раз. Затем она приступает к серии чередующихся, сближенных во времени (палинтомических) делений, при которых формирующиеся клетки, не успевая вырасти, дифференцироваться и обрести самостоятельность, приступают к делениям в свою очередь. Устанавливаются новые, характерные для Многоклеточных, средства и способы преемственности поколений - гипертрофическая дистомия преобразуется в оогенез, а повторные палинтомические деления - в дробление сформированного яйца. В результате ряд последовательных клеточных поколений (делящихся бластомеров) переходят на иждивение гипертрофированной яйцеклетки, а формирующий ее организм через положенный срок погибает. "Потенциальному бессмертию" одноклеточных Простейших противопоставляется "потенциальное бессмертие" гамет и временное, преходящее существование многоклеточной особи материнского поколения. Чередование последовательных поколений устанавливается на уровне многоклеточных особей, а не их клеток; средством преемственности поколений становится яйцо - своеобразная морфогенетическая среда развития потомка, а не тот половинный набор органелл, который переходит к дочерним особям при делении одноклеточной материнской особи Простейших.
Проведя исследование и анализ преобразований онтогенезов в восходящем морфологическом ряду Первичноротых животных, мы выявили общие тенденции в развитии преемственности поколений. Реализация этих тенденций обеспечивается и совершенствованием оогенеза, и эмбрионизацией развития за счет смещения на самые ранние стадии более поздних онтогенетических событий. Для регистрации преобразований и корректного сравнения онтогенезов потребовалась разработка проморфологических критериев - своего рода координат в ориентации процессов формообразования. Мы воспользовались соотношениями морфологических осей яйца, личинки и формирующегося организма последовательно проявляющихся по ходу развития.
В мелких и бедных желтком яйцах самых примитивных кишечнополостных и губок развиваются весьма простые бластулообразные личинки. Нередко ось их тела устанавливается независимо от исходной (протозойной) полярности яйца. Эти исходно неустановившиеся соотношения морфологических осей яйца и личинки стабилизируются у более совершенных кишечнополостных и губок. При общей ориентации процессов формообразования (протаксонии или первичноосности) исходная анимально-вегетативная полярность яйца становится средством дифференциации бластомеров, определяя равнокачественность меридиональных и разнокачественность экваториального делений бластомеров. Иными словами, она определяет пространственную дифференциацию (регионализацию) делящихся клеток. В отличие от простейших, дифференцирующихся в последовательных поколениях после отделения дочерних клеток-особей, здесь происходит синхронная дифференциация в пространстве однородных клеток одного (обычно 5 - 7) поколения. Такого рода отношения достигают предельного выражения и совершенства в спиральном дроблении кольчатых червей, моллюсков и других животных. Оно становится средством сегрегации ооплазмы - каждый отделяющийся бластомер представляет зачаток органа формирующейся личинки - трохофоры. Это наглядно проявляется в экспериментальном подавлении или изменении форм дробления, в эмбрионизации развития при обогащении яиц желтком и при формировании гетероквадрантного дробления.
Обогащение желтком бластомеров основного квартета, гипертрофия D - квадранта, эмбрионизация и депрессия трохофоры сопровождаются развитием плагиаксонии (прекрестноосности) - морфологические оси яйца и формирующегося в нем червя перекрещиваются друг с другом. Плагиаксония характерна для олигохет и пиявок, и является исходным состоянием для членистоногих разных групп. Однако членистоногие противопоставляются кольчатым червям разнообразием форм дробления яиц, сменяющим морфогенетическую точность спирального дробления. Это разнообразие становится возможным при изменении роли дробления в эмбриогенезе - оно перестает быть средством сегрегации ооплазмы. Действительно, изменение форм дробления яиц членистоногих в эксперименте обычно остается без морфогенетических последствий.
Переходя к расшифровке онтогенезов насекомых, отметим глубокую эмбрионизацию их развития, предопределенную совершенствованием процессов оогенеза и организацией яичников в овариолы. В этих яйцевых трубочках формирующийся ооцит заполняется желтком и приобретает необычную для других животных продолговатую, нередко, билатерально-симметричную форму. По размерам в отношении тела самки яйца большинства насекомых сопоставимы с новорожденными детенышами Плацентарных. Очень длинные яйца пчелы Xylocopa достигают 15 мм.
Особенно богаты желтком крупные яйца тараканов, прямокрылых, палочников и других насекомых с неполным превращением (Hemimetabola). Они формируются в овариолах паноистического типа без участия абортивных яйцеклеток - трофоцитов. Лишь у уховерток, как и у многих насекомых с полным превращением (Holometbola), овариолы мероистического типа. Здесь не все оогонии имеют равные перспективы. Испытав премейотические события, они синхронно делятся, образуя компактные цисты, из 2, 4, 8 и большего (до 128) числа клеток. Однако лишь одной из них суждено стать яйцом; все остальные преобразуются в трофоциты, сохраняющие плазматические связи (фусомы) с ооцитом и друг с другом. Перекачивая по фусомам все необходимое для яйца, включая не только собственную ооплазму, но и органеллы (митохондрии, рибосомы и т.п.), трофоциты обеспечивают ускорение и вителлогенеза (образования желтка), и числа партий откладываемых яиц. По обилию желточных гранул яйца этих насекомых уступают Hemimetabola, но превосходят их по содержанию активной плазмы.
В проморфологическом отношении эмбриогенезы даже самых примитивных насекомых своеобразны. Анимально-вегетативная полярность их яиц не проявляется вовсе, а исходная для членистоногих плагиаксония предполагает направленность органогенезов и обрастания желтка наименее экономным способом - по длинной оси продолговатого яйца. Именно поэтому оформляющаяся зародышевая полоска "преждевременно" смещается на вентральную сторону яйца и впоследствии обрастает желточную массу кратчайшим способом. Смещения зародыша (ориентационные бластокинезы) подчас приобретают самые причудливые траектории – исходно плагиаксонное положение зародыша дополняется еще одним перекрестом морфологических осей, так что возникает состояние ортоплагиаксонии (прямопрекрестноосности).
Между тем большинству насекомых свойственно изначальное смещение зоны обособления зародыша в задне-вентральном направлении и это состояние мы признаем типичным. Обращаясь к наиболее совершенным насекомым - высшим Двукрылым и Перепончатокрылым мы отмечаем изначальное положение зародыша на вентральной стороне яйца. При этом отпадает необходимость в ориентационных бластокинезах - ось тела оформляющегося зародыша с самого начала совпадает с продольной осью яйца. Возникает состояние ортаксонии (прямоосности) и все формообразовательные процессы, как и у примитивных кольчатых червей, приобретают общее направление. Правда, здесь они ориентируются на построение билатерально-симметричного организма, а не бластулообразной трохофоры с преобладающей радиальной симметрией. Примечательно, что у высших Двукрылых и Перепончатокрылых, так же как и у примитивных кольчатых червей, то-есть при ортаксонии и протаксонии, в сформированном, но еще не оплодотворенном яйце уже заложены все основные элементы плана строения развивающегося существа. Об этом свидетельствуют результаты экспериментов, в которых выключение дробления не препятствует проявлению процессов, имитирующих образование зародышевой полоски насекомых или "одноклеточной трохофоры" у кольчатых червей.
Повреждения ограниченных зон яйца дрозофилы, ответственных за образование определенных органов и структур в соответствии с "картами расположения зачатков", позволяет получить вполне развитых личинок, однако лишенных этих органов и структур. Более того, еще более ограниченные повреждения, например, в зонах слюнных желез, позволяют получить нормальных личинок, из которых развиваются лишенные желез взрослые насекомые. Учитывая, что оформление этих презумптивных зон происходит в ходе оогенеза, под контролем лишь одного материнского генотипа, мы должны признать глубочайшую интеграцию поколений, возрастающую в ряду форм Первичноротых животных. Это суждение получает вполне однозначное подтверждение в открытии гомеозисных и каскада регуляторных генов, начиная от материнских, а также в локализации конкретных морфогенов, таких как, например, Bicoid.
Итак, связь материнского и дочернего поколений становится все более глубокой и тесной Особое значение в становлении этой связи у насекомых имела организация яичников в овариолы и резкая интенсификация процессов образования желтка и ооплазмы (вителлогенеза). При этом ядерно-плазменное отношение испытывает колоссальное напряжение, а громадные запасы желтка вынуждают развивающийся эмбрион к ориентационным бластокинезам. Возникает ортоплагиаксония и, по мере редукции бластокинезов, ортаксония, допускающая преформацию всех основных зачатков личинки и даже имаго еще до начала развития потомка.

Гипертрофированная материнская особь делится на 2, затем на 4, 8, 16 и т.д. потомков, которые, не успев вырасти и дифференцироваться, продолжают в свою очередь делиться. Такой способ деления называется палинтомическим , и он очень напоминает дробление яйца многоклеточных. Так образуется множество не питающихся самостоятельно расселительных зооспор. Из них вырастают одноклеточные вегетативные особи - трофозоиты, которые уже не только делятся, но и питаются и растут (деления, разделенные фазами роста, называются монотомическими ).
Бесполое размножение путем палинтомии возможно благодаря тому, что целый ряд палинтомических поколений обеспечивается запасами, накопленными материнским организмом.
От такого жизненного цикла – один шаг до перехода к многоклеточности. Суть преобразования состояла в том, что палинтомические поколения клеток окончательно утратили свою самостоятельность одноклеточных индивидов, и, оставшись на иждивении материнского организма, превратились в бластомеры – эмбриональные клетки.


Потомок древнего боярского рода Алексей Захваткин создал остроумную эволюционную теорию
Короткая жизнь биолога Алексея Захваткина увенчалась открытием мирового значения.

Энтомолог Захар Родионов (слева) и Алексей Захваткин (справа)
Фото: Аналитическая служба МГУ имени М.В.Ломоносова
Энтомолог Захар Родионов (слева) и Алексей Захваткин (справа)
Фото: Аналитическая служба МГУ имени М.В.Ломоносова
Алексей Языков родился в 1905 году в Екатеринбурге, детство же провел в Швейцарии, где получил прекрасное домашнее образование, в частности, в совершенстве знал французский и английский языки. С началом Первой мировой войны семья Языковых спешно покинула город Монтрё и вернулась в Россию, чтобы обосноваться в Москве. Из-за происхождения — Алексей был древнего боярского рода Языковых — он после октябрьского переворота не мог поступить в высшее учебное заведение.
Пригодилось домашнее образование: Алексей зарабатывал на жизнь, делая зарисовки растений и мелких животных для профессоров Дмитрия Сырейщикова и Григория Кожевникова. C их согласия он начал посещать университетские лекции по биологии как вольнослушатель, но основного своего занятия не оставил и продолжал совершенствовать свое мастерство художника, посещая курсы живописи профессора Дмитрия Кардовского.
В 1926 году Алексею Языкову, хорошо известному в кругу московских биологов художнику-графику, предложили место лаборанта-художника в Среднеазиатском институте защиты растений в Ташкенте. Ученые этого института постоянно устраивали экспедиции, Алексей, как и в Москве, делал рисунки органических форм, но еще проявил замечательные способности к наблюдениям и исследованиям. Несмотря на отсутствие образования, через год Алексей Языков становится ассистентом, затем штатным специалистом — энтомологом, а с 1931 года — старшим научным сотрудником базового Всесоюзного института защиты растений в Ленинграде.
Однажды экспедиционный отряд, в котором был и Алексей Языков, был захвачен басмачами, боровшимися с большевистской властью. Все оборудование, все материалы и документы басмачи отобрали, а ученых отпустили. Когда стал вопрос о восстановлении утраченных документов, Алексей Языков при поддержке профессора Василия Плотникова и других коллег, оформил справку об утрате диплома о высшем образовании; кроме того, в новом паспорте он стал не Языковым, а Захваткиным, такова была фамилия его отчима.
В 1932 году в немецком энтомологическом журнале выходит первая статья Алексея Захваткина.
За амбарных клещей Алексей Захваткин и взялся.
Он разработал методы морфологического анализа, по которым и сейчас определяют вид клеща; практически с нуля создал систематику клещей. До него не было точных рисунков даже некоторых широко распространенных видов клещей. Алексей Захваткин первым разделил клещей на три отряда. Эти исследования, продолжавшиеся и в годы Великой Отечественной войны, помогли сохранить стране значительное количество хлебных припасов, в которых была острейшая нужда.
Сердце не выдержало

Все виды животные, наземные растения и большинство грибы многоклеточные, как и многие водоросли, тогда как некоторые организмы частично одноклеточные и частично многоклеточные, например слизевые формы и социальные амебы, такие как род Диктиостелиум. [2] [3]
Содержание
Эволюционная история
Вхождение
Многоклеточность независимо развивалась у эукариот как минимум 25 раз, [7] [8] а также в некоторых прокариоты, подобно цианобактерии, миксобактерии, актиномицеты, Magnetoglobus multicellularis или же Methanosarcina. [3] Однако сложные многоклеточные организмы эволюционировали только в шести эукариотический группы: животные, грибы, бурые водоросли, красные водоросли, зеленые водоросли, и наземные растения. [9] Он неоднократно развивался для Хлоропластида (зеленые водоросли и наземные растения), один или два раза для животных, один раз для бурых водорослей, трижды для грибы (хитриды, аскомицеты и базидиомицеты) [10] и, возможно, несколько раз для слизевые формы и красные водоросли. [11] Первые свидетельства многоклеточности взяты из цианобактерии-подобные организмы, жившие 3–3,5 миллиарда лет назад. [7] Для воспроизводства истинные многоклеточные организмы должны решить проблему регенерации всего организма из стволовые клетки (т.е. сперма и яйцо ячеек), вопрос, который изучается в эволюционная биология развития. Животные развили значительное разнообразие типы клеток в многоклеточном теле (100–150 различных типов клеток) по сравнению с 10–20 у растений и грибов. [12]
Потеря многоклеточности
В некоторых группах произошла потеря многоклеточности. [13] Грибы преимущественно многоклеточные, хотя ранние расходящиеся линии в основном одноклеточные (например, Микроспоридии), и было множество случаев возврата к одноклеточности грибов (например, Сахаромикотина, Криптококк, и другие дрожжи). [14] [15] Это могло также произойти в некоторых красные водоросли (например., Порфиридий), но не исключено, что они примитивно одноклеточные. [16] Утрата многоклеточности также считается вероятной в некоторых странах. зеленые водоросли (например., Хлорелла обыкновенная и немного Ulvophyceae). [17] [18] В других группах, обычно паразитов, произошло снижение многоклеточности по количеству или типам клеток (например, микозойные, многоклеточные организмы, которые ранее считались одноклеточными, вероятно, чрезвычайно редки. книдарийцы). [19]
Многоклеточные организмы, особенно долгоживущие животные, сталкиваются с проблемой: рак, который возникает, когда клетки не могут регулировать свой рост в рамках нормальной программы развития. Во время этого процесса можно наблюдать изменения морфологии тканей. Рак у животных (многоклеточные животные) часто описывается как потеря многоклеточности. [20] Ведется дискуссия о возможности существования рака у других многоклеточных организмов. [21] [22] или даже у простейших. [23] Например, галлы растений были охарактеризованы как опухоли, [24] но некоторые авторы утверждают, что у растений не развивается рак. [25]
Разделение соматических и половых клеток
В некоторых многоклеточных группах, которые называются Вейсманисты, разделение стерильных Соматическая клетка линия и половая клетка линия развивалась. Однако развитие вейсманистов относительно редко (например, позвоночные, членистоногие, Volvox), поскольку большая часть видов обладает способностью к соматический эмбриогенез (например, наземные растения, большинство водорослей, многие беспозвоночные). [26] [27]
Гипотезы происхождения

"> Воспроизвести медиа
Одна из гипотез происхождения многоклеточности заключается в том, что группа функционально-специфичных клеток агрегировалась в подобную слизню массу, называемую Grex, который перемещался как многоклеточная единица. Это по сути то, что слизевые формы делать. Другая гипотеза состоит в том, что примитивная клетка претерпела деление ядра, став тем самым ценоцит. Затем вокруг каждого ядра (а также клеточного пространства и органелл, занятых в нем) образуется мембрана, в результате чего образуется группа связанных клеток в одном организме (этот механизм наблюдается в Дрозофила). Третья гипотеза состоит в том, что при разделении одноклеточного организма дочерние клетки не могли разделиться, что привело к скоплению идентичных клеток в одном организме, который впоследствии мог развить специализированные ткани. Это то, что растение и животное эмбрионы делать так же как колониальный хоанофлагелляты. [28] [29]
Поскольку первые многоклеточные организмы были простыми, мягкими организмами без костей, раковин или других твердых частей тела, они плохо сохранились в летописи окаменелостей. [30] Одним исключением может быть демосубка, который, возможно, оставил химический след в древних породах. Самые ранние окаменелости многоклеточных организмов включают оспариваемые Грипания спираль и окаменелости черных сланцев Палеопротерозойский Ископаемое Франсвильской группы B Формирование в Габон (Габонионта). [31] В Формация Доушантуо дал микрофоссилии возрастом 600 миллионов лет с признаками многоклеточных свойств. [32]
До не давнего времени, филогенетический реконструкция прошла анатомический (особенно эмбриологический) сходства. Это неточно, так как живые многоклеточные организмы, такие как животные и растения удалены от своих одноклеточных предков более чем на 500 миллионов лет. Такое течение времени позволяет обоим расходящийся и сходящийся время эволюции, чтобы имитировать сходства и накапливать различия между группами современных и вымерших видов-предков. Современная филогенетика использует сложные методы, такие как аллоферменты, спутниковая ДНК и другие молекулярные маркеры для описания черт, общих для отдаленно родственных линий. [ нужна цитата ]
Эволюция многоклеточности могла происходить разными путями, некоторые из которых описаны ниже:
Симбиотическая теория
Эта теория предполагает, что первые многоклеточные организмы произошли от симбиоз (кооперация) разных видов одноклеточных организмов, каждый из которых выполняет разные роли. Со временем эти организмы станут настолько зависимыми друг от друга, что не смогут выжить независимо, что в конечном итоге приведет к объединению их геномов в один многоклеточный организм. [33] Каждый соответствующий организм станет отдельной линией дифференцированных клеток внутри вновь созданного вида.
Этот вид сильно зависимого симбиоза можно наблюдать часто, например, во взаимоотношениях между рыба-клоун и Морские анемоны Ритерри. В этих случаях крайне сомнительно, что какой-либо вид выживет очень долго, если другой вымрет. Однако проблема этой теории заключается в том, что до сих пор не известно, как ДНК каждого организма может быть объединена в один геном составить их как единый вид. Хотя теоретически такой симбиоз имел место (например, митохондрии и хлоропласты в клетках животных и растений -эндосимбиоз), это случалось крайне редко, и даже тогда геномы эндосимбионтов сохранили элемент различия, отдельно реплицируя их ДНК во время митоз вида-хозяина. Например, два или три симбиотических организма, образующие совокупность лишайник, хотя и зависят друг от друга в плане выживания, они должны воспроизводиться по отдельности, а затем реформироваться, чтобы снова создать один индивидуальный организм.
Клеточная (синцитиальная) теория
Эта теория утверждает, что один одноклеточный организм с множеством ядра, мог развиться внутренняя мембрана перегородки вокруг каждого своего ядра. [34] Многие протисты, такие как инфузории или же слизевые формы может иметь несколько ядер, оказывающих поддержку этому гипотеза. Однако простого присутствия нескольких ядер недостаточно для подтверждения теории. Множественные ядра инфузорий отличаются друг от друга и имеют четко дифференцированные функции. В макронуклеус обслуживает потребности организма, а микронуклеус используется для полового размножения с обменом генетическим материалом. Формы для слизи синцития формируются из отдельных амебоидных клеток, как синцитиальные ткани некоторых многоклеточных организмов, а не наоборот. Чтобы считаться действительной, эта теория нуждается в наглядном примере и механизме образования многоклеточного организма из уже существующего синцития.
Колониальная теория
Колониальная теория Геккель, 1874, предполагает, что симбиоз многих организмов одного вида (в отличие от симбиотическая теория, что предполагает симбиоз разных видов) привело к созданию многоклеточного организма. По крайней мере, некоторые из них, как предполагается, возникли на суше, многоклеточность возникает за счет разделения и последующего соединения клеток (например, ячеистые слизевые формы), тогда как для большинства многоклеточных типов (тех, которые развились в водной среде) многоклеточность возникает как следствие того, что клетки не могут разделиться после деления. [35] Механизм образования последней колонии может быть настолько простым, насколько и неполным. цитокинез, хотя многоклеточность также обычно рассматривается как клеточная дифференциация. [36]

Теория синзооспор
Некоторые авторы предполагают, что возникновение многоклеточности, по крайней мере, у Metazoa, произошло в результате перехода от временного к пространственному. дифференциация клеток, а не через постепенную эволюцию клеточной дифференциации, как утверждается в ГеккельС Теория гастреи. [38]
GK-PID
Около 800 миллионов лет назад [39] незначительное генетическое изменение в одной молекуле, называемое гуанилаткиназа домен взаимодействия с белками (GK-PID), возможно, позволил организмам перейти от одного клеточного организма к одной из многих клеток. [40]
Роль вирусов
Гены заимствованы из вирусы и мобильные генетические элементы (МГЭ) были недавно определены как играющие решающую роль в дифференцировке многоклеточных тканей и органов и даже в половом размножении, в слиянии яйцеклеток и сперматозоидов. [41] [42] Такие слитые клетки также участвуют в мембранах многоклеточных животных, таких как те, которые предотвращают проникновение химических веществ через плацента и разделение тела мозга. [41] Выявлены два вирусных компонента. Первый синцитин, который произошел от вируса. [43] Второй выявленный в 2007 году называется EFF1, который помогает формировать кожу Caenorhabditis elegans, часть целого семейства белков FF. Феликс Рей из Института Пастера в Париже построил трехмерную структуру белка EFF1. [44] и показано, что он выполняет работу по связыванию одной клетки с другой при вирусных инфекциях. Тот факт, что все известные слитые молекулы клеток имеют вирусное происхождение, предполагает, что они были жизненно важны для систем межклеточной коммуникации, которые обеспечивали многоклеточность. Без способности слияния клеток могли бы образоваться колонии, но ничего, даже столь сложного, как губка, было бы невозможно. [45]
Гипотеза доступности кислорода
Эта теория предполагает, что кислород, доступный в атмосфере ранней Земли, мог быть ограничивающим фактором для появления многоклеточной жизни. [46] Эта гипотеза основана на корреляции между возникновением многоклеточной жизни и повышением уровня кислорода в это время. Это произошло бы после Великое окислительное событие (GOE), но до последнего повышения уровня кислорода. Миллс [47] делает вывод, что количество кислорода, присутствующего во время Эдиакарский не является необходимым для сложной жизни и, следовательно, вряд ли явился движущим фактором возникновения многоклеточности.
Гипотеза снежного кома Земли
А снежный ком Земля это геологическое событие, при котором вся поверхность Земли покрыта снегом и льдом. Самый последний снежный ком на Земле произошел во время Криогенный период и состоял из двух глобальных оледенений, известных как Стуртиан и Мариноанец оледенения. Сяо [48] предполагает, что между периодом времени, известным как "Скучный миллиард"и Земля-снежок, у простой жизни могло быть время для инноваций и развития, что позже могло бы привести к эволюции многоклеточности. Гипотеза Земли-снежного кома в отношении многоклеточности предполагает, что кирогенский период в истории Земли мог быть катализатором эволюции сложной многоклеточной жизни. [49] предполагает, что период между гляцием Стурта и более поздним гляцием Марино позволил планктонным водорослям доминировать в морях, уступив место быстрому разнообразию жизни как для растений, так и для животных. Вскоре после мариноанцев сложная жизнь быстро возникла и разнообразилась в так называемых Кембрийский взрыв.
Гипотеза хищничества
Гипотеза хищничества предполагает, что для того, чтобы избежать поедания хищников, простые одноклеточные организмы развили многоклеточность, что затрудняет их употребление в качестве добычи. Herron et al [50] провели лабораторные эксперименты по эволюции одноклеточной зеленой водоросли, C. reinhardtii, используя парамеций в качестве хищника. Они обнаружили, что в присутствии этого хищника C. reinhardtii действительно развивает простые многоклеточные функции.
Преимущества
Многоклеточность позволяет организму превышать пределы размеров, обычно налагаемые распространение: одиночные клетки с увеличенным размером имеют пониженное отношение поверхности к объему и им трудно усваивать достаточное количество питательных веществ и транспортировать их по клетке. Таким образом, многоклеточные организмы обладают конкурентный преимущества увеличения размера без его ограничений. У них может быть более продолжительная продолжительность жизни, поскольку они могут продолжать жить, когда умирают отдельные клетки. Многоклеточность также позволяет увеличивать сложность, позволяя дифференциация типов клеток в одном организме.
Однако вопрос о том, можно ли считать это преимуществами. Подавляющее большинство живых организмов одноклеточные, и даже с точки зрения биомассы одноклеточные организмы гораздо более успешны, чем животные, но не растения. [51] Вместо того, чтобы рассматривать такие черты, как более продолжительная продолжительность жизни и больший размер, как преимущество, многие биологи рассматривают их только как примеры разнообразия с соответствующими компромиссами.
Читайте также: