
Строение гена эукариот кратко
Обновлено: 04.07.2024
Свидетельство и скидка на обучение каждому участнику
Зарегистрироваться 15–17 марта 2022 г.
Описание презентации по отдельным слайдам:
"Строение гена прокариот и эукариот "
10 класс
Профильный уровень
Экспрессия генов- реализация наследственной информации
Информация о структуре белка содержится
в молекуле ДНК
Ген-участок молекулы ДНК несущий информацию о строении одного белка.
Все клетки одного организма содержат одинаковый набор генов
Но по составу белков они отличаются
Клетки специализированны и каждая синтезирует свойственные им белки
У прокариот весь генетический материал – 1 кольцевая молекула ДНК.
Большую часть этой ДНК составляют структурные гены.
Конститутивные гены – гены, кодирующие постоянно необходимые белки. (Н: для кишечной палочки это гены кодирующие ферменты, расщепляющие глюкозу, или РНК- полимеразу).
Оперон- тесно связанная последовательность структурных генов,определяющих синтез группы белков, которые участвуют в одной цепи биохимических преобразований.
Оперон содержит :
Промотор – посадочная площадка для РНК-полимеразы.
Оператор – участок ДНК, с которого начинается операция – синтез и-РНК.
Оперон
Для прокариот характерны два типа регуляции генной активности
Индуцибельный оперон
Реприсибельный оперон
Н:
Lac-оперон
Н:
Trp- оперон
Индуцибельный оперон
Когда в клетку попадает субстрат, для расщепления которого необходимы ферменты данного оперона:
молекула субстрата связывается с репрессором;
репрессор теряет способность взаимодействовать с оператором, отходит и освобождает дорогу РНК-полимеразе.
РНК-полимераза синтезирует и-РНК.
Когда последняя молекула субстрата будет преобразована, репрессор возвращается на оператор, транскрипция прекращается.
Lac-оперон E. coli содержит 3 гена, отвечающие за образование белков, участвующих в расщеплении лактозы.
В отсутствие в клетке лактозы lac-оперон выключен.
Как только некоторое количество лактозы попадает в клетку, две молекулы субстрата (лактозы) взаимодействуют с репрессором, изменяют его конформацию - и он теряет сродство к оператору.
Тут же начинается транскрипция lac-оперона
Реприсибельный оперон
Триптофановый оперон E. coli состоит их пяти структурных генов, ответственных за синтез пяти ферментов, участвующих в превращении хоризмовой кислоты в триптофан, а также из промоторно-операторной области.
Если в цитоплазме ощущается дефицит триптофана, то белок-репрессор не может связаться с оператором.
РНК-полимераза, транскрибирует структурные гены, и в результате в клетке синтезируется триптофан.
Когда триптофан накапливается выше определенного уровня, он взаимодействует с репрессором и активирует его.
Активированный репрессор присоединяется к оператору и подавляет транскрипцию структурных генов триптофанового оперона.
Синтез триптофана прекращается.
Принцип работы репрессибельного триптофанового оперона (Тrр-оперон)
Клетки эукариот содержат оформленное (ограниченное оболочкой) ядро, и их генетический материал представлен линейными молекулами ДНК – хромосомами.
Строение эукариотического гена
У эукариот участок молекулы ДНК определяет структуру одного белка или молекулы РНК.
Любой ген состоит из двух основных частей
структурной
регуляторной
Эукариотический ген, кодирующий синтез определенного белка, содержит:
Обширную регуляторную зону;
Промотор (80-100 пар нуклеотидов)
Структурную часть гена, состоящую из экзонов и интронов.
Экзоны – участки ДНК, несущие информацию о строении белка.
Интроны – участки ДНК не кодирующие структуру белка; вырезаются при сплайсинге и в зрелую
и-РНК не входят.
Процесс вырезания интронов и сращивание экзонов при образовании и-РНК называется сплайсингом
Процессинг:
Кэпирование
Сплайсинг
Полиаденилирование
Кэпирование – присоединение к 5` концу модифицированного гуанина. Необходимо для связывания с рибосомой
Полиаденилирование – присоединение к 3` концу 200-300 аденозинов. Необходимо для защиты от деградации в цитоплазме и
регуляции времени жизни РНК
КЭП
Экзоны имеют небольшую длину.
Длина интрона может быть разной – от нескольких десятков пар нуклеотидов до многих тысяч.
Общая длина всех интронов часто значительно превышает суммарную длину экзонов.
Энхансеры - элементы регуляторной области генов которые участвуют в формировании комплекса транскрипционных факторов.
Инсуляторы (от англ. insulate – изолировать, отделять) регуляторные участки, разграничивающие соседние гены.
Принцип работы инсуляторов – обеспечение избирательной деятельности энхансеров
На функционирование генов оказывают влияние очень многие белки и рибонуклеопротеиды.
При этом они вступают в разнообразные взаимоотношения друг с другом, а также с прочими веществами, от чего зависит конечное влияние этих белков на активность генов
Механизм инициации транскрипции генов эукариот
Транскрипция (от лат. transcriptio – переписываю) представляет собой процесс перевода наследственной информации из последовательности ДНК в последовательность нуклеотидов РНК.
Осуществляется она путём комплементарного синтеза РНК на матричной цепи ДНК.
Образование молекулы РНК на матричной цепи ДНК по принципу комплементарности так же, как и при редупликации, называют матричным синтезом.

Структурными генами называются участки ДНК, кодирующие белковые цепи, т-РНК и р-РНК.
Экзоны — участки гена, кодирующие структуру полипептида.
Интроны— участки гена, не кодирующие структуру полипептида. Их роль до конца не ясна, вероятно они участвуют в процессах генетической рекомбинации, а также в процессах регуляции экспрессии.
Количество интрон-экзонных переходов в пределах гена может варьироваться от 0 до 50. Колебание размеров более характерно для интронов — от 20 до более чем 10000 п.н.
Регуляторные элементы могут находиться за пределами сайта транскрипции и быть общими для нескольких генов:
— Промоторы — участки присоединения РНК полимеразы
— Энхансеры — усилители транскрипции
— Сайленсеры — ослабители транскрипции
— Инсуляторы — ограничители влияния соседних регуляторных элементов
Полный ответ:
Ген представляет собой последовательность нуклеотидов ДНК размером от нескольких сотен до миллиона пар нуклеотидов, в которых закодирована генетическая информация о первичной структуре белка (число и последовательность аминокислот). Гены являются элементарными дискретными единицами наследственности. Воспроизведение и действие генов непосредственно связано с матричными процессами. В настоящее время ген рассматривается как единица функционирования наследственного материала. Химической основой гена является молекула ДНК.
Гены разделяются на структурные, регуляторные и гены-модуляторы.
Структурные генысодержат информацию о структуре белка (полипептидов) и рибонуклеиновых кислот (рибосомальной и транспортной), при этом генетическая информация реализуется в процессе транскрипции и трансляции или только транскрипции.У человека насчитывается около 30 000 структурных генов, но только часть из них экспрессирована. Размеры генов варьируют от 250 п.н. до 2 200 ООО п.н.
Общепринятая модель строения гена — экзон-интронная структура:
Экзон —постедовательностъ ДНК. которая представлена в зрелой РНК. В составе гена должен присутствовать как минимум один экзон (гены тРНК, гистонов). Максимальное количество экзонов представлено в гене мышеч-ного белка титана — 364 экзона. В среднем в гене содержится 8 экзонов.
• Фактор инициации транскрипции 5 -ACTT(T/C)TG-3' входит в состав первого экзона.
• Фактор терминации транскрипции (менее определённая последовательность) входит в состав последнего экзона.
Интрон — последовательность ДНК. включённая между экзонамн. не входит в состав зрелой РНК. Интроны имеют определенные нуклеотидные последовательности, определяющие их границы с экзонамн: на 5'конце — GU последовательность, на 3'конце —AG. Интроны содержат регуляторные элементы экспрессии гена и могут кодировать регуляторные РНК (miRNA).
Сигнал полиаденилирования 5 -ААГААА-З' входит в состав последнего экзона, начинается сразу после стоп-кодона (ТАА, TAG, TGA). Поли(А) сайты защищают мРНК от деградации.
Копирование гена происходит в направлении 5' -> 3'; на флангах (границах) находятся специфические сайты, ограничивающие ген и содержащие регуляторные элементы его транскрипции.
Регуляторные элементы — промотор, энхансеры, сайленсеры, инсуляторы — могут находиться за пределами сайта транскрипции и быть общими для нескольких генов.
• Промотор (от англ. promoter — активатор, ускоритель) — цис-регуляторная последовательность в 5 -области гена, определяющая место прикрепления РНК-полимеразы и интенсивность (частоту) транскрипции мРНК. Содержит ТАТА-бокс для связывания основного фактора транскрипции TEQD. Проксимальнее ТАТА-бокса содержатся GC бокс (5'-GGCiCGG-3') и СААГ бокс (З'-СС'ААТ-З’) для связывания дополнительных специфических белков, активирующих экспрессию генов. Активация только промотора не достаточна для экспрессии гена на физиологически значимом уровне.
Энхансеры (от англ. enhance—усиливать)—цис-позитизные регуляторные элементы и сайленсеры (от англ. silence — успокаивать) — цис-негативные регуляторные элементы состоят из 6-12 нуклеотидов, специфически взаимодействующих с белками. Энхансеры связываются с белками активаторами и усиливают экспрессию гена.
Сайленсеры связываются с белками репрессорами и блокируют экспрессию гена. Энхансеры и сайленсеры локализуются в 5'- или 3'-фланкируклцих участках, нитронах. Активность не зависит от их ориентации или локализации. Кроме того, они могут находиться на больших расстояниях от промотора (несколько сотен п.и.) и взаимодействуют с ним за счёт образования петель ДНК.

Инсуляторы(англ. MAR — matrix attachment regions). Образуют дискретные функциональные домены — петли хромосом, ограничивающие влияние соседних регуляторных элементов. В состав петли могут входить специфические последовательности, контролирующие локус (англ. LCR — locus-control region) — позитивные цис-элементы, регулирующие активность нескольких генов (рис. 1-2).
65. Классификация генов

По месту локализации генов в структурах клетки различают расположенные в хромосомах ядра, ядерные гены и цитоплазматические гены, локализация которых связана с хлоропластами и митохондриями.
По функциональному значению различают:структурные гены, характеризующиеся уникальными последовательностями нуклеотидов, кодирующих свои белковые продукты, которые можно идентифицировать с помощью мутаций, нарушающих функцию белка, и регуляторные гены - последовательности нуклеотидов, не кодирующие специфические белки, а осуществляющие регуляцию действия гена (ингибирование, повышение активности и др.).
По влиянию на физиологические процессы в клетке различают: летальные, условно летальные, супервитальные гены, гены-мутаторы, гены-антимутаторы и др.
Следует отметить, что любые биохимические и биологические процессы в организме находятся под генным контролем. Так, деление клеток (митоз, мейоз) контролируется несколькими десятками генов; группы генов осуществляют контроль восстановления генетических повреждений ДНК (репарация). Онкогены и гены - супрессоры опухолей участвуют в процессах нормального деления клеток. Индивидуальное развитие организма (онтогенез) контролируется многими сотнями генов. Мутации в генах приводят к измененному синтезу белковых продуктов и нарушению биохимических или физиологических процессов.
Гомеозисные мутации у дрозофилы позволили открыть существование генов, нормальной функцией которых является выбор или поддержание определенного пути эмбрионального развития, по которому следуют клетки. Каждый путь развития характеризуется экспрессией определенного набора генов, действие которых приводит к появлению конечного результата: глаза, голова грудь, брюшко, крыло, ноги и т. д. Исследования генов комплекса bithorax дрозофилы американским генетиком Льюисом показали, что это гигантский кластер тесно сцепленных генов, функция которых необходима для нормальной сегментации груди (thorax) и брюшка (abdomen). Подобные гены получили название гомеобоксных. Гомеобоксные гены расположены в ДНК группами и проявляют свое действие строго последовательно. Такие гены обнаружены и у млекопитающих, и они имеют высокую гомологию (сходство).
66. Отличие в строении прокариот и эукариот.
У прокариот нет ядра, кольцевая ДНК (кольцевая хромосома) расположена прямо в цитоплазме (этот участок цитоплазмы называется нуклеоид). У эукариот есть оформленное ядро (ДНК отделена от цитоплазмы ядерной оболочкой).
У прокариот нет ядра, кольцевая ДНК (кольцевая хромосома) расположена прямо в цитоплазме (этот участок цитоплазмы называется нуклеоид). У эукариот есть оформленное ядро (ДНК отделена от цитоплазмы ядерной оболочкой).
Дополнительные отличия
1) Раз у прокариот нет ядра, то нет и митоза/мейоза. Прокариоты размножаются делением надвое.
2) У прокариот из органоидов имеются только рибосомы (мелкие, 70S), а у эукариот кроме рибосом (крупных, 80S) имеется множество других органоидов: митохондрии, эндоплазматическая сеть, клеточный центр, и т.д.
3) Клетка прокариот гораздо меньше клетки эукариот: по диаметру в 10 раз, по объему – в 1000 раз.
Сходства
Клетки всех живых организмов (всех царств живой природы) содержат плазматическую мембрану, цитоплазму и рибосомы.
Транскрипция у эукариот
Определения: Ядрышко - место образования субъединиц рибосом, наблюдаемое в световой микроскоп. В ядре может быть несколько ядрышек. Мастер генов рРНК называют ядрышковым организатором. Базальные факторы транскрипции - белки, необходимые для инициации транскрипции. Энхансеры - последовательности ДНК, усиливающие транскрипцию при взаимодействии со специфическими белками. Сайленсеры – последовательности ДНК, ослабляющие транскрипцию при взаимодействии с белками.
У эукариот процессы транскрипции и трансляции разобщены во времени и пространстве (транскрипция - в ядре, трансляция - в цитоплазме). У эукариот существуют специализированные РНК-полимеразы. В ядре выделяют 3 типа РНК-полимераз: РНК-полимераза I - синтезирует rРНК (кроме 5SrРНК). РНК-полимераза II - синтезирует мРНК и некоторые sPHK. РНК-полимераза III - синтезирует тPHK, некоторые sPHK и 5SrPHK. РНК-полимеразы различаются количеством субьединиц, их аминокислотным составом, и зависимостью от катионов магния и марганца. Для РНК-полимераз I и III необходимое для работы соотношение [Mn2+]/[Mg2+] = 2. Для РНК-полимеразы II - [Mn2+]/[Mg2+] = 5. Наиболее яркое различие - чувствительность к α- аманитину (токсину бледной поганки). Он полностью подавляет работу РНК-полимеразы II в концентрации 10-8 М и РНК-полимеразы III ( в концентрации 10-6 М). РНК-полимераза I фактически нечувствительна к этому токсину. Помимо ядерных РНК-полимераз у эукариот есть еще РНК-полимеразы хлоропластов и митохондрий. Они кодируются в ядре, а не в соответствующих органеллах. В органеллах образуются свои тPHK, рРНК и рибосомные белки.
Как образуются рибосомы у эукариотГены rРНК присутствуют в количестве от 10 до 105 копий у разных видов (105 у амфибий). У человека - 300 генов, в которых закодированы rРНК.Все рибосомные гены, кроме генов 5S рибосомной РНК, сближены (т.е располагаются один за другим) и образуют несколько кластеров. Сначала синтезируется про-рРНК, после созревания которой образуются 28S, 18S и 5,8SrРНК.Интерфазные хромосомы в световой микроскоп не видны. Каждый ген прорибосомной РНК транскрибируется одновременно несколькими РНК-нолимеразами и тут же начинается процессинг.На электронномикроскопических фотографиях видна картина "рождественской елочки". Синтезируемые в ядре мРНК поступают на готовые рибосомы в цитоплазму, где синтезируются рибосомные белки, которые идут в ядро и путаются в "ветвях елки".Образуются рибосомные субъединицы. Одновременно в эукариотическом ядре находятся сотни тысяч субъединиц рибосом.
Особенности транскрипции эукариот.Единицей транскрипции у эукариот является отдельный ген, а не оперон, как у прокариот.Оператор, как таковой, отсутствует. Промотор есть, но он организован иначе.На расстоянии -25 п.н. от +1 нукл. находится ТАТА-бокс. Его позиция определяет точку инициации транскрипции. А на расстоянии -60-80 п.н. находится ЦААТ-бокс, который не является абсолютно необходимым, но присутствует перед большинством генов.Расстояние между ЦААТ и ТАТА большое и РНК-полимераза не способна накрыть всю эту область.ЦААТ опознается своим белком, а ТАТА - своим.Помимо этих есть еще несколько белков, называемых базальными факторами транскрипции.Базальные факторы транскрипции необходимы для инициации транскрипции всеми тремя ядерными РНК-полимеразами.Для любого гена, кодирующего белок, есть энхансеры (усилители). Энхансеры - это не непрерывные последовательности нуклеотидов. Существуют так называемые модули - это отдельные части энхансеров. Одинаковые модули могут встречаться в разных энхансерах. Для каждого энхансера набор модулей уникален. Модули - это короткие последовательности, не более 2-х витков спирали (20 п.н.), которые могут находиться перед, за и даже внутри гена. Таким образом, М1+М2+МЗ+М4 - один энхансер, но он состоит из 4-х модулей. Все 4 модуля узнаются своими белками, а они, сидя на ДНК, взаимодействуют друг с другом. Если в клетке присутствуют все соответствующие белки, то участку ДНК придается определенная конформация и начинается синтез мPHK. Все соматические клетки многоклеточного эукариотического организма имеют абсолютно одинаковый набор генов. Почему же клетки дифференцированы и специализированы? Дело в том, что все гены работают на фоновом уровне и не имеют фенотипического проявления. Экспрессируются лишь те гены, у которых все энхансерные модули узнаны своими белками и эти белки взаимодействуют друг с другом. Кроме энхансеров есть сайленсеры (ослабители). При соответствующем наборе белков экспрессия отдельных генов в клетке может быть пода
Механизм созревания мРНК
Транскрипция (от лат. transcriptio — переписывание) — процесс синтеза РНК с использованием ДНК в качестве матрицы, происходящий во всех живых клетках. Другими словами, это перенос генетической информации с ДНК на РНК.
Экспрессия генов — это процесс, в ходе которого наследственная информация от гена (последовательности нуклеотидов ДНК) преобразуется в функциональный продукт — РНК или белок. Некоторые этапы экспрессии генов могут регулироваться: это транскрипция, трансляция, сплайсинг РНК и стадия посттрансляционных модификаций белков.
Сплайсинг— это процесс, в котором из пре-мРНК удаляются участки, не кодирующие белок, называемые интронами; последовательности, которые остаются, несут информацию о структуре белка и называются экзонами. Иногда продукты сплайсинга пре-мРНК могут быть соединены разными способами, позволяя одному гену кодировать несколько белков. Этот процесс называется альтернативным сплайсингом.
Схема сплайсинга
Трансляяция — процесс синтеза белка из аминокислот на матрице информационной (матричной) РНК (иРНК, мРНК), осуществляемый рибосомой.
Транскрипция осуществляется ферментом РНК-полимеразой, строящей, согласно принципу комплементарности, копию участка ДНК на основании одной из цепей двойной спирали. Этот процесс как у эукариот, так и у прокариот организован одинаково. Основное различие между про- и эукариотами состоит в том, что у эукариот РНК-полимераза во время транскрипции ассоциируется с мРНК-обрабатывающими ферментами, поэтому у них обработка мРНК и транскрипция могут проходить одновременно. Короткоживущие необработанные или частично обработанные продукты транскрипции называются пре-мРНК; после полной обработки — зрелая мРНК.
Кэпирование является первым этапом процессинга мРНК. Оно осуществляется, когда синтезируемый транскрипт достигает длины 25—30 нуклеотидов. Сразу после присоединения кэпа к 5'-концу транскрипта с ним связывается кэп-связывающий комплекс CBC (англ. cap binding complex), который остаётся связанным с мРНК до завершения процессинга и важен для всех последующих его этапов. В процессе сплайсинга из пре-мРНК удаляются не кодирующие белок последовательности — интроны. Полиаденилирование необходимо для транспорта большинства мРНК в цитоплазму и защищает молекулы мРНК от быстрой деградации (увеличивает время их полужизни). Лишённые поли(А)-участка молекулы мРНК (например, вирусные) быстро разрушаются в цитоплазме клеток эукариот рибонуклеазами.
После завершения всех стадий процессинга мРНК проходит проверку на отсутствие преждевременных стоп-кодонов, после чего она становится полноценной матрицей для трансляции. В цитоплазме кэп узнаётся факторами инициации, белками, отвечающими за присоединение к мРНК рибосомы, полиадениновый хвост связывается со специальным поли(А)-связывающим белком PABP1.
Ген — участок молекулы ДНК, кодирующий первичную последовательность аминокислот в полипептиде или последовательность нуклеотидов в молекулах транспортных и рибосомных РНК. ДНК одной хромосомы может содержать несколько тысяч генов, которые располагаются в линейном порядке. Место гена в определенном участке хромосомы называется локусом. Особенностями строения гена эукариот являются: 1) наличие достаточно большого количества регуляторных блоков, 2) мозаичность (чередование кодирующих участков с некодирующими). Экзоны (Э) — участки гена, несущие информацию о строении полипептида. Интроны (И) — участки гена, не несущие информацию о строении полипептида. Число экзонов и интронов различных генов разное; экзоны чередуются с интронами, общая длина последних может превышать длину экзонов в два и более раз. Перед первым экзоном и после последнего экзона находятся нуклеотидные последовательности, называемые соответственно лидерной (ЛП) и трейлерной последовательностью (ТП). Лидерная и трейлерная последовательности, экзоны и интроны образуют единицу транскрипции. Промотор (П) — участок гена, к которому присоединяется фермент РНК-полимераза, представляет собой особое сочетание нуклеотидов. Перед единицей транскрипции, после нее, иногда в интронах находятся регуляторные элементы (РЭ), к которым относятся энхансеры и сайленсеры. Энхансеры ускоряют транскрипцию, сайленсеры тормозят ее.
Гены прокариот состоят из двух основных элементов: регуляторной части и собственно кодирующей части(рис. 27). Регуляторная часть обеспечивает первые этапы реализации генетической информации, а кодирующая часть содержит информацию о структуре полипептида, тРНК, рРНК. У прокариот структурные гены, кодирующие белки одноо метаболического пути, часто бывают объединены и называются опероном. Так, например, в лактозном опероне E. coli содержится 3 структурных гена. Для биосинтеза аминокислоты гистидин требуется 9 ферментов и ее оперон содержит 9 структурных генов.
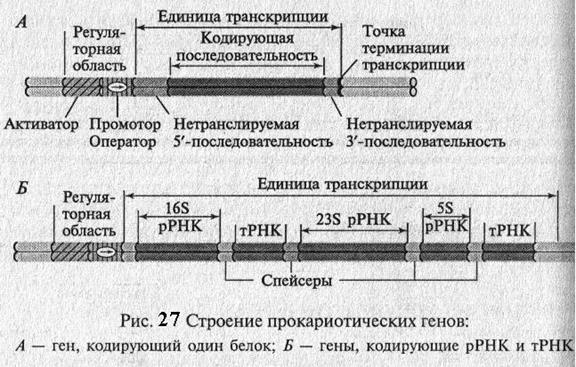
Гены, кодирующие белки, обычно содержат на 5'- и 3'- концах гена или оперона нетранслируемые последовательности (5' – НТП и 3' – НТП), которые играют важную роль в стабилизации иРНК. Гены тРНК и рРНК отделены друг от друга спейсерами (от англ. – spacer – распорка), т.е. последовательностями, которые вырезаются в ходе их созревания (процессинга)(рис. 27).
(А. С. Коничев, Г. А. Севастьянова, 2005, с. 157)
Экзоны – это последовательности участвующие в образовании зрелых РНК. Они могут быть как кодирующие так и некодирующие. Наследственная информация экзонов реализуется в синтезе определенных белков, а роль интронов до конца еще не выяснена.
Возможное значение интронов:
1. Интроны снижают частоту мутаций, соотношение интронов и экзонов у человека 3:2.
2. Интроны поддерживают структуру ДНК, т.е. играют конститутивную роль.
3. Интроны необходимы для процесса созревания иРНК. Без интронов нарушен выход иРНК в цитоплазму. При введение в ядро искусственной иРНК без интронов, она остается в ядре и в цитоплазму не выходит.
4. В последние годы четко установлено, что некоторые интроны кодируют белки – ферменты, которые их вырезают.
5. Превращаются в малые ядерные РНК (мяРНК).

(А. С. Коничев, Г. А. Севастьянова, 2005 г., с. 157)
Гены высших организмов чаще оказываются прерывистыми, но есть и непрерывистые, например, гены интерферонов, гены гистонов. Степень прерывистости может быть различной – от одного интрона как у гена актина до нескольких десятков, как у гена коллагена(рис.29).

Рис. 29. Карты некоторых прерывистых генов. Жирные линии – экзоны, тонкие - интроны (А. С. Коничев, Г. А. Севастьянова, 2005 г., с. 158)
Длина интронов часто оказывается длинней экзонов: 5 – 20 тыс. и 1 тыс. соответственно. Прерывистость гена считалось достоянием эукариот. Но в 1983г. группа ВЕЗЕ (США)обнаружила их у некоторых археобактерий. Интроны содержаться во всех типах РНК, Интроны в составе иРНК вырезаются при участии мяРНП, которые образуют с интроном сплайсосому. При помощи сплайсосом, узнается начало и конец интрона, их концы соединяются в цепи РНК и интрон вырезается (рис.32).
Эволюционное возникновение мозаичной (итрон – экзонной) структуры генов эукариот в настоящее время не находит объяснения. С точки зрения У. Гильберта появление интронов обеспечило возможность обмена экзонами между неродственными генами. В результате это привело к появлению белков с новыми функциями (гипотеза позднего возникновения интронов). По другой гипотезе интроны это эволюционные реликты, они были частью гигантских генов. Прокариоты являются эволюционным тупиком т.к. не содержат интронов.
Регуляция работы гена
Гены функционируют в клетке не сами по себе, а входят в состав более сложной генной регуляторной системы. Количество структурных генов в разных оперонах различно. Участок ДНК, на на котором проходит считывание информации, называется единицей транскрипции (Рис. 27, 28). Он ограничен промотором (зона начала транскрипции) и терминатором (зона остановки транскрипции).
1. Промотор – это строго определенная нуклеотидная последовательность, которая узнается ферментом транскрипции – РНК – полимеразой.
У E. coli промотор – это пара нуклеотидных последовательностей из 6-7 и 9 нуклеотидов каждая, отдельных друг от друга 25 нуклеотидами.
Промотор выполняет следующие функции:
а) это место присоединения РНК – полимеразы к молекуле ДНК
У про- и эукариот последовательности промоторов разные. Это учитывается в генной инженерии, в случае встраивания в геном бактерии генов человека.
Промоторы эукариот разнообразны по числу и строению элементов. Промотор эукариотического гена – это участок ДНК, на котором собираются белки транскрипции, узнающие свои сайты связывания и взаимодействующие друг с другом и с иРНК – полимеразой. В составе эукариотического гена имеются особые цис-действующие элементы регуляции - усилители или активаторы, глушители или угнетатели транскрипции. Они разнообразны по строению, положению и функциям. Они могут располагаться как на 5'- так и на3'- конце фрагмента ДНК, включающего ген, так и в составе интронов.
2. Оператор – это нетранскрибирумая последовательность нуклеотидов, участок связывания белка - репрессора. Он располагается в непосредственной близости к промотору или перекрывается с ним. У многих опероновимеется не один, а несколько сайтов связывания с регуляторными белками, которые не обязательно располагаются рядом, а могут находиться по разные стороны от промотора. Поэтому сейчас принято говорить о сайтах связывания регуляторов. Связывание белка – репрессора с оператором либо создает стерические (пространственные) затруднения для связывания РНК-полимеразы с промотором, либо препятствует продвижению ее по смысловой цепи ДНК, и определяет точку начала транскрипции. Следует отметить, что ни промотор, ни оператор в РНК не транскрибируются и зоны промотора и оператора могут перекрываться.
3. Терминатор – участок молекулы ДНК, где заканчивается процесс транскрипции.
Оператор и структурные гены образуют оперон. Именно так назвали эту структуру французские ученые Франсуа Жакоб и Жак Моно, которые первыми в 1959 -1961г. работая с бактериальными клетками предложили механизм регуляции работы гена или генной экспрессии. За эту работу в 1965г. они получили Нобелевскую премию. Как выяснили Жакоб и Моно работой оперона управляют гены – регуляторы. Они не входят в состав оперона, но являются необходимой частью регуляторной системы. Гены – регуляторы у прокариот находятся на той же хромосоме, что и оперон. У эукариот они могут располагаться далеко от промотора эукариотического гена и оказывать дистанционное влияние на его транскрипцию. Гены - регуляторы контролируют синтез белка – репрессора связывающегося с оператором. Синтез белков – репрессоров, как и всех белков, идет на рибосомах в цитоплазме. Транскрипция определяется белком-репрессором, который может закрывать оператор (репрессор активен) или открывать его (репрессор неактивен), т.е. возможны два варианта регуляции активности генов.
I. Ген – регулятор отвечает за синтез активного белка – репрессора. Белок – репрессор имеет два активных центра:
1. центр связывания с оператором
2. центр связывания с субстратом. Под субстратом (индуктором) понимают любое вещество, информация о синтезе или распаде которого закодирована в данном опероне или гене. Это могут быть гормоны, аминокислоты, углеводы, питательные вещества, яды и т.д.
Субстрата в клетке нет, поэтому активная форма белка – репрессора соединяется с оператором, т.е. оператор закрыт и через него не может пройти фермент РНК – полимераза, транскрипция не идет (рис.30). Открытие оператора идет с помощью субстрата (индуктора), поступающего в клетку. Индуктор взаимодействует с белком – репрессором, что приводит к изменению его конформации (пространственной структуры). У инактивированного белка репрессора резко снижается родство к зоне оператора и он отсоединяется от него. Оператор свободен и это позволяет РНК-полимеразе начать транскрипцию. Она продолжается до тех пор, пока в клетке есть субстрат, т.е. пока есть необходимость в продуктах данного оперона или гена (рис.31).
При сокращении количества субстрата его уже не хватает на молекулы белка – репрессора и активный белок – репрессор присоединяется к оператору. Транскрипция прекращается. Следует отметить, что в клетке белок реперссор синтезируется постоянно и его количество строго определенное. Например, в клетке E. сoli находится около 10 молекул белка-репрессора, который регулирует работу лактозного оперона.
II. Ген – регулятор отвечает за синтез неактивной формы белка – репрессора, т.е. он не может присоединится к оператору. Оператор свободен иРНК – полимераза свободно проходит к структурным генам. Оперон будет работать до тех пор, пока есть необходимость в продуктах данного оперона.
Когда данный продукт клетке уже не нужен (он не расходуется в биохимических процессах и накапливается в клетке), субстрат взаимодействует с неактивным белком – репрессором, активирует его. Белок-репрессор закрывает оператор и выключает транскрипцию.
Такой способ регуляции метаболизма в клетке чрезвычайно экономичен, т.к. клетка синтезирует продукт в таком количестве, которое необходимо для поддержания определенного уровня обменных процессов. При избытке конечного продукта данный метаболический путь выключается. То есть, мы видим взаимодействие между внутриклеточной средой и генетическим аппаратом для обеспечения тонкой регуляции клеточного метаболизма.
Например, образование некоторых ферментов индуцируется присутствием их субстрата:
1. наличие в крови алкоголя индуцирует в клетках печени усиленный синтез фермента, разрушающего алкоголь – алкогольдегидрогеназы.
2. действие половых гормонов при формировании вторичных половых признаков также основано на усилении транскрипции определенных генов.
3. по такому типу работают гены железистых клеток, вырабатывающие секреты для жизнедеятельности организма.
Если у бактерии на включение гена в работу требуется несколько минут, то у эукариот от нескольких часов до нескольких дней.
Включение и работа генов и оперонов зависят от ряда факторов:
1. Специализации клетки
2. Физиологического состояния
3. Возраста клетки
4. Условий внешней среды
5. Пространственной структуры ДНК (изгибы, петли, сверхспирали и т.д.)
6. Степени метилирования генов.
Показано, что гены материнских и отцовских хромомсом могут быть метилированы по – разному и это регулирует активность разных генов. Например, ген – индуцирующий образование опухолей. Если он передается потомству от отца, то транскрибируется только в сердце, а если от матери, то он вообще не экспрессируется. Исследования показали, что у самок этот ген метилирован, а у самцов – деметилирован.
Любой из этих факторов может оказать существенное влияние на процесс считывания генетической информации.
Читайте также: