
Резонансная теория гельмгольца кратко
Обновлено: 05.07.2024
В 1863 г. Гельмгольц предложил резонансную теорию. Он считал, что в улитке происходят явления механического резонанса, в результате которого сложные звуки в ней разлагаются на простые тоны. То обстоятельство, что основная мембрана с ее эластическими волокнами, натянутыми в поперечном направлении, имеет разную ширину у основания и верхушки улитки (у основания — узкая, у верхушки — широкая), позволило Гельмгольцу считать ее образованием, разные участки которого способны резонировать на звуки неодинаковой высоты.
Теория Гельмгольца с гениальной простотой разъяснила основные свойства уха, т. е. определение высоты, силы и тембра. Согласно резонансной теории, любой чистый тон имеет свой ограниченный участок на основной мембране.
Одиночный звук, по его мнению, раздражает строго определенные нервные волокна — именно те, которые снабжают соответствующий участок мембраны, и раздражение этих волокон ощущается как звук строго определенной высоты.
Резонансной теорией легко объясняется различение тембра звука и способность уха разлагать сложный звук на его составные части. Каждый сложный звук, согласно этой теории, раздражает столько точек на основной мембране, сколько в нем содержится синусоид, т. е. основной тон и все обертоны вызывают соколебание в соответствующих участках мембраны. Все сигналы из внутреннего уха, дошедшие по системе проводников до слуховых центров, интегрируются в них, и мы слышим соответственный тембровый звук. Сила звука определяется числом раздражаемых нервных элементов. Естественно, чем сильнее звук, тем более широкий участок основной мембраны приходит в соколебание. Гельмгольц допускал резонанс отдельных участков мембраны, но не говорил о резонансе свободно колеблющихся струн. Таким образом, из теории Гельмгольца следует три основных вывода:
1) в улитке происходит первичный анализ звуков;
2) каждый простой звук имеет свой участок на основной мембране;
3) низкие звуки вызывают колебание участков основной мембраны у верхушки улитки, а высокие — у основания ее.
Несмотря на огромное количество новых фактов, полученных при изучении функции внутреннего уха, эти три вывода сохраняют свое значение до настоящего времени.
Бекеши теория слуха - Теория, объясняющая первичный анализ звуков в улитке сдвигом столба пери- и эндолимфы и деформацией основной мембраны при колебаниях основания стремени, распространяющихся по направлению к верхушке улитки в виде бегущей волны.
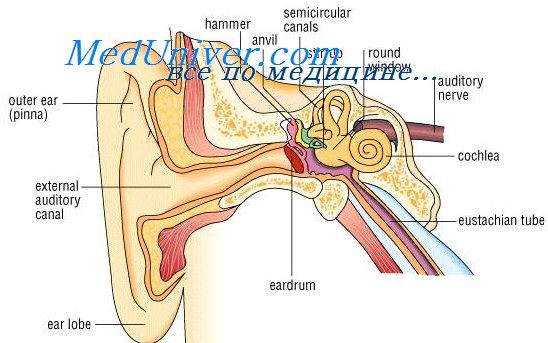
Анатомия, физиология полукружных каналов и преддверия лабиринта.
УЧЕБНИК СТР 337 (АНАТОМИЯ)
Перепончатые полукружные каналы расположены внутри костных и открываются 3 ампулярными и 2 простыми концами, т. е. 5 отверстиями, в utriculus. На поперечном разрезе полукружного канала видно, что перепончатый канал прилежит к одной из костных стенок. В перилимфатическом пространстве имеются поддерживающие соединительнотканные перемычки, по которым идут сосуды.
Более сложно устроены ампулярные концы перепончатого лабиринта, почти целиком заполняющие костные расширения. В них находятся ампулярные гребешки, на которых размещены опорные и нейроэпителиальные волосковые клетки (рецепторный аппарат). Особенностью строения последних является большая длина волосков, склеенных между собой эластической массой и образующих так называемую купулу. Это образование возвышается почти до полного соприкосновения с противоположной стенкой ампулы. Малейшее перемещение эндолимфы в канале неминуемо влечет за собой отклонение купулы, которая затем медленно возвращается в исходное положение.
К ампулярному гребешку подходят ампулярные веточки вестибулярного нерва.
В углублениях преддверия помещаются два перепончатых мешочка — эллиптический и круглыЙ. Утрикулюс расположен в верхне-задней части преддверия, а саккулюс — внизу, вблизи основного завитка улитки. Оба мешочка содержат рецепторный аппарат, расположенный на статическом пятнышке. Оно состоит из опорных и нейроэпителиальных волосковых клеток, короткие волоски которых заключены в желатинообразную массу; на ней покоится отолитовая мембрана с входящими в ее состав кристаллами солей кальция.
Оба мешочка соединены между собой посредством тонкого канала, который продолжается в эндолимфатический проток. Последний помещается в костном канале — водопроводе преддверия и заканчивается расширением — эндолимфатическим мешком, лежащим в дупликатуре твердой мозговой оболочки на задней поверхности пирамидки.
Теория Гельмгольца с гениальной простотой разъяснила основные свойства уха, т. е. определение высоты, силы и тембра. Согласно резонансной теории, любой чистый тон имеет свой ограниченный участок на основной мембране.
Одиночный звук, по его мнению, раздражает строго определенные нервные волокна — именно те, которые снабжают соответствующий участок мембраны, и раздражение этих волокон ощущается как звук строго определенной высоты.
Резонансной теорией легко объясняется различение тембра звука и способность уха разлагать сложный звук на его составные части. Каждый сложный звук, согласно этой теории, раздражает столько точек на основной мембране, сколько в нем содержится синусоид, т. е. основной тон и все обертоны вызывают соколебание в соответствующих участках мембраны. Все сигналы из внутреннего уха, дошедшие по системе проводников до слуховых центров, интегрируются в них, и мы слышим соответственный тембровый звук. Сила звука определяется числом раздражаемых нервных элементов. Естественно, чем сильнее звук, тем более широкий участок основной мембраны приходит в соколебание. Гельмгольц допускал резонанс отдельных участков мембраны, но не говорил о резонансе свободно колеблющихся струн. Таким образом, из теории Гельмгольца следует три основных вывода:
1) в улитке происходит первичный анализ звуков;
2) каждый простой звук имеет свой участок на основной мембране;
3) низкие звуки вызывают колебание участков основной мембраны у верхушки улитки, а высокие — у основания ее.
Несмотря на огромное количество новых фактов, полученных при изучении функции внутреннего уха, эти три вывода сохраняют свое значение до настоящего времени.
Первый вывод вполне гармонирует с учением И. П. Павлова о способности к первичному анализу как концевых приборов афферентных нервов, так и в особенности сложных рецепторных аппаратов. Подтверждением вывода о пространственной локализации звуков в улитке явились опыты, предпринятые Л. А. Андреевым. Он вырабатывал у собак слюнной условный рефлекс на ряд тонов. После полного разрушения лабиринта одного уха он изолированно разрушал то основание, то верхушку улитки другого уха и обнаруживал выпадение условного рефлекса слюноотделения то на высокие, то на низкие тоны. Этим было доказано, что в улитке имеет место первичный анализ звуков.
Все работы последнего времени направлены скорее не на отрицание резонансной теории слуха, а на дальнейшее ее углубление и развитие. Новые наблюдения говорят в пользу того, что под влиянием звуков в лимфе улитки происходят сложные гидродинамические процессы и деформации мембран в улитке зависят от них не в меньшей степени, чем от механических свойств самой основной перепонки (Бекеши, Флетчер). При быстрых колебаниях подножной пластинки относительно большая инерция столба лимфы в обеих лестницах не позволяет ему следовать за быстрыми колебаниями стремени. Это обстоятельство и возрастающее трение в scala vestibuli при быстрых колебаниях ведут к такому повышению давления в этом канале, что рейснерова мембрана, а вслед за ней и основная мембрана прогибаются и колебание передается дальше на лимфу барабанной лестницы и на мембрану круглого окна. Чем выше звук, тем ближе к круглому окну (т. е. основанию) получается прогиб основной перепонки. Самые низкие звуки вызывают деформацию ее у верхушки, т. е. поблизости от геликотремы.
Таким образом, за основу для слуховых теорий в настоящее время принимается теория пространственного расположения (place theory) звуков, согласно которой любому тону соответствует определенный участок на основной мембране.
Как видно из изложенного выше, новейшие теории объясняют избирательное отношение основной мембраны к звукам разной высоты не столько механическими свойствами этой мембраны, ее резонансом, сколько сложными явлениями в ушной лимфе, среди которых главное место занимает перемещение ее столба в улитковых ходах. Это перемещение лимфы передается податливым перепончатым образованием, которые деформируются на большем или меньшем протяжении.
Новейшие опыты Бекеши на моделях и на улитке морской свинки действительно показали, что основная мембрана совершает сложные колебания — при высоких звуках волны деформации захватывают главным образом основные завитки, при низких — всю мембрану. Места максимальной деформации соответствует пространственному расположению звуков на основной перепонке, в этих участках наблюдались вихревые движения лимфы.
Трудности доказательства правильности пространственной теории возникают потому, что все перечисленные выше наблюдения не дают основания утверждать, что под влиянием чистого звука возникает изолированное раздражение столь малого по протяженности участка — в 0,02 мм. Приходится считаться с воздействием каждого тона на гораздо большем протяжении основной мембраны с максимальной деформацией ее в определенной точке. При этом трудно объяснить, почему ощущается только один тон, так как раздражаются и рядом лежащие участки кортиева органа.

Для объяснения этих фактов приходится пользоваться гипотезами, затрагивающими механизм трансформации механической энергии в нервное возбуждение.
Бекеши, Флетчер и др. допускают, что ощущение высоты возникает благодаря раздражению тех нервных приборов, которые расположены в точке максимального изгиба перепонки; нервные же процессы волосковых клеток, находящихся рядом, при этом тормозятся (эффект контраста).
Другое затруднение состоит в том, что с механической точки зрения (т. е. одной только степенью деформации) невозможно объяснить огромное (в миллионы раз) различие чувствительности уха к разным частотам.
Это затруднение отпадает, если допустить, как это делает П. П. Лазарев, что при механическом раздражении волосковых клеток в них возникает химическая реакция, сила которой зависит от количества разлагающегося вещества (слухового пурпура). При этом освобождаются ионы, которые и вызывают процесс нервного возбуждения.
По новейшим данным, в волосковых клетках всегда имеются запасы гликогена, количество которого уменьшается под влиянием звуковой нагрузки (Я. А. Винников).
О глубоких химических изменениях в спиральном ганглии после сильного звукового воздействия сообщают Хиден, Хамбергер и Нильсон (Hyden, Hamberger, Nilsson). При помощи цитохимического способа и фотографирования в лучах коротковолнового спектра (2670 А) они обнаружили уменьшение содержания рибонуклеиновой кислоты и протеина в ганглиозных клетках, в то время как липоидные фракции клеток количественно оказались неизмененными. Роль медиатора в кортиевом органе играет ацетилхолин (Гиссельсон — Giesselson).
Ввиду тех затруднений, которые оказались не вполне объяснимыми пространственной теорией, некоторые авторы (Уивер — Wever, Pебул — Reboul и др.) допускают, что различение высоты определяется двумя факторами — фактором пространственного расположения рецепторных образований и фактором времени, т. е. числом импульсов в секунду. Последнее вполне правдоподобно для частот до 700-1000 Гц, так как такой ритм в неискаженном виде обнаруживается в проводниковой системе. Правильная передача частот нарушается при более высоких звуках, поэтому для них пространственный фактор играет преобладающую роль.
Успехи электрофизиологии органов чувств дают некоторые новые данные о процессах, совершающихся в нервных проводниках и корковых центрах соответствующих анализаторов.
При действии звука в улитке возникают электрические потенциалы — микрофонные токи улитки.
Улитковые (микрофонные) токи повторяют сложную кривую звуковой волны как в отношении амплитуды, так и частоты колебаний до 10 000 Гц и выше. Они возникают в районе основной мембраны, мало изменяются от действия наркотических веществ, при утомлении и улавливаются лучше всего в тех точках, куда благодаря электропроводности тканей они легко проникают. Например, высокочастотные токи особенно хорошо отводятся с мембраны круглого окна.
Микрофонные токи улитки нельзя смешивать с акционными токами, возникающими в нервных образованиях при их возбуждении. При отведении потенциалов с круглого окна всегда получается смесь микрофонных токов и акционных токов слуховых волокон. По времени микрофонные токи улитки возникают несколько раньше, чем акционные токи со слуховых волокон. Токи действия волокон слухового нерва очень чувствительны к действию наркотиков, холода и расстройству кровообращения; при раздражении слухового нерва обнаруживается рефрактерная фаза, т. е. одиночные волокна его передают не больше 500-800 импульсов в секунду. Таким образом, частота нервных импульсов в волокнах слухового нерва не является повторением частоты звуковых колебаний, а представляет собой информацию о различных качествах звука, которая окончательно расшифровывается в корковых концах звукового анализатора.
На основании последних микроэлектрофизиологических исследований рецепции, проведенных Гранитом (Granit), можно думать, что потенциал улитки служит, подобно синаптическим потенциалам мышц, сетчатки и т. д., своего рода генераторным потенциалом.
Токи, обнаруживаемые в центральных проводниках, уже не похожи на токи улитки. С удалением от кортиева органа частота их уменьшается и время возникновения все больше и больше запаздывает.
В подкорковых ядрах звукового анализатора формируется в грубой форме восприятие звука — животное, лишенное коры, реагирует лишь на звуки большой интенсивности. И только в корковом ядре (или конце) звукового анализатора возникает звуковое ощущение, соответствующее сигнальному значению слышимого звука. Корковый отдел не только принимает и анализирует информацию, поступающую из внутреннего уха, но имеет и обратную, эфферентную связь с улиткой, через посредство которой осуществляется регулирующая, настраивающая роль коры в отношении улитки (Ресмуссен, Г. В. Гершуни).
Теория Гельмгольца. Теории функционирования улитки
В 1863 г. Гельмгольц обосновал свою резонансную теорию, исходя из предположения, что улитка при помощи явлений физического резонанса может разложить сложные звуки на простые тоны. Ввиду того что основная мембрана благодаря эластическим волокнам натянута в поперечном направлении и так как она имеет разную ширину у основания и верхушки улитки, Гельмгольц считал, что она представляет собой подходящее образование, которое разными участками резонирует на звуки разной высоты
Особенно много возражений против резонансной теории имеется со стороны физиков, и в настоящее время резонансная теория в старой трактовке должна быть оставлена. Новые наблюдения и теоретические соображения говорят против того, что в улитке при прохождении звука имеет место механический резонанс наподобие резонанса струн. Так как основная мембрана составляет одну цельную натянутую перепонку, любая деформация будет более или менее сильно сказываться на широкой полосе или даже на всей мембране, но с максимумом в определенном месте.
Согласно этой теории, деформация основной перепонки, вызванная толчком стремени, движется с определенной быстротой в виде передвигающейся волны деформации по основной мембране.
Различие во взглядах отдельных авторов состоит лишь в том, что одни считают, что волна деформации быстро затухает, пройдя известное пространство по мембране, другие же считают, что бегущая волна проходит по всей длине перепонки, и третьи, наконец, допускают, что путем отражения образуются стоячие волны наподобие фигур Хладни (теория Эвальда).
Применяя современные достижения акустики, Бекеши (1928) изучил на модельных опытах, а также путем наблюдений за основной мембраной у морской свинки, произведенных при помощи оптических приборов и микроманипуляторов, характер изменений, которым подвергается основная мембрана при звуковом раздражении.
На весьма совершенных моделях, подбирая правильную упругость и толщину специальной резиновой пленки, он мог показать, что картины, полученные Эвальдом, являлись артефактом. В его опытах на мембране под влиянием звука возникала бегущая волна с быстро затухающей амплитудой. На месте максимальной амплитуды наблюдались вихри, скорость вращения которых пропорциональна величине амплитуды.
При сильных звуках получались точечные прорывы мембран, которые вызывались действием двух вихрей по обе стороны мембраны. Локализация этих перфораций зависела от частоты колебаний: чем выше звук, тем ближе к основанию образуется отверстие.
На препаратах улитки морской свинки Бекеши наблюдал, что экскурсии основной мембраны имеют более широкую зону при низких звуках и колебание перепонки заметно только у верхушки улитки. При помощи микроманипулятора он смог даже измерить величину прогиба основной мембраны.
Из современных теорий особенного внимания заслуживает теория Флетчера—Роафа, так как она расширяет наши понятия о резонансе—общепринятом в физике явлении, которое наилучшим образом объясняет анализ сложпого звука на его оставные. Эти авторы воспользовались данными Луца (Lutz), который показал, что в резонансе могут участвовать не только струны и мембраны, но и столбики жидкостей. Луц наполнял U-образные трубки разными порциями воды и вызывал колебания жидкости при помощи вибраторов. Оказалось, что интенсивность колебания столба жидкости зависит от частоты колебания вибратора.
При медленных вибрациях наибольший размах колебаний столба жидкости наблюдался в трубках, содержащих большое количество воды, наоборот, при частых колебаниях наиболее энергичные соколебания совершала жидкость небольшого объема.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
В 1863 г. Гельмгольц предложил резонансную теорию. Он считал, что в улитке происходят явления механического резонанса, в результате которого сложные звуки в ней разлагаются на простые тоны. То обстоятельство, что основная мембрана с ее эластическими волокнами, натянутыми в поперечном направлении, имеет разную ширину у основания и верхушки улитки (у основания — узкая, у верхушки — широкая), позволило Гельмгольцу считать ее образованием, разные участки которого способны резонировать на звуки неодинаковой высоты.
Теория Гельмгольца с гениальной простотой разъяснила основные свойства уха, т. е. определение высоты, силы и тембра. Согласно резонансной теории, любой чистый тон имеет свой ограниченный участок на основной мембране.
Одиночный звук, по его мнению, раздражает строго определенные нервные волокна — именно те, которые снабжают соответствующий участок мембраны, и раздражение этих волокон ощущается как звук строго определенной высоты.
Резонансной теорией легко объясняется различение тембра звука и способность уха разлагать сложный звук на его составные части. Каждый сложный звук, согласно этой теории, раздражает столько точек на основной мембране, сколько в нем содержится синусоид, т. е. основной тон и все обертоны вызывают соколебание в соответствующих участках мембраны. Все сигналы из внутреннего уха, дошедшие по системе проводников до слуховых центров, интегрируются в них, и мы слышим соответственный тембровый звук. Сила звука определяется числом раздражаемых нервных элементов. Естественно, чем сильнее звук, тем более широкий участок основной мембраны приходит в соколебание. Гельмгольц допускал резонанс отдельных участков мембраны, но не говорил о резонансе свободно колеблющихся струн. Таким образом, из теории Гельмгольца следует три основных вывода:
1) в улитке происходит первичный анализ звуков;
2) каждый простой звук имеет свой участок на основной мембране;
3) низкие звуки вызывают колебание участков основной мембраны у верхушки улитки, а высокие — у основания ее.
Несмотря на огромное количество новых фактов, полученных при изучении функции внутреннего уха, эти три вывода сохраняют свое значение до настоящего времени.
93. Анатомия, физиология полукружных каналов и преддверия лабиринта.
Перепончатые полукружные каналы расположены внутри костных и открываются 3 ампулярными и 2 простыми концами, т. е. 5 отверстиями, в utriculus. На поперечном разрезе полукружного канала видно, что перепончатый канал прилежит к одной из костных стенок. В перилимфатическом пространстве имеются поддерживающие соединительнотканные перемычки, по которым идут сосуды.
Более сложно устроены ампулярные концы перепончатого лабиринта, почти целиком заполняющие костные расширения. В них находятся ампулярные гребешки (cristae ampullares), на которых размещены опорные и нейроэпителиальные волосковые клетки (рецепторный аппарат). Особенностью строения последних является большая длина волосков, склеенных между собой эластической массой и образующих так называемую купулу (cupula). Это образование возвышается почти до полного соприкосновения с противоположной стенкой ампулы. Малейшее перемещение эндолимфы в канале неминуемо влечет за собой отклонение купулы, которая затем медленно возвращается в исходное положение.
К ампулярному гребешку подходят ампулярные веточки вестибулярного нерва.
В углублениях преддверия помещаются два перепончатых мешочка — эллиптический и круглый, носящие название utriculus и sacculus. Утрикулюс расположен в верхне-задней части преддверия, а саккулюс — внизу, вблизи основного завитка улитки. Оба мешочка содержат рецепторный аппарат, расположенный на статическом пятнышке — macula utriculi et sacculi seu macula statica. Оно состоит из опорных и нейроэпителиальных волосковых клеток, короткие волоски которых заключены в желатинообразную массу; на ней покоится отолитовая мембрана с входящими в ее состав кристаллами солей кальция.
Оба мешочка соединены между собой посредством тонкого канала — ductus utriculo-saccularis, который продолжается в эндолимфатический проток (ductus endolymphaticus). Последний помещается в костном канале — водопроводе преддверия (canaliculus vestibuli, seu aquaeductus vestibuli) и заканчивается расширением — эндолимфатическим мешком, лежащим в дупликатуре твердой мозговой оболочки на задней поверхности пирамидки.
Читайте также: