
Особенности строения коры кратко
Обновлено: 05.07.2024
Все стебли древесных растений, произрастающие в умеренных широтах, характеризуются определенным строением. Так из чего же состоит стебель? Стебель состоит из:
- древесина. Это центральная часть стебля;
- камбий. Это тонкий слой образовательной ткани;
- кора. Она находится снаружи.
Остановимся на строении стебля подробнее и рассмотрим элементы стебля: кору, древесину и сердцевину.
Древесина
Почти весь объем древесины представляет собой отмершие клетки. В основном — сосуды и трахеи, выполняющие проводящую функцию, а также склеренхимные клетки (то есть механические).
Древесина (ксилема) является основной частью стебля. Она включает сосуды (трахеи), трахеиды, древесные волокна (механическая ткань). Одно кольцо древесины образуется в течение года. Годичные кольца древесины служат для определения возраста растения.
Но в случае с тропическими растениями, сделать это довольно сложно: тропические растения растут постоянно на протяжении года, поэтому их кольца почти незаметны.
Годичные кольца хорошо видны весной, с пробуждением растения, и осенью, когда растение засыпает на зиму. Весенняя древесина включает тонкостенные клетки, а осенняя — толстостенные. Это говорит о том, что переход от весны к осени постепенный, а от осени к весне — внезапный.
Древесина также состоит из паренхимных клеток, которые в основном концентрируются в центральной части и образуют сердцевину.
Сердцевина — центральная часть стебля.
Внешний слой сердцевины включает живые паренхимные клетки и служит местом откладывания питательных веществ, а центральный слой — из больших и часто отмерших клеток.
Клетки сердцевины характеризуются наличием между ними межклеточного пространства. Также стоит отметить сердцевидный луч.
Сердцевидный луч — это ряд паренхимных клеток, которые берут начало в сердцевине и двигаются к первичной коре: они направлены радиально через древесину и луб. Луч выполняет важные функции: запасающую и проводящую.
Различают первичную и вторичную кору, так как у коры есть два отдела: луб и пробка.
Первичная кора — это участок стебля, состоящий из двух слоев: колленхимы или механической ткани, которая находится под перидермой, и паренхимы первичной коры, выполняющей запасающую функцию.
Первичная кора с типом ткани покрывным выполняет свою функцию недолго — сразу же за ней образуется вторичная покровная ткань — перидерма. Она включает 3 слоя клеток:
- пробку (внешний слой);
- пробковый камбий (средний слой);
- феллодерму (внутренний слой).
Пробка находится снаружи и возникает в результате многоразового заложения слоев перидермы. Таким образом она выполняет защитную функцию. На поверхности пробки можно обнаружить трещины. Их появление — результат утолщения стебля: пробковые клетки мертвые и не могут растягиваться.
Вторичная кора — это луб или флоэма. Луб включает в себя ситоподобные элементы, паренхимные клетки и лубовые волокна. Он прилегает к камбию.
Лубовые волокна — механическая ткань, поэтому они выполняют опорную функцию. Они образуют слой, получивший название твердый луб. Другие элементы луба образуют мягкий луб.
Появление клеток луба — результат деления и дифференциации камбия.
Камбий
Какую роль играет камбий? Камбий представляет собой образовательную ткань. Снаружи вторичную кору образуют клетки луба, а внутри — клетки древесины.
Стебель растет в толщину за счет деления клеток камбия. Зимой клетки камбия не делятся — их деление возобновляется весной.
Проводящие элементы древесины (ксилемы) обеспечивают перемещение воды и растворенных в ней веществ от корней до листьев.
Проводящие элементы луба (флоэмы) перемещают продукты ассимиляции от листков до корней.
Распределение флоэмы и ксилемы при образовании проводящих пучков осуществляется в определенном порядке и с учетом расположения других структур стебля. Ксилема входит в состав древесины и располагается в середине от камбия. Флоэма — составляющая луба: она располагается снаружи от камбия.
Мы рассмотрели внутреннее строение стеблей.
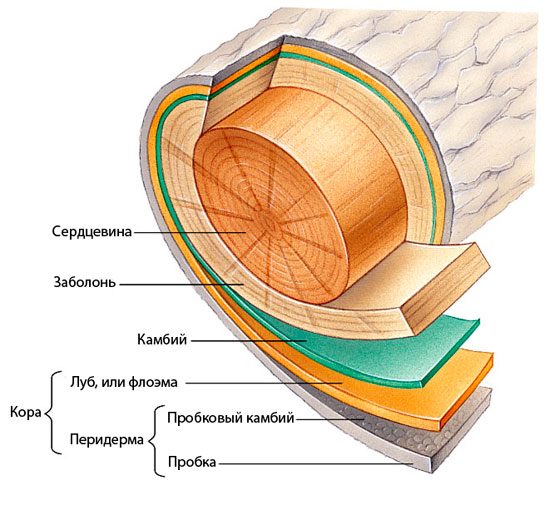
Первичное и вторичное анатомическое строение стебля
Анатомия стебля
Первичное строение стебля характеризуется наличием центрального цилиндра и первичной корки, между которыми граница определяется довольно условно.
Первичная кора состоит из следующих тканей:
- механические;
- ассимиляционные;
- запасающие;
- выделительные;
- воздухоносные.
Участки паренхимы разделяют проводящие пучки, которые собраны из первичных проводящих тканей. Расположение первичной флоэмы — периферия пучка. Первичная ксилема направлена к середине стебля. Расположение сердцевины — центральная часть.
Возникновение первичного камбия происходит вначале в первичных пучках. Поэтому между прослойками пучкового камбия появляются перемычки межпучкового камбия. Проводящие пучки легко различимы, так как пучковый камбий закладывает проводящие элементы, а межпучковый — паренхиму.
Строение стеблей древесных растений отдельных видов отличаются не пучковым типом вторичного утолщения.
Проводящие пучки сближаются и образуют 3 концентрических слоя:
- Древесину или вторичную ксилему.
- Камбий.
- Луб или вторичную флоэму.
Центральная часть — это сердцевина, которую составляют живые тонкостенные паренхимные клетки. Функция таких клеток заключается в накапливании питательных веществ.
Древесина занимает около 90% объема ствола и располагается снаружи от сердцевины.
Отдельно нужно отметить механические древесные волокна, обеспечивающие стволу прочность.
Древесина также включает паренхимные клетки (они образуют сердцевинные лучи) и клетки вертикальной паренхимы. Камбий находится между корой и древесиной и состоит из образовательной ткани. Последняя, в свою очередь, образует ксилему и флоэму.
Снаружи от камбия — вторичная кора или луб: ее образует камбий.
В луб входят ситовидные трубки, лубяные волокна и лубяная паренхима. Луб также может выполнять функцию накапливания питательных веществ. Около луба располагается запасающая паренхима, а за ней — перидерма или вторичная покровная ткань. Слой перидермы, который выполняет функцию защиты — это пробка. Пробка трансформируется в корку (третичную покровную ткань) спустя несколько лет.
Как минеральные вещества передвигаются по стеблю
Корни всасывают воду и минеральные соли из почвы, и они перемещаются по стеблю к листьям, цветам и плодам. Такое движение называют восходящим током. Оно осуществляется по древесине с помощью основных проводящих сосудов. Эти сосуды — мертвые пустые трубки, которые образуются из живых паренхимных клеток. Восходящий ток может осуществляться трахеидами: мертвыми клетками, которые связаны друг с другом при помощи окаймленных пор.
Образование органических веществ происходит в листьях. Затем они доставляются во всех органы растения, в том числе корень и стебель.
Нисходящий ток — это обратная транспортировка. В ней принимают участи луб (по нему перемещаются) и ситовидные трубки (с их помощью). Ситовидные трубки — это живые клетки, которые связаны между собой ситечками: тонкими перегородками с отверстиями. Эти трубки находятся в продольных и поперечных стенках. При помощи сердцевидных лучей питательные вещества у древесных растений перемещаются в горизонтальной плоскости.
Как органические вещества откладываются в стеблях
Внутреннее строение стебля создано для откладывания питательных веществ. Органические вещества внутри клеток или в оболочках клеток накапливаются в специальных запасающих тканях. Эти ткани образуются из паренхимных клеток. Среди таких органических веществ — крахмал, аминокислоты, инсулин, сахара, масла, белки.
Органические вещества откладываются в стебле в разных местах: в паренхимных клетках первичной коры, в живых клетках сердцевины, в сердцевидных лучах.
Запасающие ткани играют важную роль в питании растения органическими веществами. Запас органических веществ растениями — это еще и продукт питания для животных и человека. Питательные вещества растений используются людьми в качестве сырья.
В зависимости от экологических условий у большинства растений меняется характер элементов, из которых они состоят. Вместе с тем перераспределяются и ткани, большинство из которых проходит через все органы растения непрерывно. Однако видоизменяются они в разных частях в соответствии с их функциями.
В начальный период развития в стебле древесного и травянистого двудольного растения чаще всего выделяют первичную кору, центральный цилиндр и сердцевину.
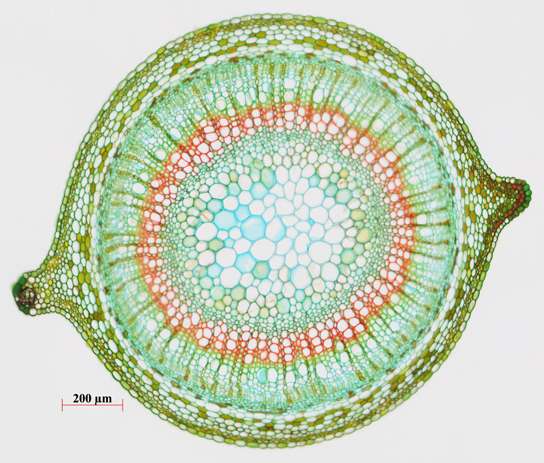
Стебель
Вам будет интересно: Куранты - это символ России. Описание главных часов страны
Первичная кора стебля является его внешней частью. Покрыта она эпидермисом и простирается до центрального цилиндра. Включает в себя основную паренхиму, ассимиляционную, механическую, выделительную, запасающую, секреторную и другие ткани. Преимущественно формируется многослойной туникой конуса нарастания. Первичная кора при переходе к строению стебля вторичного типа деформируется и по результатам деятельности феллогена отторгается в корковый слой.
Особенности строения первичной коры
Между двумя соседствующими тканями: эпидермисом и эндодермой, заключена эта кора. Для разных групп растений неодинаковы цитологические свойства этой части стебля.
Первичная кора в своем составе помимо двух прилегающих друг к другу тканей имеет:
- субэпидермальный слой — гиподерму, который большей частью состоит из живых клеток с зелеными пластидами;
- механические ткани, наиболее распространенной из которых является колленхима (встречаются также волокна и склереиды);
- основную паренхиму.
Функции
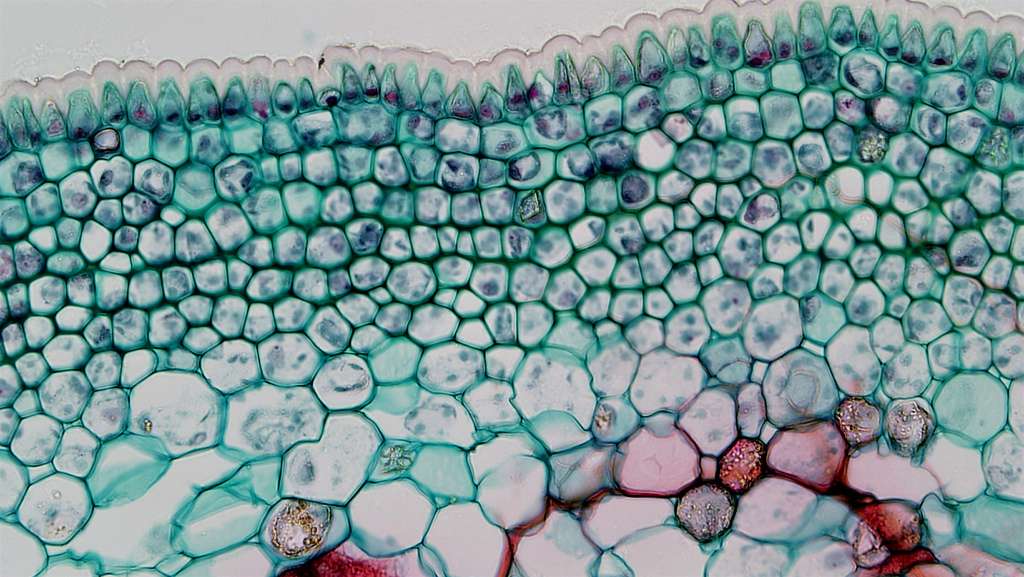
Первичная кора выполняет следующие функции:
- защищает стелу;
- способствует выборочному поглощению веществ из почвы и их транспортировке в стелу;
- содействует в загрузке ксилемы;
- является хранителем запасов воды (корневые шишки аспарагуса);
- в ней также развиваются гифы грибов, образующие микоризу.
Эндодерма
Во всех органах растения присутствует эндодерма как внутренний слой коры. Наиболее дифференцирована она в корнях и представлена в стебле преимущественно однорядным, узким слоем клеток, которые расположены очень компактно.
На первых этапах развития эндодерма дифференцируется в онтогенезе растения и с клетками коры имеет общее начало, поэтому справедливым будет ее назвать самым глубоким слоем коры.
Стадии эндодермы
Меристематическая фаза эндодермы называется проэндодермой, или эмбриональной эндодермой. О типичной эндодерме можно говорить только после того, как утолщенная полоса другого химического состава возникает на мельчайших целлюлозных стенках ее клеток. На поперечном сечении эта полоса хорошо заметна. Она опоясывает поперечные и радиальные стенки клеток. Полоса носит название Каспари в честь ученого, впервые подробно описавшего ее. Первой стадией развития эндодермы является клетка с такой полосой.
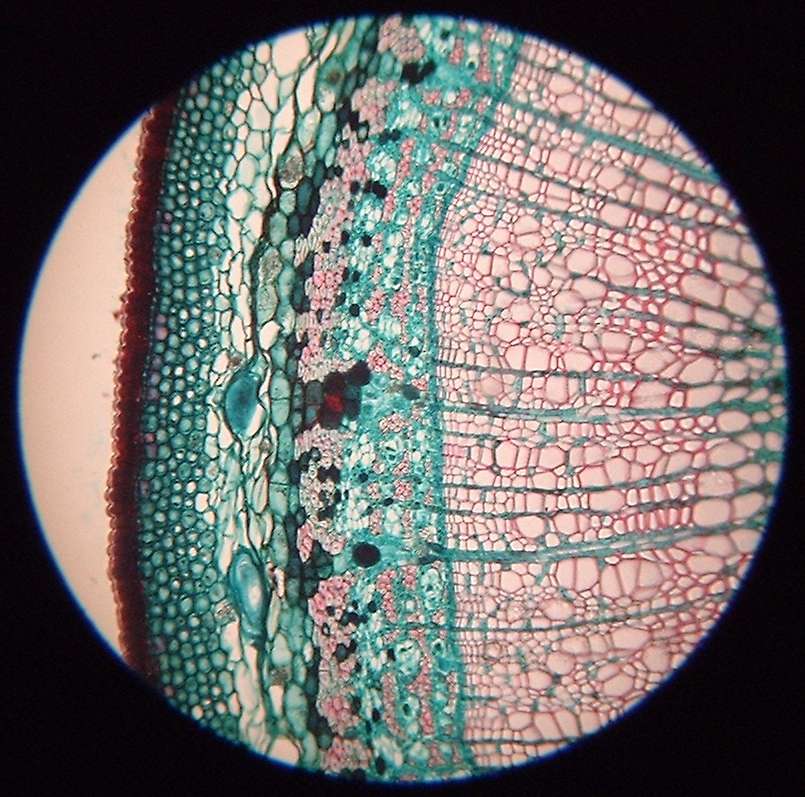
Вторая стадия обусловлена появлением на стенках клеток субериновой пластинки, которая равномерно образуется по всей стенке. Полностью не объяснен механизм формирования суберина, однако известно, что причиной его возникновения является окисление и конденсация фенолов и ненасыщенных жирных кислот при содействии ферментативной системы.
Многочисленные слои целлюлозы постепенно накладываются на вторичную стенку на третьей стадии эндодермы. В большинстве случаев эти слои видны в микроскоп без предварительной обработки. Они лигнифицированы и могут содержать в себе всевозможные включения.
У каких растений присутствует эндодерма?
Эндодерма широко распространена среди разнообразных групп растений. Только у псилофитов (самых низших форм ископаемых, не имеющих листьев) она отсутствует. У птеридофитов эндодерма на первой и второй стадиях, за некоторыми исключениями, находится в корне, черешках вайи, стебле и листочках перистого листа, то есть она проходит через все тело растения. Эндодерма также находится в корнях голосеменных растений, там она быстро пересекает первую стадию и переходит во вторую, но третьей никогда не достигает. Также она не встречается в стеблях голосеменных, только более или менее глубоко заходит в гипокотиль у хвойных.
Очень правильное строение имеет эндодерма в корнях покрытосеменных. В зависимости от вида растения, первая, вторая или третья стадия могут сохраняться на большой длине корня. Для стеблевых органов и корней водных растений характерно длительное продолжение первой стадии эндодермы.

Как правило, типичная эндодерма отсутствует в надземных органах покрытосеменных. Однако отличительной чертой внутреннего слоя коры от остальных клеток является то, что он содержит в себе в большом количестве крупные крахмальные зерна. Данный слой считают гомологом эндодермы, так как он занимает ее место.
Более старые участки занимает обычная коровая паренхима, но бывает и так, что крахмалоносное влагалище, так еще называют внутренний слой первичной коры, разграничивается как типичная эндодерма, имеющая полосы Каспари.
Перидерма
Первичная кора у древесных растений недолговечна. Перидерма (вторичная покрывающая ткань) закладывается в различных слоях коры у разных растений на ветках первого года жизни. Все ткани, которые оказались снаружи перидермы, вскоре отмирают, так как они изолированы от центрального цилиндра и живых тканей коры. В связи с тем, что феллоген способствует отложению пробковой ткани, объем тканей первичной коры будет постепенно сокращаться. Когда произойдет заложение феллогена, она будет отодвинута наружу слоями пробки в эндодерму или в перициклу, где вскоре засохнет.
В это же время в центральном цилиндре происходят значительные изменения, благодаря деятельности камбия.
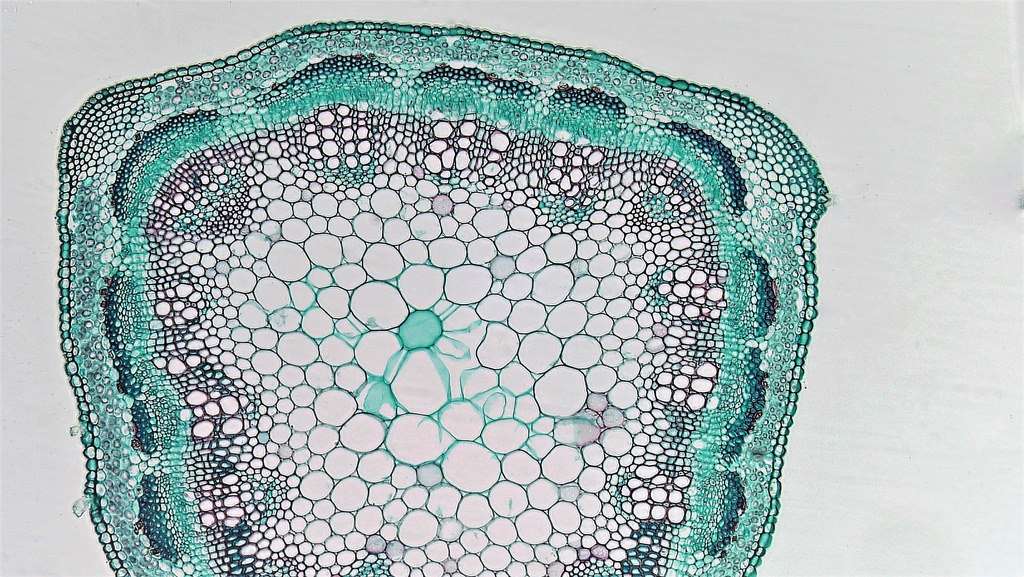
Обычно вторичную кору, древесину и сердцевину различают во вторичной структуре стебля.
Такие понятия, как первичная и вторичная кора, не являются гомологичными. Последняя отличается от первой составом, функцией и происхождением и представляет собой совокупность тканей, которые лежат снаружи от камбия, включая твердый и мягкий луб.
Если же сохраняются остатки первичной коры, то их называют вторичными покровными тканями. Именно таким образом различные по функциональному значению и происхождению ткани попадают во вторичную кору.
Строение коры головного мозга: слои, клетки
Толщина коры больших полушарий (от греч. pallium — мантия) варьирует от 2 до 4 мм, наиболее тонкая ее часть находится в области первичной зрительной коры, а наиболее утолщенная — в первичной моторной области. Больше половины общей поверхности коры скрыто стенками борозд. Мозг содержит приблизительно 86 млрд, нейронов (кора больших полушарий содержит только 19% общего количества, но составляет 81% массы мозга), аналогичное число глиальных клеток и разветвленную капиллярную сеть.
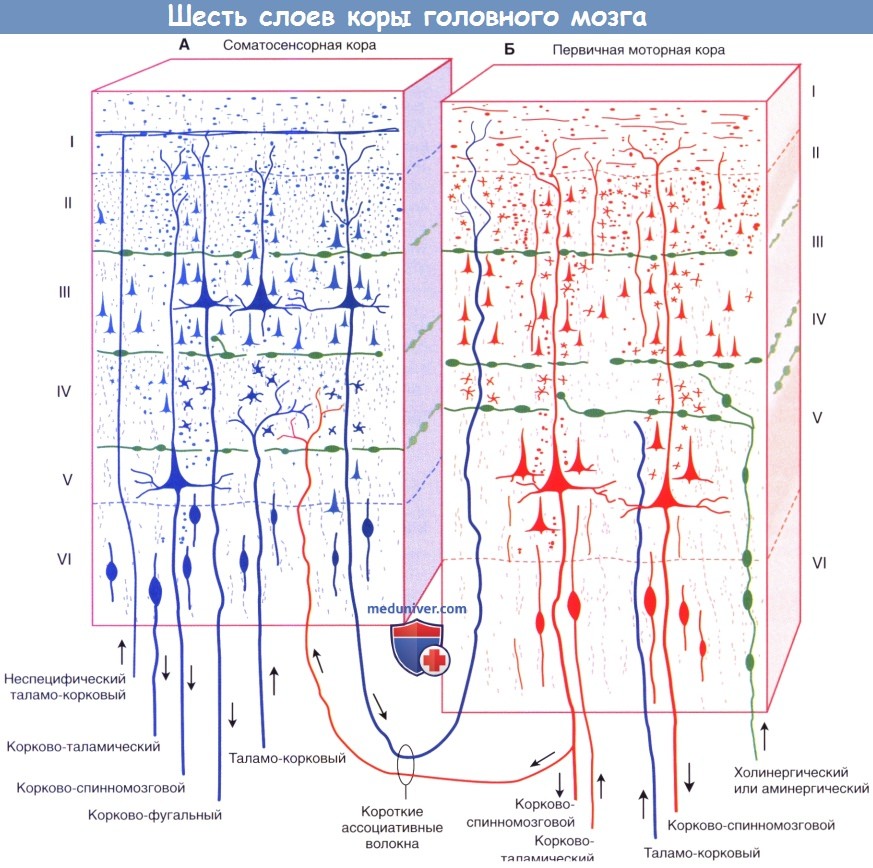
Шестислойное строение изокортекса коры головного мозга.
(А) Соматосенсорная кора. (Б) Первичная моторная кора, кортикальные пластинки обозначены цифрами от I до VI.
Клеточные пластинки новой коры (неокортекса):
I. Молекулярный слой образован концами апикальных дендритов пирамидных клеток и наиболее дистальными ветвями аксонов, проходящих к коре от интраламинарных ядер таламуса.
II. Наружный зернистый слой образован мелкими пирамидными и звездчатыми клетками.
III. Наружный пирамидный слой состоит из средних пирамидных клеток и звездчатых клеток.
IV. Внутренний зернистый слой образован звездчатыми клетками, к которым подходят восходящие волокна от релейных таламических ядер. Звездчатые (зернистые) клетки особенно многочисленны в первичной соматосенсорной коре, первичной зрительной коре и первичной слуховой коре; к ним подходят афферентные чувствительные волокна. Эти отделы обозначают как гранулярная кора. Наоборот, первичная моторная кора — место начала корково-спинномозгового и корково-бульбарного путей, имеет относительно небольшое количество зернистых клеток в слое IV и большое число пирамидных клеток в слоях III и V, которые смазывают (перекрывают) отдельные слои. Эту область обозначают как агранулярная кора.
V. Внутренний пирамидный слой образован крупными пирамидными клетками, волокна которых направляются к полосатому телу, стволу мозга и спинному мозгу.
VI. Слой веретеновидных клеток образован видоизмененными пирамидными клетками, волокна которых направляются к таламусу.
Взаимодействие между группами колонн позволяет осуществлять более сложную деятельность, модели поведения или решать когнитивные задачи.
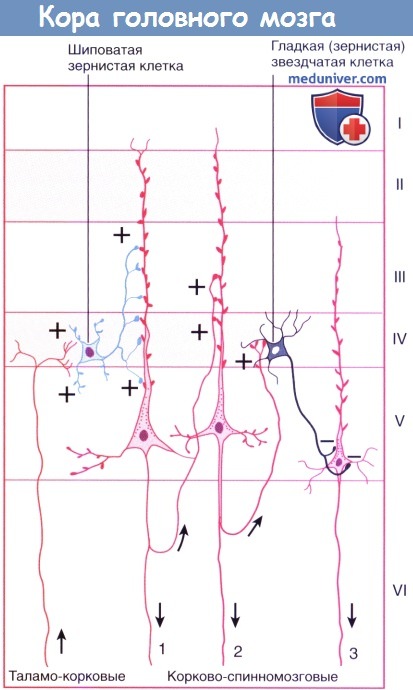
Восходящие/нисходящие проводящие пути. Стрелки указывают направление проведения импульса. Знаки +/- обозначают возбуждение/торможение.
Пирамидная клетка 1 возбуждается шиповатой зернистой клеткой; она возбуждает клетку 2 внутри своей колонны клеток; клетка 3 в соседней колонне тормозится гладкой зернистой клеткой.
в) Типы клеток. Морфологически кортикальные нейроны разделяют на две большие группы. Большая часть (60-85%) представлена пирамидными нейронами (из-за их формы) — единственными, волокна которых покидают кору (и к которым подходит большая часть волокон), что объясняет их альтернативное название — корковые проекционные нейроны; их волокна — возбуждающие глутаматергические. К оставшимся 15-40% нейронам относят непирамидные, или вставочные, нейроны; несмотря на то, их взаимосвязи не выходят за пределы коры, они осуществляют регуляцию и значительное влияние на ее деятельность; их тип — преимущественно тормозной ГАМК-ергический.
Внутри каждой группы можно выделить множество подгрупп в зависимости от морфологии, взаимосвязей, электрофизиологических свойств, типа развития, физиологических характеристик, молекулярных маркеров и т. д. (Примерами основных морфологических и функциональных клеточных типов служат пирамидные клетки, шиповатые зернистые клетки (измененные пирамидные клетки) и группа непирамидных тормозных вставочных нейронов.)
• Шиповатые зернистые клетки — один из вариантов атипичных пирамидных клеток, лежащих в слое IV и наиболее многочисленных в первичной сенсорной коре. Их шиповатые дендриты не выходят за пределы слоя IV, а аксоны могут подниматься или опускаться, образуя возбуждающие глутаматергические синаптические контакты с пирамидными клетками. На них переключается большая часть волокон восходящих таламических путей к слою IV с дальнейшим радиальным распространением.
Для организации их можно разделить на три крупных семейства в зависимости от экспрессируемых этими вставочными нейронами биомаркеров: парвальбумин, соматостатин и серотонин (5-гидрокситриптамин, 5НТ) 3а-рецептор (5HT3aR).
• Парвальбумин-экспрессирующие вставочные нейроны не имеют шигшков на дендритах. К ним подходят возбуждающие волокна от таламуса и коры, а тормозные — от других вставочных нейронов того же типа. Считают, что они играют роль в стабилизации активности корковых нейронных сетей. Как и в коре мозжечка, эти нейроны обладают фокусирующим действием на кору больших полушарий, тормозя слабо возбужденные колонны клеток. Клетки-канделябры (названы так из-за канделяброподобных групп аксоаксональных синаптических окончаний) наиболее многочисленны в слое II, образуют контакты с начальным сегментом аксона пирамидной клетки и играют ключевую роль в корково-корковых взаимодействиях.
• Соматостатин-экспрессирующие вставочные нейроны. Примером служат клетки Мартинотти, расположенные в пластинках V и VI, аксоны которых направляются к пластинке I. Получая сигналы от пирамидных клеток, они способны ограничивать их периферическое возбуждение и объединять несенсорную информацию, осуществляя, таким образом, регулирование обработки сигналов в дендритах их пирамидных клеток в зависимости от внешних обстоятельств.
• 5НТ3а-экспрессирующие вставочные нейроны — гетерогенная группа, к которой относят большую часть вставочных нейронов поверхностных корковых слоев. Получая информацию от корковых и таламических нейронов, они могут участвовать в процессах обучения, осуществляя влияние на корковые нейронные цепи. Он нейроглиеформных клеток (паутинообразных клеток)—одного из главных типов вставочных нейронов слоев II и III — веерообразно расходятся дендриты, обладающие уникальным свойством образования синапсов друг с другом и другими типами вставочных нейронов; этот факт указывает на их важнейшую роль в синхронизации корковых нейронных цепей. Другая морфологически гетерогенная группа вставочных нейронов, помимо ГАМК, экспрессирует вазоактивный интестинальный полипептид; другие типы вставочных нейронов этой группы экспрессируют также холецистокинин и другие пептидные рецепторы.
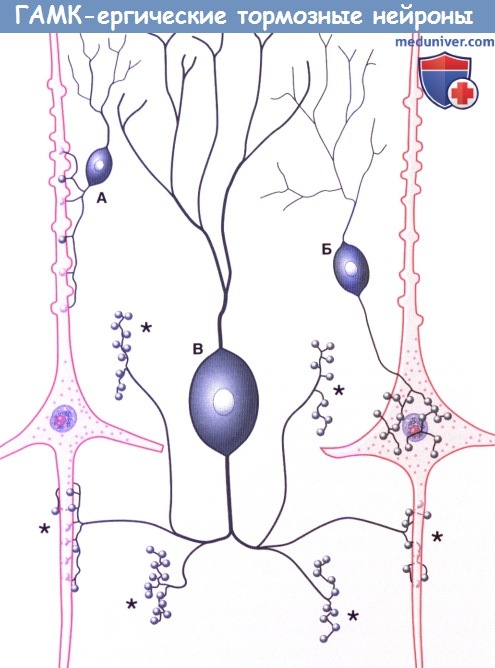
Три морфологических типа ГАМК-ергических тормозных нейронов:
А. Аксодендритическая клетка, образующая синапс с апикальным отделом пирамидного нейрона.
Б. Корзинчатая клетка, образующая аксоаксональные синапсы с пирамидными клетками.
В. Клетка-канделябр, формирующая аксоаксональные синапсы (*) с начальными сегментами аксонов двух пирамидных клеток, показанных здесь, и с начальными сегментами четырех других клеток, не показанных на рисунке.
(На основе DeFelipe, 1999, с разрешения автора.)
г) Афферентные волокна. Восходящие волокна к любому отделу коры могут иметь четыре различных источника (в первую очередь, корковые) и оканчиваются в разных отделах.
1. Длинные и короткие ассоциативные волокна от мелких и средних пирамидных клеток в слоях II и III из других корковых зон того же полушария.
2. Комиссуральные волокна от средних пирамидных клеток слоев II и III проходят через мозолистое тело от аналогичных или топографически идентичных участков противоположного полушария.
3. Таламо-корковые волокна от соответствующего специфического или ассоциативного ядра (например, волокна от вентрального заднего таламического ядра к соматосенсорной коре) и от дорсомедиального таламического ядра к предлобной (префронтальной) коре (описана ниже) оканчиваются в пластинке IV. Неспецифические таламо-корковые волокна от внутрипластинчатых ядер оканчиваются во всех пластинках.
4. Холинернические и аминергические волокна от базальных ядер, гипоталамуса и ствола мозга. Эти волокна обозначены на рисунке ниже зеленым цветом. Несмотря на то, что они распространены по всей коре, их возбуждение не сопровождается генерализованной или неспецифической реакцией. Анатомическая специфичность волокон (корковая, пластинчатая и клеточная) обусловливает возбуждение или торможение только определенных групп нейронов. Ядра их начала и нейромедиаторы:
• базальное ядро Мейнерта (базальные ядра мозга), ацетилхолин;
• серобугорно-сосцевидное ядро (задний отдел гипоталамуса), гистамин;
• черная субстанция, компактный слой (вентральный отдел покрышки среднего мозга), дофамин;
• ядра шва (средний мозг и ростральный отдел моста), серотонин;
• голубое пятно (ростральный отдел моста), норадреналин. Эти пять групп нейронов имеют особое значение в психиатрии и подробно рассмотрены в отдельной статье на сайте.
д) Эфферентные волокна. Единственные эфферентные волокна, покидающие кору больших полушарий, представлены возбуждающими аксонами пирамидных клеток. Одна часть аксонов пирамидных клеток соединяется с длинными и короткими ассоциативными волокнами, другая образует комиссуральные или проекционные пути. Ассоциативные и комиссуральные пути составляют большой объем белого вещества полушарий головного мозга.
• Примерами коротких ассоциативных путей (проходят между соседними зонами коры в составе поверхностного белого вещества в виде U-волокон) служат пути, направляющиеся в моторную кору от сенсорной коры и обратно. Примерами длинных ассоциативных путей служат волокна между префронтальной корой (лежащей кпереди от моторных зон коры) и ассоциативными сенсорными зонами. Источник этих волокон — пирамидные клетки, расположенные преимущественно в слоях II и III.
• Комиссуральные волокна мозга представлены исключительно аксонами пирамидных клеток, проходящих через мозолистое тело, переднюю и заднюю спайки (и другие мелкие спайки) к аналогичным участкам противоположного полушария (например, волокна от первичной корковой зоны направляются к соответствующей ассоциативной области другого полушария), а также к несоответствующим зонам [подобные комиссуральные связи отсутствуют между первичной зрительной корой, первичной соматосенсорной и моторной корой, иннервирующими дистальный отдел верхней конечности]. Волокна начинаются от пирамидных клеток, расположенных преимущественно в слоях II и III.
• Основная часть проекционных волокон от первичной сенсорной и моторной коры подходит к базальным ганглиям. К таламусу направляются волокна от всех отделов коры. К другим основным проводящим путям относят корково-мостовой (к ипсилатеральным ядрам моста), корково-ядерный (к двигательным и чувствительным ядрам черепных нервов моста и продолговатого мозга на противоположной стороне) и корково-спинномозговой. Источником этих волокон служат пирамидные клетки, расположенные преимущественно в слоях V и VI (отдающие волокна преимущественно к специфическим релейным ядрам таламуса).
Древесный ствол имеет сложную структуру, которая включает следующие элементы (рис. 1):
- древесина:
- сердцевина;
- внутренняя (ядровая) часть ствола;
- внешние слои древесины (заболонь);
- камбий;
- внутренний слой коры (луб);
- корка (слой отмерших клеток, пробковый слой или вторичная кора, перидерма).
![строение древесного ствола и коры]()
Рис. 1. Анатомическое строение древесного ствола и коры: а – фрагмент среза слоев коры; б – поперечный срез ствола; 1– заболонь (наружный слой древесины); 1 – камбий; 3 – луб; 4 – корка; 5 – сердцевина; 6 – ядро
1. Древесина
1.1. Сердцевина
В раннем возрасте древесина всех деревьев состоит только из заболони. Через некоторое время, по мере созревания, происходит отмирание центральной части и образуется сердцевина. Сердцевина – это небольшого диаметра центральная часть ствола отмершей древесины. Она малой прочности, состоит из рыхлой первичной ткани, которая легко загнивает.
1.2. Внутренняя часть ствола (ядро)
В древесине выделяется внутренняя часть ствола, или, как ее еще называют, ядро, состоящая из спелой древесины. В процессе созревания деревьев происходит отмирание спелой древесины. Этот процесс сопровождается потемнением центральной части ствола. У разных пород различная степень интенсивности потемнения. Интенсивное потемнение наблюдается у так называемых ядровых пород (рис. 2, а, в, г, д). К ядровым породам относятся, например, лиственница, сосна.
Породы с однородной окраской древесины называются безъядровыми (рис. 2, б). К ним относятся, например, ель, пихта.
![структуры пород деревьев]()
Рис. 2. Примеры структуры пород деревьев: а – ядровая порода (сосна); б – безъядровая порода; в – округлое двухзональное ядро (в центре сформировавшееся ядро, а вокруг него морозное ядро); г – звездчатое ядро; д – срез осины
Стенки клеток ядра у хвойных пород пропитаны смолою. Движения влаги по клеткам ядра нет, поэтому древесина ядровой части ствола обладает большей прочностью и стойкостью к загниванию, чем наружная часть древесины, называемая заболонью.
1.3. Внешние слои древесины (заболонь)
Ядровую часть окружает физиологически активная наружная светлоокрашенная зона древесины, примыкающая к образовательной ткани – камбию. Эта часть ствола называется заболонью. От внутренней части ядровой древесины она отличается более светлой окраской. По клеткам заболони перемещается влага с растворенными в ней питательными веществами, поэтому она в сравнении со спелой ядровой древесиной имеет более высокую влажность, меньшую механическую прочность, наименьшую стойкость к биологическому повреждению, поражениям грибами и насекомыми. В пиломатериалах вследствие значительной усушки заболонь усиливает коробление древесины.
2. Кора дерева
Кора выполняет следующие физиологические функции дерева в процессе его роста:
- проводит по стволу питательные вещества, выработанные листьями;
- защищает дерево от внешних неблагоприятных условий.
Кора дерева имеет сложную структуру и состоит из следующего комплекса тканей в виде слоев:
- внутренний слой, прилегающий к камбию (луб);
- наружный слой (корка).
У каждой зоны слоев специфическая структура и химический состав, каждая зона имеет свои биологические функции. Переход от одного слоя к другому может быть резким или плавным в зависимости от породы дерева.
Соотношение зон этих слоев разное у различных пород и даже у деревьев одной породы, произрастающих в разных местах. Доля корки в коре составляет от 20 до 80 %.
Чем больше диаметр ствола дерева, тем толще кора на его поверхности (табл. 1). Но с увеличением возраста дерева, а значит, и его диаметра относительный объем коры уменьшается, так как чем больше диаметр ствола, тем меньше доля коры в объеме ствола [2].
Таблица 1. Зависимость толщины коры от диаметра ствола [2]
Толщина коры уменьшается от комля к вершине ствола, и соответственно изменяется относительный объем коры по высоте ствола.
В пределах одной породы относительный объем коры тем больше, чем хуже условия произрастания дерева. Показатели объемов коры различных пород существенно различаются (табл. 2) [2].
Таблица 2. Содержание коры в объеме стволовой части дерева
Кора имеет сложную структуру, которая включает граничный слой между древесиной и корой – камбий, внутренний и наружный слои коры, а также физиологические граничные зоны.
2.1. Камбий
На границе между внутренним слоем коры и древесиной располагается граничный слой, который называется камбиальным слоем или камбием. В стволе дерева камбий состоит из живых клеток древесины. Благодаря тому, что эти клетки непрерывно делятся, происходит рост тканей древесины и луба. В результате такой жизнедеятельности увеличивается диаметра древесного ствола и толщина слоя коры. В структуре ствола в сравнении с клетками других частей дерева камбиальный слой имеет самую низкую механическую прочность, на чем и основывается принцип окорки лесоматериалов.
2.2. Внутренний слой коры (луб или флоэма)
В зоне от камбия до корки расположен лубяной слой. С внутренней стороны он переходит в камбиальную зону, которая формирует клетки луба и древесины, а с внешней стороны − в пробковый камбий.
Ткани луба делятся на три вида:
- ситовидные трубки с клетками;
- механические ткани (склереиды и волокна);
- лубяная паренхима с паренхимными клетками. Каждый вид ткани выполняет свои функции.
Луб обеспечивает движение по фильтрующим трубкам, из которых он состоит, вдоль ствола дерева ко всем органам питательных органических веществ (продуктов фотосинтеза), образующихся в листьях. Волокнистая структура луба ориентирована волокнами вдоль ствола дерева.
У некоторых хвойных пород, например у сосны, клетки луба с внутренней стороны мало отличаются по строению от клеток камбия, и они называются камбиформа. Наличие клеток камбиформа определяет постепенный переход от камбия к лубу. Переход луба с внешней стороны к коре более резкий.
Различие в механической прочности и соотношении связей клеток камбия с древесиной, камбиформа с лубом и луба с коркой используется для определения видов окорки с полной очисткой древесины от луба (чистой окорки) или оставлением этого слоя (грубой окорки).
В зоне перехода между лубомрикокйо расположен слой паренхимных клеток. При делении этих клеток с внутренней стороны откладываются клетки лубяной паренхимы, а с внешней стороны образуются пробковые клетки. Оболочки последних не имеют пор и пропитаны суберином, что делает их непроницаемыми для воды и воздуха. Каркасную структуру корки образуют лубяные волокна и каменистые клетки. В целом пробковые клетки выполняют защитную функцию. Кроме перечисленных особенностей, луб имеет еще ряд свойств.
Так, в отличие от клеток корки клетки тканей луба имеют повышенное содержание урановых кислот и пентозанов, суберин отсутствует, а оболочки состоят из целлюлозы и гемицеллюлозы, не содержат лигнина [1].
В лубе содержится больше экстрактивных веществ, урановых кислот и пентозанов, но меньше лигнина. Также содержится в 1,5 раза меньше целлюлозы, чем в древесине [1].
Объемное соотношение корки и луба вдоль ствола меняется. В комлевой части сосны, где значительно развита покровная ткань, корка составляет основную долю объема коры. У ели изменение по строению и объемному соотношению корки и луба по длине ствола незначительно.
2.3. Корка
В технической литературе корка может называться по-разному, например слой отмерших клеток, пробковый слой или вторичная кора, перидерма. Во всяком случае, под этими терминами понимается наружный омертвевший слой коры, который выполняет функцию защиты тканей луба и древесины от механических повреждений, проникновения грибов и бактерий, резких колебаний температуры, испарения влаги, внешних климатических воздействий. У различных пород древесины корка имеет разную структуру и прочность. У хвойных пород она рыхлая, непрочная, поэтому в процессе окорки легко разрушается инструментами и удаляется с поверхности ствола.
Корка лиственных пород, осины и особенно березы имеет большую прочность корковой наружной части и значительно утолщенный слой луба. Эти факторы усложняют процесс окорки. Кроме того, кора березы, или береста, имеет высокую прочность в продольном направлении, а разрывается и отделяется в основном только в поперечном направлении. Это еще больше затрудняет окорку и измельчение коры.
Читайте также:
- Четвертичный сектор экономики кратко
- Философские взгляды на семью руссо кратко
- Кризис европейской цивилизации и поиски выхода из него американский вариант кратко
- Расскажите о художественных объединениях 1920 о борьбе литературно художественных группировок кратко
- Способ байера получение глинозема кратко

