
Окисление жирных кислот биохимия кратко
Обновлено: 02.07.2024
Бета-окисление – это метаболический процесс, включающий несколько стадий, посредством которых молекулы жирных кислот расщепляются для производства энергии. Более конкретно, бета-окисление состоит в том, чтобы долго разрушаться жирные кислоты которые были превращены в ацил-СоА-цепи в постепенно уменьшающиеся жирные ацил-КоА-цепи. Эта реакция высвобождает ацетил-КоА, FADH2 и NADH, три из которых затем вступают в другой метаболический процесс, называемый циклом лимонной кислоты или Цикл Кребса, в котором производится АТФ для использования в качестве энергии. Бета-окисление продолжается до тех пор, пока не образуются две молекулы ацетил-КоА, и цепь ацил-КоА полностью не разрушается. В эукариотических клетках бета-окисление происходит в митохондрии в то время как в прокариотических клетках это происходит в цитозоль.
Для того, чтобы произошло бета-окисление, жирные кислоты должны сначала попасть в клетка сквозь клеточная мембрана, а затем привязать к коэнзим A (CoA), образуя жирный ацил CoA и, в случае эукариотических клеток, попадает в митохондрии, где происходит бета-окисление.
Где происходит бета-окисление?
Бета-окисление происходит в митохондриях эукариотических клеток и в цитозоле прокариотических клеток. Однако, прежде чем это произойдет, жирные кислоты должны сначала проникнуть в клетку, а в случае эукариотических клеток – в митохондрии. В случаях, когда цепочки жирных кислот слишком длинные, чтобы проникнуть в митохондрии, бета-окисление также может происходить в пероксисомах.
Во-первых, белковые транспортеры жирных кислот позволяют жирным кислотам проходить через клеточную мембрану и проникать в цитозоль, поскольку отрицательно заряженные цепи жирных кислот не могут пересекать его иначе. Затем фермент жирная ацил-КоА-синтаза (или FACS) добавляет группу СоА к цепи жирных кислот, превращая ее в ацил-КоА.
В зависимости от длины цепь ацил-КоА будет проникать в митохондрии одним из двух способов:
Как упомянуто выше, если цепь ацил-КоА слишком длинная, чтобы обрабатываться в митохондриях, она будет разрушена в результате бета-окисления в пероксисомах. Исследования показывают, что очень длинные цепочки ацил-КоА разрушаются до тех пор, пока они не станут длиной 8 атомов углерода, после чего они транспортируются и вступают в цикл бета-окисления в митохондриях. Бета-окисление в пероксисомах дает H2O2 вместо FADH2 и NADH, в результате чего выделяется тепло.
Бета-этапы окисления
Бета-окисление происходит в четыре этапа: дегидрирование, гидратация, окисление и тиолизис. Каждый шаг катализируется отдельным ферментом.
Вкратце, каждый цикл этого процесса начинается с цепи ацил-КоА и заканчивается одной ацетил-КоА, одним FADH2, одним NADH и водой, и цепь ацил-КоА становится на два атома углерода короче. Общий выход энергии за цикл составляет 17 молекул АТФ (подробности о распаде см. Ниже). Этот цикл повторяется до тех пор, пока не образуются две молекулы ацетил-КоА, в отличие от одного ацил-КоА и одной ацетил-КоА. Четыре стадии бета-окисления описаны ниже, и их можно увидеть в ссылках на рисунки в конце каждого объяснения.
дегидрогенизация

На первом этапе ацил-КоА окисляется ферментом ацил-КоА-дегидрогеназой. Двойная связь образуется между вторым и третьим атомами углерода (С2 и С3) цепи ацил-КоА, входящей в цикл бета-окисления; конечным продуктом этой реакции является транс-2-еноил-КоА (транс-дельта-2-еноил-КоА). На этом этапе используется FAD и вырабатывается FADH2, который входит в цикл лимонной кислоты и образует АТФ для использования в качестве энергии. (Обратите внимание, что на следующем рисунке отсчет углерода начинается с правой стороны: самый правый углерод ниже атома кислорода – это С1, затем С2 слева, образующий двойную связь с С3, и т. Д.)
гидратация

На втором этапе двойная связь между С2 и С3 транс-Δ2-еноил-КоА гидратируется с образованием конечного продукта L-β-гидроксиацил-КоА, который имеет гидроксильная группа (ОН) в С2, вместо двойной связи. Эта реакция катализируется другим ферментом: еноил-КоА-гидратазой. Этот шаг требует воды.
оксидирование
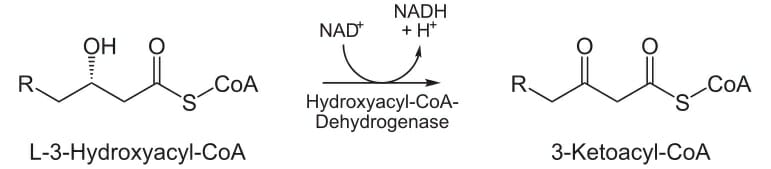
На третьей стадии гидроксильная группа в С2 L-β-гидроксиацил-КоА окисляется NAD + в реакции, которая катализируется 3-гидроксиацил-КоА-дегидрогеназой. Конечными продуктами являются β-кетоацил-КоА и NADH + H. NADH входит в цикл лимонной кислоты и производит АТФ, который будет использоваться в качестве энергии.
тиолиза

Наконец, на четвертом этапе β-кетоацил-КоА расщепляется тиольной группой (SH) другого КоА. молекула (КоА-SH). Фермент, который катализирует эту реакцию, является β-кетотиолазой. Расщепление происходит между С2 и С3; следовательно, конечные продукты представляют собой молекулу ацетил-КоА с исходными двумя первыми атомами углерода (С1 и С2) и цепью ацил-КоА на два атома углерода, которая короче, чем исходная цепь ацил-КоА, которая вступила в цикл бета-окисления.
Конец бета-окисления
В случае четных ацил-СоА-цепей бета-окисление заканчивается после того, как четырехуглеродная ацил-КоА-цепь распадается на две единицы ацетил-КоА, каждая из которых содержит два атома углерода. Молекулы ацетил-КоА вступают в цикл лимонной кислоты с образованием АТФ.
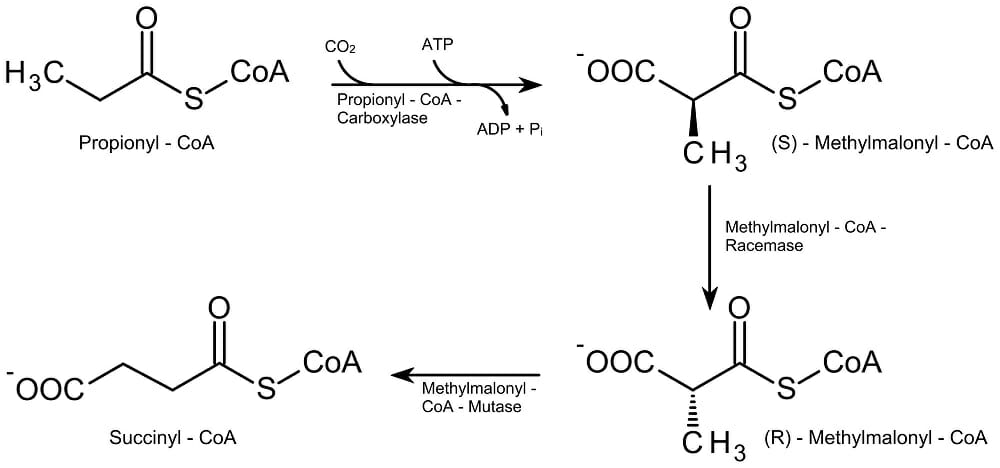
В случае нечетных цепей ацил-СоА бета-окисление происходит таким же образом, за исключением последнего этапа: вместо разрыва четырехцепочечной цепи ацил-КоА на две единицы ацетил-КоА, пятиуглеродный ацил -СоА цепь расщепляется на трехуглеродный пропионил-КоА и двухуглеродный ацетил-КоА. Другая химическая реакция затем превращает пропионил-КоА в сукцинил-КоА (см. Рисунок ниже), который входит в цикл лимонной кислоты с образованием АТФ.
Выход энергии и конечные продукты
Каждый цикл бета-окисления дает 1 FADH2, 1 NADH и 1 ацетил-КоА, что по энергии эквивалентно 17 молекулам АТФ:
- 1 FADH2 (x 2 ATP) = 2 ATP
- 1 НАДН (х 3 АТФ) = 3 АТФ
- 1 ацетил-КоА (х 12 АТФ) = 12 АТФ
- Всего = 2 + 3 + 12 = 17 спс
Однако теоретический выход АТФ выше, чем реальный выход АТФ. В действительности в каждом цикле бета-окисления образуется эквивалент примерно 12-16 АТФ.
Помимо выхода энергии, жирная цепь ацил-CoaA становится короче на два атома углерода с каждым циклом. Кроме того, бета-окисление дает большое количество воды; это полезно для эукариотических организмов, таких как верблюды, учитывая их ограниченный доступ к питьевой воде.
викторина
1. Что делает бета-окисление?A. Расщеплять углеводы.B. Расщеплять белки.C. Расщепляют жирные кислоты.D. Расщепляют жирные кислоты и белки.D. Расщеплять углеводы и белки.
Ответ на вопрос № 1
С верно. Бета-окисление расщепляет жирные кислоты для производства энергии. В частности, жирные ацил-КоА-цепи расщепляются на ацетил-КоА, FADH2, NADH и воду.
2. Каков порядок этапов цикла бета-окисления?A. Дегидрирование, гидратация, окисление и тиолизис.B. Гидратация, дегидрирование, окисление и тиолизис.C. Дегидрирование, окисление, тиолиз и гидратация.D. Гидратация, дегидрирование, тиолиз и окисление
Ответ на вопрос № 2
верно. Стадии в порядке: дегидрирование, гидратация, окисление и тиолизис.
3. Какие конечные продукты дает каждый цикл бета-окисления?A. Одна ацил-КоА, одна NADH, вода и одна цепь ацетил-КоА на два атома углерода короче.B. Одна двухуглеродная ацил-КоА, одна NADH, вода и другая ацил-КоА цепь на два атома углерода короче.C. Одна ацил-КоА, одна FADH2, одна NADH, вода и одна цепь ацетил-КоА на два атома углерода короче.D. Одна ацетил-КоА, одна FADH2, одна NADH, вода и одна ацил-КоА цепь на два атома углерода короче.
Ответ на вопрос № 3
D верно. Эффективный цикл бета-окисления дает одну (двухуглеродную) ацетил-КоА, одну FADH2, одну NADH, воду и одну ацил-КоА-цепочку на два атома углерода. FADH2, NADH и ацетил-КоА позже вступают в цикл лимонной кислоты, что дает АТФ.
Окисление жирных кислот (β-окисление)
Для преобразования энергии, заключенной в жирных кислотах, в энергию связей АТФ существует метаболический путь окисления жирных кислот до СО2 и воды, тесно связанный с циклом трикарбоновых кислот и дыхательной цепью. Этот путь называется β-окисление, т.к. происходит окисление 3-го углеродного атома жирной кислоты (β-положение) в карбоксильную группу, одновременно от кислоты отщепляется ацетильная группа, включающая С 1 и С 2 исходной жирной кислоты.
Элементарная схема β-окисления
Реакции β-окисления происходят в митохондриях большинства клеток организма (кроме нервных клеток). Для окисления используются жирные кислоты, поступающие в цитозоль из крови или появляющиеся при липолизе собственных внутриклеточных ТАГ. Суммарное уравнение окисления пальмитиновой кислоты выглядит следующим образом:
Пальмитоил-SКоА + 7ФАД + 7НАД + + 7Н2O + 7HS-KoA → 8Ацетил-SКоА + 7ФАДН2 + 7НАДН
Этапы окисления жирных кислот
1. Прежде, чем проникнуть в матрикс митохондрий и окислиться, жирная кислота должна активироваться в цитозоле. Это осуществляется присоединением к ней коэнзима А с образованием ацил-SКоА. Ацил-SКоА является высокоэнергетическим соединением. Необратимость реакции достигается гидролизом дифосфата на две молекулы фосфорной кислоты.
Ацил-SКоА-синтетазы находятся в эндоплазматическом ретикулуме, на наружной мембране митохондрий и внутри них. Существует широкий ряд синтетаз, специфичных к разным жирным кислотам.
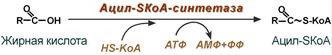
Реакция активации жирной кислоты
2. Ацил-SКоА не способен проходить через митохондриальную мембрану, поэтому существует способ переноса жирной кислоты в комплексе с витаминоподобным веществом карнитином (витамин В11). На наружной мембране митохондрий имеется фермент карнитин-ацилтрансфераза I.
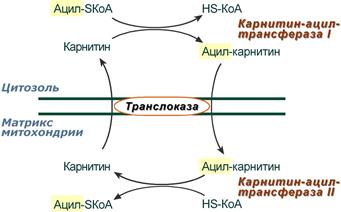
Карнитин-зависимый транспорт жирных кислот в митохондрию
Карнитин синтезируется в печени и почках и затем транспортируется в остальные органы. Во внутриутробном периоде и в первые годы жизни значение карнитина для организма чрезвычайно велико. Энергообеспечение нервной системы детского организма и, в частности, головного мозга осуществляется за счет двух параллельных процессов: карнитин-зависимого окисления жирных кислот и аэробного окисления глюкозы. Карнитин необходим для роста головного и спинного мозга, для взаимодействия всех отделов нервной системы, ответственных за движение и взаимодействие мышц. Существуют исследования, связывающие с недостатком карнитина детский церебральный паралич и феномен "смерти в колыбели".
Дети раннего возраста, недоношенные и дети с малой массой особенно чувствительны к недостаточности карнитина. Эндогенные запасы у них быстро истощаются при различных стрессовых ситуациях (инфекционные заболевания, желудочно-кишечные расстройства, нарушения вскармливания). Биосинтез карнитина недостаточен, а поступление с обычными пищевыми продуктами неспособно поддержать достаточный уровень в крови и тканях.
3. После связывания с карнитином жирная кислота переносится через внутреннюю митохондриальную мембрану транслоказой . На внутренней стороне этой мембраны фермент карнитин-ацилтрансфераза II вновь образует ацил-SКоА, который вступает на путь β-окисления.
4. Процесс собственно β-окисления состоит из 4-х реакций, повторяющихся циклически. В них последовательно происходит окисление (ацил-SКоА-дегидрогеназа), гидратирование (еноил-SКоА-гидратаза) и вновь окисление 3-го атома углерода (гидроксиацил-SКоА-дегидрогеназа). В последней, трансферазной, реакции от жирной кислоты отщепляется ацетил-SКоА . К оставшейся (укороченной на два углерода) жирной кислоте присоединяется HS-КоА, и она возвращается к первой реакции. Все повторяется до тех пор, пока в последнем цикле не образуются два ацетил-SКоА.
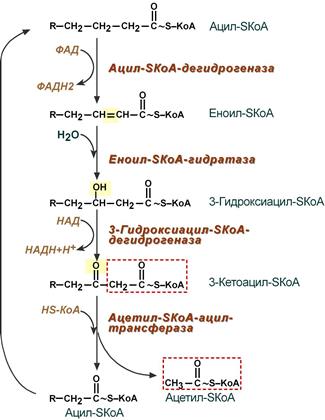
Последовательность реакций β-окисления жирных кислот
Расчет энергетического баланса β-окисления
Ранее при расчете эффективности окисления коэффициент P/O для НАДH принимался равным 3,0, для ФАДH2 – 2,0.
По современным данным значение коэффициента P/O для НАДH соответствует 2,5, для ФАДH2 – 1,5.
При расчете количества АТФ, образуемого при β-окислении жирных кислот необходимо учитывать:
Читайте также: