
Нервная трубка это кратко
Обновлено: 02.07.2024
В психиатрии в настоящее время повысился интерес к развитию мозга в связи с множеством данных, свидетельствующих о том, что разного рода нарушения в процессе его формирования и дифференцировки клеточных элементов могут играть существенную роль не только при органически обусловленных задержках психического развития, но и при эндогенных психозах (шизофрении, аффективном психозе). Когда речь идет о нарушении развития структурных и функциональных мозговых систем или искажении таких тонких процессов, как развитие межклеточных связей, обобщенно говорят о дизонтогенезе мозга.
Процесс внутриутробного развития человека делят на 2 последовательных периода: 1) эмбриональный, или зародышевый; 2) плодный, или фетальный. Границей между ними является конец 2-го — начало 3-го месяца беременности, когда зародыш превращается в плод [Фалин Л.И., 1976].
Эмбриональный период характеризуется процессами органогенеза. К концу этого периода формируются и занимают свое постоянное положение зачатки всех органов. В течение же плодного периода происходят рост и функциональное созревание органов и тканей. К концу 3-го месяца (когда зародыш достигает длины 65 мм) он приобретает все характерные особенности, свойственные человеку.
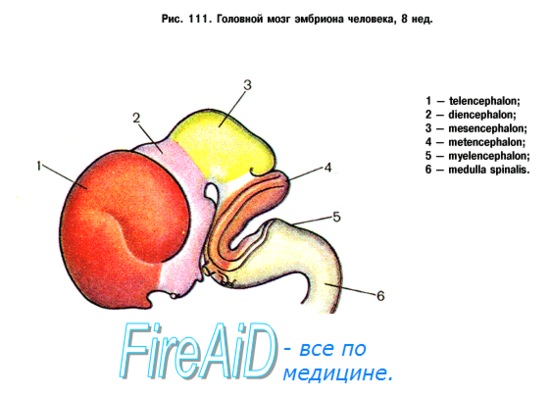
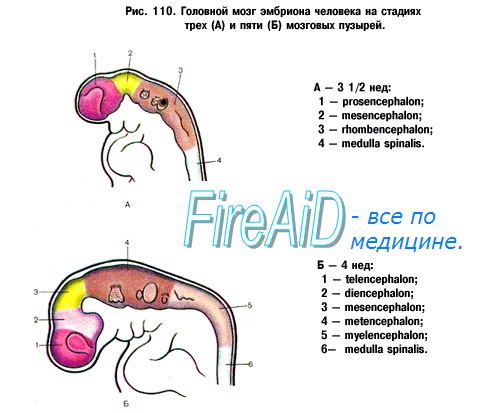
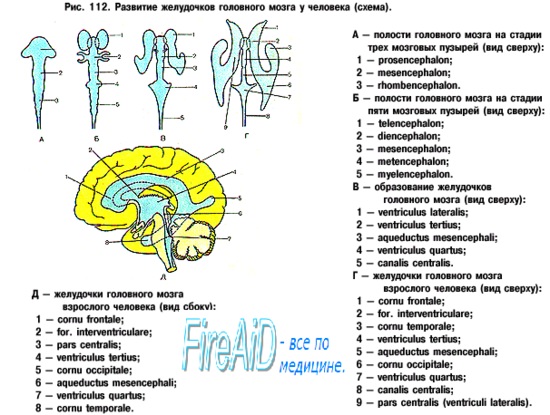
ЦНС развивается из эмбриональной закладки эктодермы — наружного зародышевого листка, который носит название нервной (мозговой) пластинки. Образование ее относится к 3-й неделе развития эмбриона. В середине 4-й недели из нее образуется нервная (мозговая, медуллярная) трубка, краниальный расширенный конец которой дает начало закладке головного мозга, а вся остальная часть превращается в спинной мозг. Нервная трубка перетяжками делится на 3 мозговых пузыря. На 5-й неделе из первичных мозговых пузырей образуются 3 главных отдела мозга: передний, средний и ромбовидный. В дальнейшем передний отдел образует конечный и межуточный мозг, а ромбовидный делится на задний и продолговатый. Из конечного мозга образуются кора и базальные ганглии, а из межуточного мозга — таламус и гипоталамус.
На следующих этапах развития мозга отдельные его участки развиваются неравномерно. Особенно быстро растут отделы, соответствующие полушариям. Они быстро покрывают все другие части мозга в виде плаща ( pallium ), разделяясь на лобную, височную и затылочную доли. Соответственно этому увеличиваются мозговые желудочки, развиваются базальные ганглии.
Одновременно с развитием нервных клеток происходят развитие и дифференцировка глиальных элементов, которые происходят из спонгиобластов, за исключением микроглии, которая, как считают большинство исследователей, формируется из мезенхимы. Спонгиобласты представляют собой тонкие вытянутые клетки, снабженные двумя основными отростками. Внутренние, более короткие, образуют внутреннюю пограничную мембрану, в то время как длинные наружные отростки пронизывают всю толщу нервной трубки. Часть спонгиобластов сохраняет первоначальное положение, превращаясь в эпендиму, выстилающую полости мозговых желудочков.. Однако большая часть спонгиобластов выселяется вместе с нейробластами в толщу плащевого слоя, где они дифференцируются в астроцитарную глию и олигодендроглию. Астроциты хорошо выявляются уже на 3-м месяце эмбриональной жизни, олигодендроглия — во второй половине эмбрионального периода, когда начинается миелинизация проводящих путей.
Сосуществование в вентрикулярной зоне мозговой трубки на самых ранних стадиях развития предшественников нейронов и глиальных клеток было подтверждено с помощью таких клеточных маркеров, как нейронспецифичная энолаза и глиальный фибриллярный кислый белок (ГФКБ). Более того, в соответствии с данными, приведенными A . M . Adinolfi и W . J . Freed (1989), эти методы позволили установить, что отсюда незрелые нейроны мигрируют путем амебовидных движений в различных направлениях к месту их дальнейшей дифференцировки вдоль глиальных отростков, которые распространяются радиально от вентрикулярной зоны к поверхности. Есть точка зрения, что эти "направляющие" глиальные клетки исчезают после созревания нейрона. Исчезновением глиальных радиальных отростков объясняют остановку дальнейшей миграции нейронов во взрослом мозге. Но, согласно другим представлениям, они превращаются в астроциты. Это не противоречит мнению о том, что нейроны созревают первыми и только после завершения нейрогенеза начинается процесс деления и дифференцировки глиальных клеток.
Развитие синапса начинается на 4—5-й неделе пренатального периода с постсинапса, т.е. с той части синаптического комплекса, который расположен на клетке, принимающей внеклеточные сигналы. К наружной клеточной мембране направляются пузырьки, первичная сборка мембраны которых осуществляется в комплексе Гольджи. Как указывает АА. Милохин (1983), каждый пузырек приносит с собой как бы своеобразный квант генетически детерминированной мембраны со всем необходимым для функции рецептора материалом. Встраивание внутриклеточного пузырька в наружную мембрану клетки — первый этап образования рецепторного поля постсинапса. Формирование пресинапса происходит путем уплотнения участка наружной мембраны нейрона. После этого уплотненные зоны мембран соседних клеток сближаются, образуя первичный недифференцированный контакт десмосомного типа в виде зон прилипания — puncta adherentia . Иногда между такими противолежащими мембранами видна пластинка несколько уплотненного или сетевидного межклеточного вещества, расположение которого соответствует будущей синаптической щели. Описанные процессы хорошо видны уже у эмбрионов 7—8 нед. развития [Иваньшина А.З., 1976]. На стадии 9—10 нед., кроме контактов типа puncta adherentia , уже имеются и асимметричные контакты с большим утолщением одной из мембран (постсинаптической) и появлением вблизи контакта пузырьков разной величины. В дальнейшем происходит не только дифференцировка, но и увеличение числа синапсов и можно видеть самые разные стадии их формирования. Следует подчеркнуть, что синапсы первоначально образуются в избыточном количестве, а затем по мере увеличения специфичности функции отдельных клеток и образуемых ими структур мозга происходит удаление (элиминация) "лишних" контактов [ Huttenlocher R . R . et al ., 1982].
Установление первых межклеточных контактов на 5-й неделе эмбриогенеза служит своего рода границей в онтогенезе нервной клетки — началом ее перехода из стадии нейробласта в стадию юного нейрона [Милохин А.А., 1983]. После приобретения первых синаптических связей нейрон уже становится частью определенной функциональной системы, которая стимулирует ее дальнейшее развитие и дифференцировку. Но и сами синапсы оказывают влияние на аксон нейрона: образование их означает конец роста по механизму "контактного торможения" (" contact inhibition "). Как указывает E . J . Ebel (1980), если бы этого торможения не было, то могла бы образоваться большая масса отростков, способная исказить развитие мозговых структур.
В процессе развития мозга выявляется чрезвычайно высокая пластичность мозговых структур, как на микроскопическом, так и ультраструктурном уровне. Речь идет о том, что при гибели части нейронов и элиминации части популяций образующихся синапсов возникают реакции на эти процессы в виде роста терминалей аксонов — коллатерального спроутинга и реактивного синаптогенеза.
Гибель нейронов в процессе развития является естественной. Это помогает сформироваться окончательной зоне не только расположения тел нейронов, но и их проекции [ Huttenlocher P . R ., 1984]. Более того, отмечается гибель части аксонов и дендритов при сохранности тела клетки, что считается важным элементом формирования проводящих путей и всех видов межклеточных связей, обеспеченных синаптическими контактами. Существуют данные о том, что в некоторых отделах ЦНС дегенерирует до 50 % синапсов только в течение 2-й постнатальной недели. В работах же I . Feinberg (1972, 1983) приводятся данные о том, что уменьшение числа синапсов в коре головного мозга человека происходит и в возрасте между 2 и 16 годами, причем особенно заметные сдвиги такого рода (потеря до 40 % синапсов) имеют место между поздним детским и ранним подростковым периодом. При этом затягивание или ускорение пубертатного периода заканчивается соответственно чрезмерным снижением числа синапсов или, напротив, образованием избытка их.
Коллатеральный спроутинг с образованием новых аксонных терминалей связан с появлением новых синаптических контактов. Важно отметить, что спроутинг отмечается не только в процессе развития мозга, но и как реакция зрелого мозга на повреждение.
Понятие пластичности нервной системы широко распространено в современной функциональной нейроанатомии. В этом случае имеется в виду способность нервной системы к функциональным и структурным перестройкам не только в виде уже охарактеризованных реакций в процессе развития и созревания мозговых структур (коллатеральный спроутинг и т.п.), но и путем изменений в системе межклеточных контактов, а также в результате перестройки функции нейрохимических систем на уровне нейротрансмиттеров.
Пластичность нервной системы не всегда может оцениваться положительно. Она может иметь и отрицательные последствия, ибо под влиянием тех или иных вредностей на этапе развития мозга могут возникать так называемые ошибочные межклеточные связи ( neuronal misconnections ), которым сейчас придается определенное значение в развитии эндогенной психической патологии [ Goldman R ., 1989].
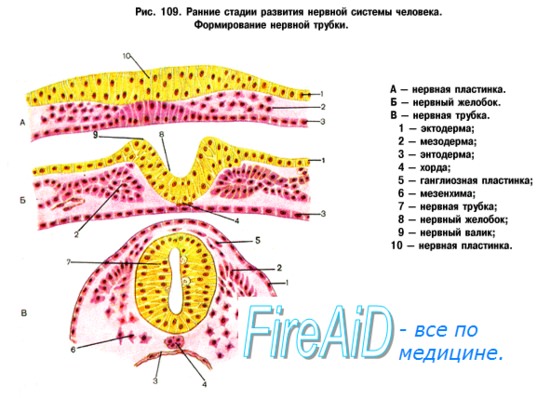
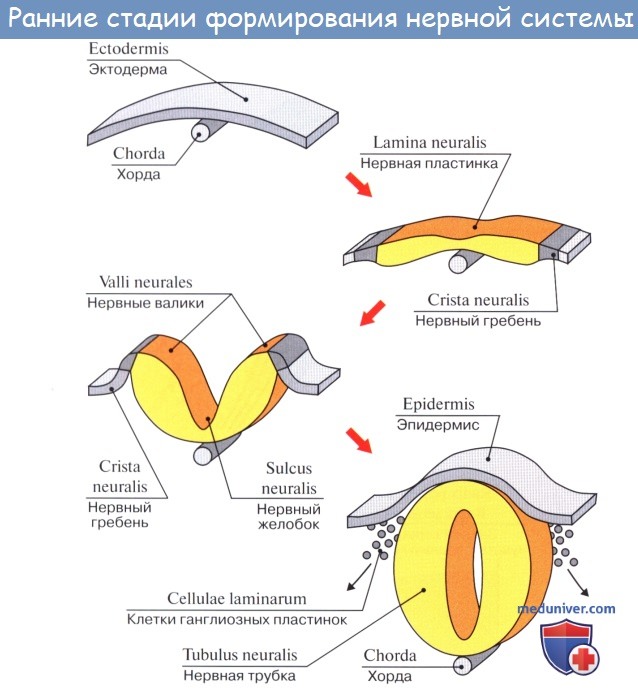
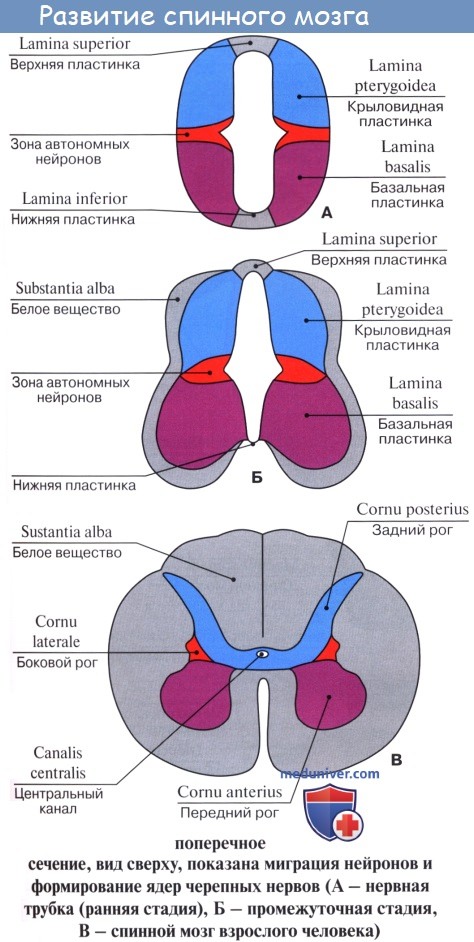
Изложенные закономерности филогенеза обусловливают эмбриогенез нервной системы человека. Нервная система происходит из наружного зародышевого листка, или эктодермы. Эта последняя образует продольное утолщение, называемое медуллярной пластинкой.
Нервная пластинка первоначально состоит только из одного слоя эпителиальных клеток. Во время замыкания ее в мозговую трубку количество клеток в стенках последней увеличивается, так что возникает три слоя: внутренний (обращенный в полость трубки), из которого происходит эпителиальная выстилка мозговых полостей (эпендима центрального канала спинного мозга и желудочков головного); средний, из которого развивается серое вещество мозга (зародышевые нервные клетки — нейробласты); наконец, наружный, почти не содержащий клеточных ядер, развивающийся в белое вещество (отростки нервных клеток — нейриты).
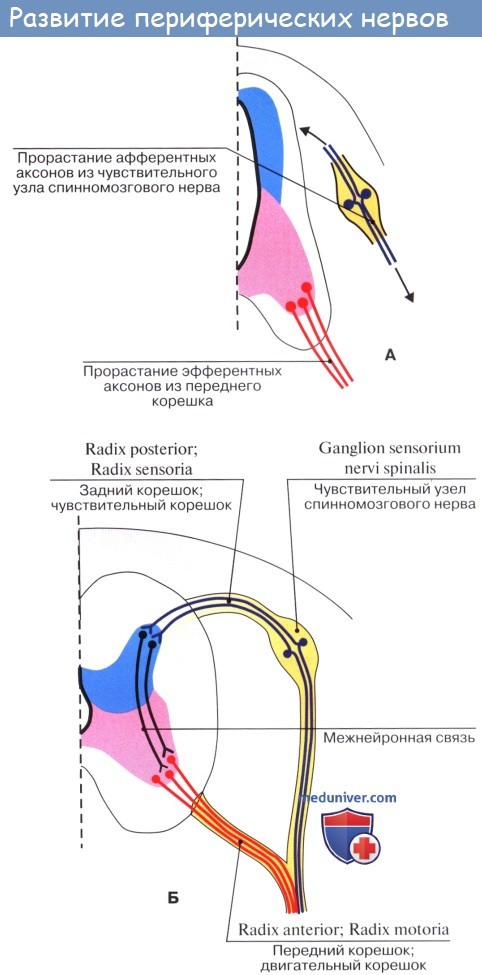
Пучки нейритов нейробластов распространяются или в толще мозговой трубки, образуя белое вещество мозга, или же выходят в мезодерму и затем соединяются с молодыми мышечными клетками (миобластами). Таким путем возникают двигательные нервы.
Чувствительные нервы возникают из зачатков спинномозговых узлов, которые заметны уже по краям медуллярной бороздки у места перехода ее в кожную эктодерму. Когда бороздка смыкается в мозговую трубку, зачатки смещаются на ее дорсальную сторону, располагаясь по средней линии. Затем клетки этих зачатков перемещаются вентрально и располагаются вновь по бокам мозговой трубки в виде так называемых нейральных гребней.
Оба нейральных гребня перешнуровываются четкообразно по сегментам дорсальной стороны зародыша, вследствие чего получается на каждой стороне ряд спинномозговых узлов, ganglia spinalia. В головной части мозговой трубки они доходят только до области заднего мозгового пузырька, где образуют зачатки узлов чувствительных черепных нервов. В ганглиозных зачатках развиваются нейробласты, принимающие вид биполярных нервных клеток, один из отростков которых врастает в мозговую трубку, другой идет на периферию, образуя чувствительный нерв.
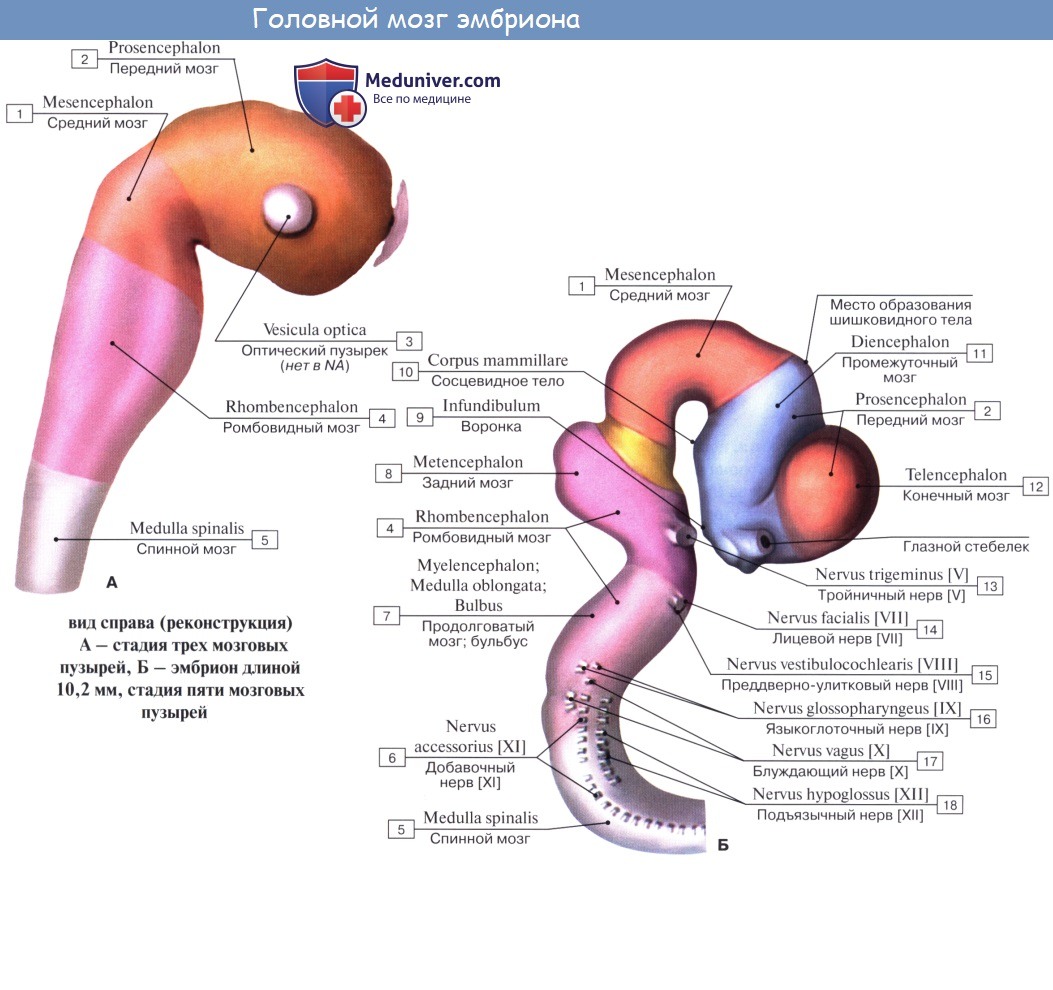
Организация ствола мозга ядерного ствола во время эмбрионального развития (по Херику)
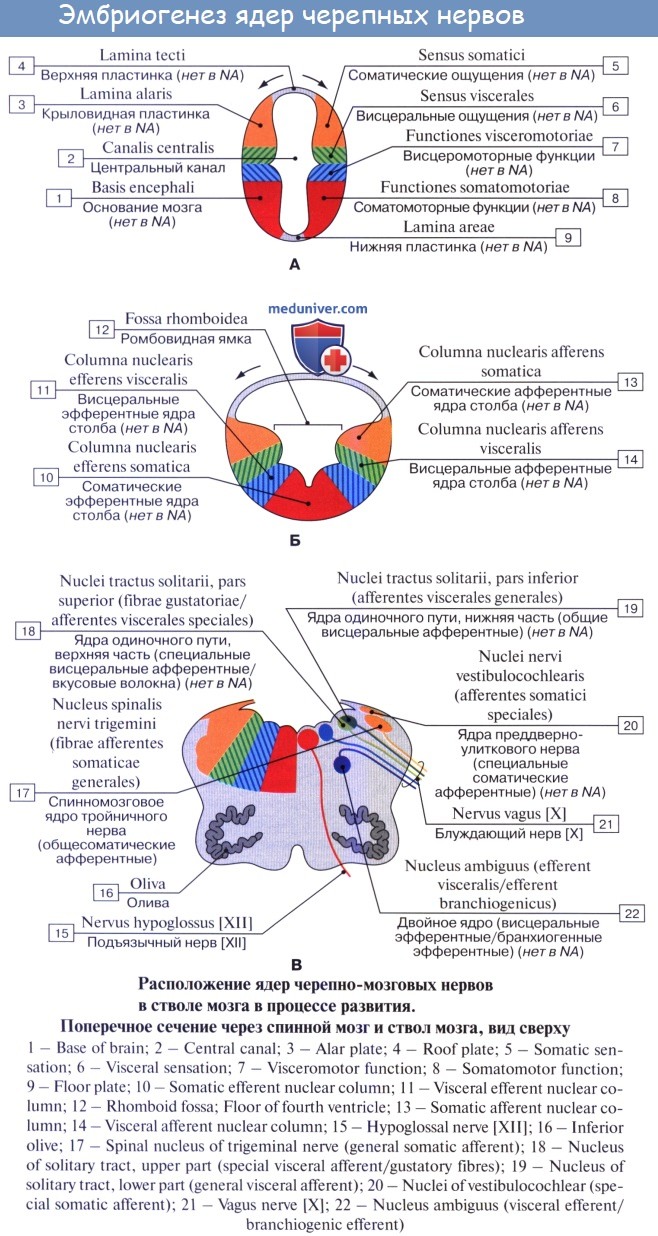
Функциональная организация ствола мозга определяется расположением ядер черепных нервов, которое можно объяснить миграцией популяций нейронов во время эмбрионального развития.
A. Начальная форма: двигательные нейроны - вентральные, чувствительные - дорсальные (дорсовентральная организация). Стрелки показывают направление миграции популяций нейронов.
Б. Ранние этапы эмбрионального развития: нейроны крыловидной пластинки (чувствительные ядра) мигрируют латерально, нейроны базальной пластинки (двигательные ядра) мигрируют медиально. Медиальнолатеральная организация.
B. Ствол мозга взрослого человека: порядок расположения четырех продольных ядерных колонн (одна соматическая двигательная, одна висцеральная двигательная, одна соматическая чувствительная, одна висцеральная двигательная). В каждой из 4 колонн ядра выполняют одинаковые функции и расположены друг над другом в краниокаудальном направлении. Ядра в соматической чувствительной и висцеральной чувствительной колоннах делятся на общие и отдельные чувствительные ядра. Аналогично в висцеральных двигательных колоннах выделяют общие (парасимпатические) и отдельные двигательные ядра. Разделение на общие и отдельные ядра не представлено в соматических двигательных ядерных колоннах.
Эмбриогенез - физиология развития от оплодотворения до рождения
Редактор: Искандер Милевски. Дата последнего обновления публикации: 15.8.2020
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
(tubus neuralis), зачаток ЦНС у хордовых. Образуется в процессе нейруляции путём углубления дна нервной пластинки, поднятия и смыкания её краёв. На переднем конце Н. т. нек-рое время сохраняется отверстие — нейропор, а на заднем — ведущий в кишечник нейроэнтерический (нервно-кишечный) канал.
зачаток нервной системы у хордовых животных. Образуется из эктодермы на стадии развития зародыша, называемой нейрулой. В дальнейшем развивается в спинной и головной мозг. У ланцетников, в отличие от позвоночных, нервная трубка сохраняется как основной элемент нервной системы. Располагается на спинной стороне животного над хордой.



Нейробласты — клетки с большим округлым ядром, плотным ядрышком и бледной цитоплазмой — дают начало всем нейронам ЦНС. Нейроны — классический пример клеток, относящихся к статической популяции. Ни при каких условиях они in vivo не способны к пролиферации и обновлению. Обонятельные нейроны (происходят из обонятельных плакод) эпителиальной выстилки носовых ходов — единственное известное исключение.
Глиобласты — предшественники макроглии [астроциты и олигодендро(глио)циты]. Все типы макроглии способны к пролиферации.
Генез клеток микроглии спорен. Согласно наиболее распространённой точке зрения, клетки микроглии относят к системе мононуклеарных фагоцитов. Достаточно вероятным представляется их нейроэктодермальный генез. В этом случае подразумевается гетерогенность популяции клеток микроглии.
Раннее развитие мозга



Морфогенетические процессы — индукция, адресная миграция клеток, направленный рост аксонов, гибель клеток, так называемые нейротрофические взаимодействия — хорошо выражены в развивающейся нервной системе. Жёсткость организации мозга определяют два момента: адресная миграция клеток и направленный рост их отростков.
Адресная миграция клеток


Направленный рост аксонов
Направленный рост аксонов, как и адресная миграция клеток, осуществляется в рамках концепции “сигнал–ответ”. Эта концепция объясняет, как нейрон узнаёт свою область иннервации и находит своего клеточного партнёра, и как в развивающемся мозге многочисленные переплетающиеся отростки нейронов устанавливают связи с высокой точностью. Отросток нейрона — аксон — сразу и без ошибок находит свои мишени. Направленный рост аксонов осуществляет конус роста.
Конус роста — мобильная терминаль отростка нейрона, специализированная на удлинение и навигацию этого отростка, впервые детально описан Сантъяго Рамон-и-Кахалем. Конус имеет на конце булавовидное утолщение (ламеллоподия), от которого отходят тонкие пальцевидные отростки — филоподии. Они растут в различных направлениях и исследуют потенциальное пространство роста аксона. Согласно представлению С. Рамон-и-Кахаля о хемотропизме, рост аксонов происходит по градиенту концентрации специфических химических факторов, вырабатываемых в мишенях. Действительно, in vitro градиенты фактора роста нервов (NGF) и других веществ (например, ацетилхолина) влияют на направление роста аксонов. Наиболее разработано представление о меченых путях, которые образованы молекулярными метками (своего рода знаками навигационной обстановки), закономерно распределёнными в потенциальном пространстве роста аксонов. По мере роста пионерский аксон последовательно считывает одну за другой метки, расположенные в межклеточном пространстве или на поверхности клеток, и растёт в нужном направлении. Вслед за ним мигрируют отростки других аксонов, совокупность которых формирует тракты в ЦНС и нервы на периферии. Примером клеток, направляющих рост аксонов, могут служить временно живущие нейроны Кахаля–Ретциуса. Ключевой момент представления о заранее размеченных путях — узнавание — обеспечивают молекулы адгезии, которые встроены в плазмолемму ламеллоподии и филоподий и взаимодействуют с комплементарными молекулами (ламинин, фибронектин, коллаген, тенасцин и др.) во внеклеточном матриксе. Это обеспечивают фиксацию конуса роста на поверхности мишени в нужном месте и в нужное время.


Характеристика
Молекул внеклеточного матрикса
Ламинин, тенасцины, коллаген, фибронектин, иммуноглобулины, аносмин‑1 ( KAL ), хондроитин/гепарансульфат протеогликаны, тромбоспондин‑1
HomA , протеогликаны, интегрины
Многие из них важны для нейроонтогенеза. Все продолжают экспрессироваться в зрелом мозге
Sema 3A‑F, Sema 4A‑G, Sema 5A и B, Sema 6A‑C, Sema 7A
Плексин A, нейропилин
Хеморепелленты, вызывают коллапс конуса роста сенсорных аксонов in vivo . Некоторые разновидности могут выступать в роли аттрактантов. Могут быть интегральными мембранными белками или секретироваться
N – CAM , PSA - N – CAM , L 1, TAG -1/аксонин‑1, DM - Grasp , кадгерины
Интегральные и мембраносвязанные гликопротеины, участвуют в прикреплении аксонов к субстрату, фасцикуляции, направленном росте, в гомофильных и гетерофильных взаимодействиях
Миелин-ассоциированный гликопротеин ( MAG ), nogo
Интегральные мембранные белки экспрессируются в зрелом миелине, ингибиторы роста аксонов in vitro и in vivo . Могут выступать в роли рецепторов для некоторых молекул внеклеточного матрикса
Рецепторов, связанных с тирозинкиназам
Ephrin A1-A5, Ephrin B1-B3
Лиганды и рецепторы, связаны с мембраной или являются трансмембранными белками, участвуют в процессах миграции клеток и роста аксонов в нейроонтогенезе
DCC, нейрогенин, Unc ‑ 5
Важны для миграции нейробластов и направленного роста аксонов в развивающемся мозге. Связывание с рецептором DCC прослежено как при позитивном, так и негативном регулировании
NGF, NT-3, BDNF, CNTF, GDNF
Trk A,B,C, p75, Lifrb, GFRa1
Нейротропная и нейротрофичекая активность
FGF, IGF, PDGF, VEGFa
FGFr1-4, flk-1, IGFr1, PDGFr, EGFr
Нейротрофическая активность, контроль пролиферации, дифференцировки и гибели клеток
TGF b , LIF, TNF, EGF
TGF b тип I; TGF b , тип II; LIFrb, gp130, TNFr1-2
Нейротрофичекая активность, поддержание дифференцировки нейронов
Возможно, участвуют в синаптогенезе. Изменяют направление роста аксонов чувствительных нейронов
Репелленты для различных аксонов и мигрирующих клеток, но и стимуляторы ветвления и удлинения аксонов
Механизм роста аксона
Перемещение конуса роста осуществляется при помощи молекулярного мотора. В конусе роста различают центральный и периферический домен. Периферический домен объединяет филоподии и часть ламеллоподии. Здесь актин полимеризуется на ведущем конце нити, что поддерживает рост аксона в ортоградном направлении, но тянется в обратном (ретроградном) направлении за счет работы миозинового мотора. Ретроградное перемещение актина сдерживает проникновение микротрубочек в периферический домен. Сигналы взаимодействия поверхностных рецепторов с лигандами угнетают ретроградное перемещение актиновых нитей и равновесие между сборкой и перемещением актиновых нитей в ортоградном направлении, с одной стороны, и ретроградным перемещением актиновых нитей за счет работы мотора, с другой стороны, нарушается в пользу первого и в итоге обеспечивает перемещение (рост) всей структуры конуса роста в ортоградном направлении. Остановка ретроградного перемещения актина позволяет микротрубочкам проникать в периферический домен. Этот рост микротрубочек осуществляется путём полимеризации тубулина. В результате аксон удлиняется.
Реакции актина на ведущем конце ламеллоподии . Сеть цитоскелета на ведущем конце ламеллоподии содержит интенсивно ветвящиеся актиновые нити с (+)–концом, направленным в сторону роста, и (–)–концом, образующим угол ~70º с другими нитями. В состоянии покоя (+)-конец актиновой нити обычно покрыт кэппинг–белком. Под действием стимула происходит полимеризация актина на свободном (+)–конце нити, расположенной в ведущей части конуса роста. Нить удлиняется и возникает усилие в направлении поверхностной мембраны. В диссоциации кэппинг–белка и освобождении (+)–конца актиновой нити имеют значение увеличение уровня инозитолдифосфата, разрезание нити ADF/кофилином и нуклеация актина, зависимая от комплекса Arp2/3. Этот комплекс активируется при связывании с белком N – WASP ( W iskott - A ldrich S yndrome P rotein , тяжелая врожденная нейтропения, тромбоцитопения), одна из форм которого, в свою очередь, активируется при участии Cdc42.
Физиологическая гибель клеток
Запрограммированная массовая смерть нейронов происходит на строго определённых этапах онтогенеза. Естественная гибель нейронов прослежена как в ЦНС, так и в периферической нервной системе. Объём субпопуляции гибнущих нейронов оценивают в широком интервале, от 25 до 75%. Иногда в популяции погибают все нейроны (например, несущие метку для направленного роста аксонов). Выраженная гибель нейронов в сформированной нервной ткани наблюдается при дегенеративных заболеваниях нервной системы, таких как болезнь Альцхаймера, Паркинсона, Хантингтона, Кройтцфельдта–Якоба, боковой амиотрофический склероз и др.
Эта концепция предусматривает информационный обмен между нейронами, отличающийся от химической передачи возбуждения в синапсах. Согласно концепции, такой обмен поддерживает фенотип взаимодействующих клеток на уровне, адекватном выполнению их функции. Нейротрофические взаимодействия реализуются через нейротрофические факторы. Эти химические факторы вырабатываются в одних нейронах или в ненервных клетках и воздействуют на другие нейроны. Нейротрофические факторы поддерживают выживание и дифференцировку нейронов в нейроонтогенезе и при регенерации. Наиболее изученным нейротрофическим фактором является фактор роста нервов ( NGF ).
Нейроны относят к статической клеточной популяции. Это означает, что в постнатальном онтогенезе не происходит образования новых нейронов. Следовательно, погибающие нейроны не восстанавливаются. Но из этого не следует, что в нервной системе отсутствует регенерация. Она осуществляется за счёт восстановления целостности повреждённых нейронов, роста их отростков, размножения глиальных и шванновских клеток. Всё это, а также изменение связей между регенерирующими и интактными клетками при благоприятных условиях, может привести к восстановлению функции в нервной системе.
Нейроны (термин предложил Вильгельм фон Вальдейер) — главные клеточные типы нервной ткани. Эти возбудимые клетки осуществляют передачу электрических сигналов (между собой при помощи нейромедиаторов в синапсах) и обеспечивают способность мозга к переработке информации.



Перикарион содержит ядро, комплекс Гольджи, гранулярную эндоплазматическую сеть, митохондрии, лизосомы, элементы цитоскелета.
Ядро нейрона имеет мелкодисперсный хроматин и ядрышко. В силу относительно большого диаметра ядро в световом микроскопе выглядит (особенно в крупных нейронах) как оптически пустое. Ядрышко крупное и резко базофильное.
Комплекс Гольджи хорошо развит, особенно в крупных нейронах. Его особенность — расположение между ядром и местом отхождения аксона, что отражает мощный транспорт белков, синтезированных в гранулярной эндоплазматической сети перикариона, в аксон.
Аксонный холмик — свободная от гранулярной эндоплазматической сети и рибосом область перикариона, содержащая много микротрубочек и нейрофиламентов, место, где начинается аксон и генерируется потенциал действия.
Гранулярная эндоплазматическая сеть. В перикарионе и дендритах развита гранулярная эндоплазматическая сеть. Она соответствует глыбкам хроматофильного вещества, которые впервые обнаружил Франц Ниссль при окраске метиленовым синим, почему её в нейронах иногда называют веществом Ниссля (тигроид). Тигролиз — распыление хроматофильного (тигроидного) вещества, отражающее глубокие дистрофические изменения при нарушении целостности нейрона ( например , при сдавлении или перерезке аксона).
Митохондрии многочисленны. Значительные энергетические потребности нервных клеток обеспечива ет преимущественно аэробный метаболизм, почему нейроны крайне чувствительны к гипоксии.
Пигменты . В нейронах (особенно с возрастом) накапливается липофусцин. Нейроны некоторых ядер мозга нормально содержат иные пигменты, почему эти образования и получили своё название (substantia nigra, locus coeruleus).
Отростки, отходящие от перикариона, — аксон и дендриты (рис. 8–13). Отростки нейрона участвуют в образовании синапсов.
Аксон (нейрит) — длинный отросток, как правило, не ветвящийся, но образующий концевые разветвления, содержащие синаптические пузырьки; проводит пачки импульсов (спайки) от перикариона. Объём аксона может достигать 99% суммарного объёма нейрона. Длина аксона может быть весьма значительной — десятки сантиметров.
Синтез белка в аксоне . Считается общепринятым, что синтез белка в нейроне происходит только в перикарионе и дендритах. Однако в последнее время появились достаточно убедительные доказательства синтеза белка в аксоне. Многочисленные белки аксоплазмы синтезируются в перикарионе и перемещаются в составе медленного аксонного транспорта. Длина аксона варьирует от нескольких микрон до метра, и транспортировка белковых молекул по аксону может занять дни, недели и даже месяцы. Следовательно, при срочном ответе нейрона на изменившиеся условия функционирования локальный синтез белка экономит время и энергию, необходимые для транспортировки вновь синтезированного белка в нужный компартмент аксона. Учитывая тот факт, что белки имеют определённый период полураспада и во время медленного транспорта по аксону постоянно подвергаются деградации можно предположить, что внутриаксонный синтез белка также необходим для поддержания белкового состава аксоплазмы и массы аксона в целом.
Цитоскелет нейронов состоит из микротрубочек, промежуточных филаментов (нейрофиламенты) и микрофиламентов.

Читайте также: