
Нейтральные мутации это кратко
Обновлено: 06.07.2024
Кроме того, трудно представить себе такой ландшафт, который точно отразил бы реальное соотношение вредных, полезных и нейтральных мутаций. Как правило, большинство мутаций нейтральны — по крайней мере у высших организмов, эукариот, у которых в геномах много участков, слабо влияющих на приспособленность. На втором месте по численности вредные мутации, на последнем — полезные. Нелегко вообразить склон такой формы, чтобы из каждой точки было больше разных путей, параллельных горизонту, чем путей, ведущих вверх или вниз. Но реальные склоны реальных ландшафтов приспособленности именно таковы.
Нейтральные мутации — это по определению такие мутации, которые не влияют на приспособленность, или, что то же самое, не подвергаются действию отбора. Нам пора познакомиться поближе с этим самым распространенным в природе классом мутаций. Забудем на время о ландшафте приспособленности и обратимся к другой модели, которая описывает процессы, происходящие с генетическими вариантами (аллелями) в популяции.
Допустим, у нас есть маленькая популяция мюмзиков из 40 особей. Для простоты примем, что мюмзики гаплоидны, т. е. имеют одинарный набор хромосом — один-единственнный экземпляр генома, а не два, как у нас с вами, диплоидных организмов. Достигнув возраста в один год, каждый мюмзик рожает нескольких детенышей, а потом сразу умирает от старости. Все детеныши одинаковые, но выжить из них могут только 40 — больше мюмзиков просто не помещается в старой железной бочке на дне пруда, где живет изучаемая популяция. Смертность детенышей случайна — ровно 40 случайно выбранных везунчиков станут взрослыми и через год произведут потомство.
Поскольку смертность случайна, приспособленность мюмзиков определяется только их плодовитостью, т. е. числом детенышей. Плодовитость зависит от генотипа. Допустим, у мюмзика есть один ген, влияющий на плодовитость. Обозначим его буквой А. Между прочим, не смейтесь: такая модель вполне годится для изучения некоторых законов популяционной генетики.
Некогда у всех мюмзиков был только один вариант (аллель) гена А. Обозначим его А1. Но потом у одной особи произошла мутация, и в результате появился второй вариант гена — аллель А2. Допустим (опять-таки для простоты), что в нашей популяции, когда мы приступили к ее изучению, у половины мюмзиков ген А был представлен первым вариантом, у другой половины — вторым. Таким образом, частота аллеля А2 равна 0,5 (q2 = 0,5) и частота аллеля А1 такая же (q1 = 0,5).
Нам нужно ответить на вопрос: как будет меняться со временем частота аллеля A2, если мутация, которая привела к его возникновению, была нейтральной?
Раз мутация была нейтральной, значит, плодовитость обладателей обоих аллелей одинакова. Допустим, они все рожают ровно по десять детенышей. Разумеется, потомство наследует родительский аллель гена А.

Здесь самое время сказать, что случайные, не зависящие от приспособленности изменения частот аллелей называются генетическим дрейфом. Под властью дрейфа находятся все нейтральные мутации (а в маленьких популяциях отчасти также и вредные с полезными, но об этом позже).
Можно ли вычислить вероятность того, что нейтральный аллель в конце концов зафиксируется, а не элиминируется? Да, это просто. В нашем примере аллели А1 и А2 исходно находились в равном положении, потому что у обоих была частота 0,5. Очевидно, что в такой ситуации шансы на фиксацию у них должны быть одинаковыми и равными 0,5. В половине случаев А1 зафиксируется, А2 элиминируется, в половине — наоборот.
Ну а если конкурирующих нейтральных аллелей не два, а, скажем, четыре и начальные частоты у них тоже одинаковые (0,25)? В этом случае дело кончится фиксацией одного из аллелей и элиминацией трех остальных, причем для каждого аллеля вероятность фиксации равна 0,25. Таким образом, очевидно, что вероятность фиксации нейтральной мутации в будущем просто-напросто равна ее частоте в данный момент: Pfix = q.
Сколько нейтральных мутаций будет фиксироваться в популяции в каждом поколении? (Имеются в виду мутации уже не в одном и том же, а в разных генах.) Если мы сумеем это вычислить, то получим прекраснейший инструмент — молекулярные часы. Тогда мы сможем по количеству нейтральных генетических различий определить, когда жил последний общий предок сравниваемых организмов.
Теперь нужно понять, какая часть из этих U ? N только что появившихся мутаций в итоге зафиксируется. Это и будет искомая величина — темп фиксации нейтральных мутаций в генофонде популяции за одно поколение. Обозначим ее буквой V.
Нам остался один шаг до молекулярных часов. После того как два вида, произошедшие от общего предка, разделились, в их генофондах независимо накапливаются нейтральные мутации. За время t (измеряемое в поколениях) первый вид накопит V ? t = U ? t нейтральных мутаций, и второй вид накопит столько же. Совместными усилиями они накопят 2U ? t нейтральных отличий друг от друга. Зная темп мутагенеза U и подсчитав число различий между геномами сравниваемых видов (обозначим его буквой D), определяем время жизни их последнего общего предка: t = D/2U. Это и есть знаменитые молекулярные часы.
К счастью, разные участки генома накапливают нейтральные изменения с очень разной скоростью[18]. Быстро меняющиеся участки ДНК используются для датировки недавних событий, а те, что меняются медленно, хороши для датировки событий глубокой древности.
Данный текст является ознакомительным фрагментом.
Продолжение на ЛитРес
Глава 10 Следы, оставленные три с половиной миллиона лет назад! Дарт, Брум и современные исследователи. Дрейф континентов. Поименный список гоминидов. Люси и ее сородичи.
Глава 10 Следы, оставленные три с половиной миллиона лет назад! Дарт, Брум и современные исследователи. Дрейф континентов. Поименный список гоминидов. Люси и ее сородичи. Столь длительная сохранность доисторических следов у Лаетоли – случай фантастический, но не
ГЛАВА 3 Эволюция земной коры. Дрейф континентов и спрединг океанического дна. Мантийная конвекция
ГЛАВА 3 Эволюция земной коры. Дрейф континентов и спрединг океанического дна. Мантийная конвекция Горные породы, формирующие кору Земли, как мы помним, бывают изверженные — первичные, образовавшиеся при охлаждении и затвердевании магмы, и осадочные — вторичные,
Движение — простейшая форма поведения
Движение — простейшая форма поведения Тропизмы Первое наиболее четкое различие между животными и растениями ясно каждому: растения не могут передвигаться, тогда как животные этим свойством обладают. И тем не менее именно движение растений (поворот к солнцу цветов)
Дрейф и отбор: кто кого?
Дрейф и отбор: кто кого? Генетический дрейф царствует над нейтральными мутациями (аллелями), отбор — над полезными и вредными. Отбор, повышающий частоту полезных мутаций, называют положительным. Отбор, отбраковывающий вредные мутации, — отрицательным, или
ДРЕЙФ ГЕНОВ
ДРЕЙФ МАТЕРИКОВ
ДРЕЙФ МАТЕРИКОВ В 1912 году немецкий ученый Альфред Вегенер предположил, что около 200 миллионов лет назад все материки Земли составляли единый массив суши, который он назвал Пангеей. В последующие 200 миллионов лет Пангея разделилась на несколько материков, которые стали
Глава 6. Движение
Глава 6. Движение Природы зуб и коготь ал” — это, должно быть, одна из самых цитируемых, к месту и не к месту, аллюзий на Дарвина в англоязычной литературе. Но как бы не к месту ее иногда ни цитировали, эта фраза верно передает если не истинную суть естественного отбора, то,
9.3. Амебоидное движение
9.3. Амебоидное движение Амебы движутся с помощью объемного тока своей цитоплазмы в вырастающие выступы — псевдоподии. В норме они перемещаются по поверхности твердых объектов посредством непрерывного растягивания своих передних концов. Но если до псевдоподий
Движение живых объектов
Движение живых объектов Глядя на то, как двигается тот или иной объект, уже можно сказать, живое ли это существо или просто лист, летящий по ветру. А можно сказать и намного больше. Можно, например, увидеть, что это человек, и увидеть, что он делает. Информации для этого
Движение
3. Ощущение и движение
3. Ощущение и движение Жара повсюду. Ее нельзя больше не замечать. Воздух раскален, как в доменной печи, — он настолько горячий, что мои глаза, защищенные очками, кажутся прохладными по сравнению с остальным лицом. Руки тоже прохладные, но на тыльной стороне перчаток
Движение
Движение — это сон
Движение — это сон Чего только не делают люди во сне — разговаривают, улыбаются, смеются, плачут, стонут, чмокают, морщатся, вздрагивают, жестикулируют, скрежещут зубами и даже бродят по дому.Людей, которые бы не двигались во сне, не существует. Клейтман исследовал одного
Внутреннее ухо — движение желе и колебание волосков
Внутреннее ухо — движение желе и колебание волосков Представьте себе, что мы заходим в слуховой проход, проходим сквозь барабанную перепонку, мимо трех косточек среднего уха и оказываемся глубоко внутри черепа. Здесь расположено внутреннее ухо — заполненные
Нейтральная мутация * нейтральная мутацыя * neutral mutation — 1. Мутация, не имеющая селективных преимуществ или отрицательного влияния на организм, в котором она произошла, напр., мутация в криптических генах или некодирующей ДНК. 2. Генетическое изменение, фенотипическая экспрессия которого не приводит к изменениям величины адаптивности, или приспособленности, организма к условиям среды, имеющимся в настоящее время (Теория нейтральных мутаций и случайного дрейфа генов в молекулярной эволюции).
Генетика. Энциклопедический словарь. - Минск: Белорусская наука . Картель Н. А., Макеева Е. Н., Мезенко А. М. . 2011 .
Смотреть что такое "Нейтральная мутация" в других словарях:
нейтральная мутация — neutralioji mutacija statusas T sritis augalininkystė apibrėžtis Mutacija, neturinti poveikio organizmui, kuriame įvyko. atitikmenys: angl. neutral mutation rus. нейтральная мутация … Žemės ūkio augalų selekcijos ir sėklininkystės terminų žodynas
Мутация нейтральная м молчащая — Мутация нейтральная, м. молчащая * мутацыя нейтральная; м., якая маўчыць * neutral mutation or silent m. 1. Мутация, которая никак не влияет на приспособленность особи. 2. Мутация, не имеющая фенотипического проявления. 3. Изменение в… … Генетика. Энциклопедический словарь
Мутация — Эта статья о термине из области биологии, см. также Мутация (значения) Главный мутаген табачного дыма бензпирен связанный с одним из нуклеотидов молекулы … Википедия
Нонсенс-мутация — Нонсенс мутация точечная мутация в последовательности ДНК, которая приводит к появлению стоп кодона, в результате чего происходит преждевременная терминация синтеза нужного белка. Обычно такой фрагмент не может выполнять функции изначально… … Википедия
neutral mutation — neutralioji mutacija statusas T sritis augalininkystė apibrėžtis Mutacija, neturinti poveikio organizmui, kuriame įvyko. atitikmenys: angl. neutral mutation rus. нейтральная мутация … Žemės ūkio augalų selekcijos ir sėklininkystės terminų žodynas
neutralioji mutacija — statusas T sritis augalininkystė apibrėžtis Mutacija, neturinti poveikio organizmui, kuriame įvyko. atitikmenys: angl. neutral mutation rus. нейтральная мутация … Žemės ūkio augalų selekcijos ir sėklininkystės terminų žodynas
Эволюция — Эта статья о биологической эволюции. Другие значения термина в заглавии статьи см. на Эволюция (значения). Фи … Википедия
Мутант — Эта статья о термине из области биологии, см. также Мутация (значения) Главный мутаген табачного дыма бензопирен связанный с одним из нуклеотидов молекулы ДНК. Мутация стойкое (то есть такое, которое может быть унаследовано потомками данной… … Википедия
Мутанты — Эта статья о термине из области биологии, см. также Мутация (значения) Главный мутаген табачного дыма бензопирен связанный с одним из нуклеотидов молекулы ДНК. Мутация стойкое (то есть такое, которое может быть унаследовано потомками данной… … Википедия
Мутации — Эта статья о термине из области биологии, см. также Мутация (значения) Главный мутаген табачного дыма бензопирен связанный с одним из нуклеотидов молекулы ДНК. Мутация стойкое (то есть такое, которое может быть унаследовано потомками данной… … Википедия
Виды мутаций у человека. Варианты
Последовательность ядерной ДНК у любых двух человек идентична почти на 99,9%. Только очень небольшая доля последовательности ДНК различается у разных людей, обеспечивая генетическую изменчивость. Некоторые различия в последовательности ДНК не имеют влияния на фенотип, тогда как другие — непосредственные причины болезней. Между двумя крайностями — изменения, ответственные за генетически предопределенную фенотипическую изменчивость в анатомии и физиологии, переносимость пищи, реакции на лечение или побочные эффекты медикаментов, восприимчивость к инфекциям, склонность к опухолям и, возможно, даже изменчивость в различных чертах личности, спортивных способностях и художественном таланте.
Одно из важных понятий генетики человека и медицинской генетики — то, что генетические болезни — только наиболее очевидное и часто крайнее проявление генетических различий, один конец непрерывного спектра изменений от редких вариантов, вызывающих болезнь, через более частые варианты, увеличивающие восприимчивость к болезни, до наиболее частых изменений, не имеющих явного отношения к болезни.
Виды мутаций у человека
Мутация — любое изменение в последовательности нуклеотидов или расположения ДНК. Мутации можно классифицировать на три категории: влияющие на количество хромосом в клетке (геномные мутации), изменяющие структуру отдельных хромосом (хромосомные мутации) и изменяющие индивидуальные гены (генные мутации). Геномные мутации — изменения числа неповрежденных хромосом (анеуплоидии), возникающие вследствие ошибок в расхождении хромосом в мейозе или митозе.

Хромосомные мутации — изменения, затрагивающие только часть хромосомы, например частичные дупликации, делеции, инверсии и транслокации, которые могут происходить спонтанно или возникать вследствие аномального расхождения транслоцированных хромосом в ходе мейоза. Генные мутации — изменения в последовательности ДНК ядерного или митохондриального генома, от мутации в единственном нуклеотиде до изменений, захватывающих много миллионов пар оснований. Множество типов мутаций представлены разнообразными аллелями в отдельных локусах при более чем тысяче разных генетических заболеваний, а также среди миллионов вариантов ДНК, обнаруживаемых во всем геноме в нормальной популяции.
Описание разных мутаций не только увеличивает осведомленность о генетическом разнообразии человека и хрупкости человеческого генетического наследия, но также содействует получению информации, необходимой для обнаружения и скрининга генетических болезней в конкретных семьях риска, а также — для некоторых болезней — в популяции в целом.
Геномная мутация, приводящая к утрате или дублированию целой хромосомы, изменяет дозу и, таким образом, уровень экспрессии сотен или тысяч генов. Аналогично затрагивающая большую часть одной или нескольких хромосом хромосомная мутация также может влиять на экспрессию сотен генов. Даже небольшая генная мутация может иметь большие последствия, в зависимости от того, какой ген затронут и к чему приводит изменение в экспрессии этого гена. Мутация гена в виде изменения единственного нуклеотида в кодирующей последовательности может вести к полной утере экспрессии гена или образованию белка с измененными свойствами.
Некоторые изменения ДНК, тем не менее, не имеют фенотипических эффектов. Хромосомная транслокация или инверсия может не влиять на критическую часть генома и абсолютно не иметь фенотипических эффектов. Мутация в пределах гена может не иметь эффекта вследствие того, что либо не изменяет аминокислотную последовательность полипептида, либо, даже если это происходит, изменение в закодированной аминокислотной последовательности не изменяет функциональные свойства белка. Следовательно, не все мутации имеют клинические последствия.
Все три типа мутаций происходят со значимой частотой во множестве разных клеток. Если мутация происходит в ДНК половых клеток, она может передаваться последующим поколениям. В отличие от этого, соматические мутации происходят случайным образом только в части клеток определенных тканей, приводя к соматическому мозаицизму, наблюдаемому, например, при многих опухолях. Соматические мутации не могут передаваться последующим поколениям.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Нейтральные мутации – не влияют на приспособленность.
Приспособленность – это эффективность размножения.
Например, если каждый носитель аллеля а1 оставляет в среднем 5 жизнеспособных потомков, а каждый носитель аллеля а2 (т.е. другого варианта того же гена А) оставляет в среднем 4 столь же жизнеспособных потомков, то приспособленность аллеля А1 по отношению к аллелю А2 = 5/4 = 1.25
Как правило, абсолютную приспособленность не измеряют – это не очень понятно, как сделать. А вот относительную приспособленность померить можно: просто посмотреть, с какой скоростью размножаются носители данного аллеля по сравнению со всеми остальными (т.е. с теми, у кого этого аллеля нет).
Если же Fa1/Fa2 ≈ 1, то это нейтральное различие. Если в генофонде был сначала только аллель а1, а потом в результате мутации появился аллель а2, причем Fa1/Fa2 ≈ 1, то это значит, что мутация, в результате которой возник аллель а2, была нейтральной.
Разберемся, как будет меняться частота нейтральной мутации (нейтрального аллеля) со временем, с течением поколений.
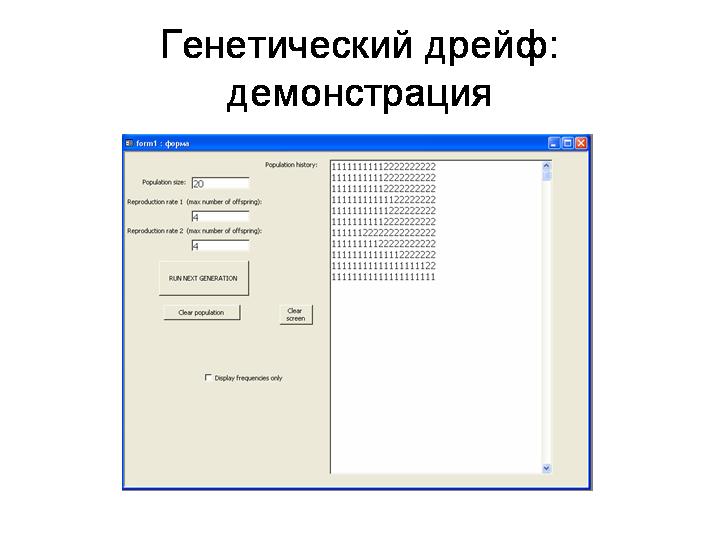
· Частота нейтрального аллеля в популяции изменяется по закону случайных блужданий. A random walk is a mathematical formalisation of a trajectory that consists of taking successive random steps. Случайные колебания частот аллелей, не связанные с действием отбора, называются генетическим дрейфом.
· Конечный исход этих случайных блужданий строго предопределен. Это всегда одно из двух: либо аллель зафиксируется (достигнет частоты q=1), либо он элиминируется (достигнет частоты q=0). Третьего не дано. Одно из двух событий рано или поздно ОБЯЗАТЕЛЬНО произойдет. Случайно блуждая, частота аллеля непременно когда-нибудь наткнется либо на верхнюю планку (фиксация), либо на нижнюю (элиминация).
Формула 1. Чему равна вероятность фиксации (Pfix) нейтральной мутации? Если исходная частота q=0.5, то совершенно ясно, что аллель с равной вероятностью зафиксируется или элиминируется, т.е. Pfix = 0.5.
Представим себе ситуацию, когда у нас исходно есть не 2, а 4 нейтральных аллеля с одинаковыми частотами: q1=q2=q3=q4=0.25
Таким образом, можно считать доказанным, что конечным результатом дрейфа будет фиксация одного из аллелей и элиминация трех других. Ясно, что, поскольку исходно все 4 аллеля были в абсолютно равном положении, шансы на фиксацию у них должны быть одинаковы. В сумме эти шансы дают 1 (один из аллелей точно зафиксируется).
Следовательно, Pfix1 = Pfix2 = Pfix3 = Pfix4 = ј = 0.25
Вероятность фиксации нейтрального аллеля равна его исходной частоте (т.е. частоте в тот момент, для которого мы пытаемся рассчитать вероятность).

Поскольку мы могли с тем же успехом взять не 2, не 4, а любое другое число нейтральных аллелей, очевидно, что:
Вероятность фиксации нейтрального аллеля равна его частоте. Это первая из трех формул, которую мы с вами должны вывести. А чему равна вероятность элиминации аллеля? Очевидно, 1-q.
Формула 2. Следующий очень важный вопрос – определить, сколько нейтральных мутаций будет фиксироваться в популяции в каждом поколении. Если мы сможем это установить, то мы обретем великий инструмент – молекулярные часы. Зная, с какой скоростью фиксируются в популяции нейтральные мутации, и зная, сколько их накопилось за какой-то период, мы сможем вычислить длительность этого периода. Например, сколько разных нейтральных мутаций есть в геномах человека и шимпанзе? Это можно подсчитать. Если бы мы еще знали, с какой скоростью фиксируются нейтральные мутации, мы могли бы на основе данных о числе накопившихся мутаций вычислить, как давно разошлись эволюционные линии предков человека и шимпанзе, понять, когда жил их последний общий предок.
Итак, сколько же нейтральных мутаций будет фиксироваться в популяции за одно поколение? Вывод этой формулы – это абсолютнейший шедевр биологической математики. Надеюсь, вы оцените.
Итак, от чего зависит скорость накопления нейтральных мутаций?
Очевидно, она зависит от того, с какой скоростью они появляются – то есть от скорости нейтрального мутагенеза, а также от того, какая часть вновь появляющихся нейтральных мутаций будет фиксироваться (а не элиминироваться).
· V – искомая величина: среднее количество нейтральных мутаций, которые будут фиксироваться в генофонде популяции за одно поколение.
· M – темп появления новых нейтральных мутаций (на всю популяцию). Это среднее количество новых нейтральных мутаций, появляющихся в популяции за одно поколение.
· m – то же, в расчете на одну особь. Это среднее количество новых нейтральных мутаций, имеющихся в геноме родившейся особи. Эту величину реально можно измерить.
· Pfix – доля (или процент) возникших в популяции нейтральных мутаций, которые в конечном счете зафиксируются. Эта доля, очевидно, есть то же самое, что вероятность фиксации такой мутации.
· N – численность популяции. Это тоже можно измерить (правда, она могла меняться в прошлом, и мы не знаем, как она менялась – это плохо).
Приступим к выводу формулы.
Очевидно, что V = M• Pfix
Искомая величина V равна числу мутаций, появляющихся за поколение, умноженному на ту долю, которая зафиксируется из них в конечном счете.
Чему равно M? Очевидно, что M = N•m
Это, надеюсь, ясно без пояснений.
А как найти Pfix, т.е. как выразить эту величину через те величины, которые можно измерить? Ну, мы ведь уже знаем, что Pfix = q , т.е. для каждой мутации вероятность зафиксироваться равна частоте этой мутации в данный момент. Новая, только что появившаяся мутация, очевидно, изначально есть только у одной особи. То есть ее частота q=1/N. Следовательно, и вероятность ее фиксации равна 1/N. Следовательно, из всех новых мутаций зафиксируется их часть, равная 1/N. Вот вам и искомое значение Pfix:
V = M• Pfix M = N•m Pfix =1/N
Подставляем в первую формулу две другие, получаем:
Численность популяции великолепным образом сокращается! Итого:
V = m
Поэтому у нас есть молекулярные часы. Мы их имеем благодаря генетическому дрейфу и благодаря тому, что многие мутации нейтральны. Принцип молекулярных часов:
где D – количество нейтральных различий между двумя видами, t – время, прошедшее с тех пор, когда жил последний общий предок этих двух видов. Двойка тут появляется потому, что нейтральные мутации накапливались обоими видами. Каждый из них накопил за время t количество нейтральных мутаций, равное D/2.
Дарвин (рекомендации по эссе).

Главный вклад Дарвина в умственное развитие человечества. До Дарвина нам был известен только ОДИН механизм возникновения сложных объектов, предназначенных для какой-то цели или функции – разумный дизайн. Пейли и часы. Все подобные объекты трактовались как результат чьего-то сознательного проектирования. Или придумывались некие невнятные дополнительные сущности – жизненная сила, стремление к совершенству и т.п. (ламаркизм, номогенез). Что с научной точки зрения ничуть не лучше гипотезы о разумном дизайне. Дарвин открыл другой механизм – механизм самопроизвольного возникновения сложных объектов, приспособленных для выполнения функций – объектов, которые выглядят (для нас) так, как будто бы их кто-то нарочно спроектировал.
Нейтральные мутации – не влияют на приспособленность.
Приспособленность – это эффективность размножения.
Например, если каждый носитель аллеля а1 оставляет в среднем 5 жизнеспособных потомков, а каждый носитель аллеля а2 (т.е. другого варианта того же гена А) оставляет в среднем 4 столь же жизнеспособных потомков, то приспособленность аллеля А1 по отношению к аллелю А2 = 5/4 = 1.25
Как правило, абсолютную приспособленность не измеряют – это не очень понятно, как сделать. А вот относительную приспособленность померить можно: просто посмотреть, с какой скоростью размножаются носители данного аллеля по сравнению со всеми остальными (т.е. с теми, у кого этого аллеля нет).
Если же Fa1/Fa2 ≈ 1, то это нейтральное различие. Если в генофонде был сначала только аллель а1, а потом в результате мутации появился аллель а2, причем Fa1/Fa2 ≈ 1, то это значит, что мутация, в результате которой возник аллель а2, была нейтральной.
Разберемся, как будет меняться частота нейтральной мутации (нейтрального аллеля) со временем, с течением поколений.
· Частота нейтрального аллеля в популяции изменяется по закону случайных блужданий. A random walk is a mathematical formalisation of a trajectory that consists of taking successive random steps. Случайные колебания частот аллелей, не связанные с действием отбора, называются генетическим дрейфом.
· Конечный исход этих случайных блужданий строго предопределен. Это всегда одно из двух: либо аллель зафиксируется (достигнет частоты q=1), либо он элиминируется (достигнет частоты q=0). Третьего не дано. Одно из двух событий рано или поздно ОБЯЗАТЕЛЬНО произойдет. Случайно блуждая, частота аллеля непременно когда-нибудь наткнется либо на верхнюю планку (фиксация), либо на нижнюю (элиминация).
Формула 1. Чему равна вероятность фиксации (Pfix) нейтральной мутации? Если исходная частота q=0.5, то совершенно ясно, что аллель с равной вероятностью зафиксируется или элиминируется, т.е. Pfix = 0.5.
Представим себе ситуацию, когда у нас исходно есть не 2, а 4 нейтральных аллеля с одинаковыми частотами: q1=q2=q3=q4=0.25
Таким образом, можно считать доказанным, что конечным результатом дрейфа будет фиксация одного из аллелей и элиминация трех других. Ясно, что, поскольку исходно все 4 аллеля были в абсолютно равном положении, шансы на фиксацию у них должны быть одинаковы. В сумме эти шансы дают 1 (один из аллелей точно зафиксируется).
Следовательно, Pfix1 = Pfix2 = Pfix3 = Pfix4 = ј = 0.25
Вероятность фиксации нейтрального аллеля равна его исходной частоте (т.е. частоте в тот момент, для которого мы пытаемся рассчитать вероятность).
Поскольку мы могли с тем же успехом взять не 2, не 4, а любое другое число нейтральных аллелей, очевидно, что:
Вероятность фиксации нейтрального аллеля равна его частоте. Это первая из трех формул, которую мы с вами должны вывести. А чему равна вероятность элиминации аллеля? Очевидно, 1-q.
Формула 2. Следующий очень важный вопрос – определить, сколько нейтральных мутаций будет фиксироваться в популяции в каждом поколении. Если мы сможем это установить, то мы обретем великий инструмент – молекулярные часы. Зная, с какой скоростью фиксируются в популяции нейтральные мутации, и зная, сколько их накопилось за какой-то период, мы сможем вычислить длительность этого периода. Например, сколько разных нейтральных мутаций есть в геномах человека и шимпанзе? Это можно подсчитать. Если бы мы еще знали, с какой скоростью фиксируются нейтральные мутации, мы могли бы на основе данных о числе накопившихся мутаций вычислить, как давно разошлись эволюционные линии предков человека и шимпанзе, понять, когда жил их последний общий предок.
Итак, сколько же нейтральных мутаций будет фиксироваться в популяции за одно поколение? Вывод этой формулы – это абсолютнейший шедевр биологической математики. Надеюсь, вы оцените.
Итак, от чего зависит скорость накопления нейтральных мутаций?
Очевидно, она зависит от того, с какой скоростью они появляются – то есть от скорости нейтрального мутагенеза, а также от того, какая часть вновь появляющихся нейтральных мутаций будет фиксироваться (а не элиминироваться).
· V – искомая величина: среднее количество нейтральных мутаций, которые будут фиксироваться в генофонде популяции за одно поколение.
· M – темп появления новых нейтральных мутаций (на всю популяцию). Это среднее количество новых нейтральных мутаций, появляющихся в популяции за одно поколение.
· m – то же, в расчете на одну особь. Это среднее количество новых нейтральных мутаций, имеющихся в геноме родившейся особи. Эту величину реально можно измерить.
· Pfix – доля (или процент) возникших в популяции нейтральных мутаций, которые в конечном счете зафиксируются. Эта доля, очевидно, есть то же самое, что вероятность фиксации такой мутации.
· N – численность популяции. Это тоже можно измерить (правда, она могла меняться в прошлом, и мы не знаем, как она менялась – это плохо).
Приступим к выводу формулы.
Очевидно, что V = M• Pfix
Искомая величина V равна числу мутаций, появляющихся за поколение, умноженному на ту долю, которая зафиксируется из них в конечном счете.
Чему равно M? Очевидно, что M = N•m
Это, надеюсь, ясно без пояснений.
А как найти Pfix, т.е. как выразить эту величину через те величины, которые можно измерить? Ну, мы ведь уже знаем, что Pfix = q , т.е. для каждой мутации вероятность зафиксироваться равна частоте этой мутации в данный момент. Новая, только что появившаяся мутация, очевидно, изначально есть только у одной особи. То есть ее частота q=1/N. Следовательно, и вероятность ее фиксации равна 1/N. Следовательно, из всех новых мутаций зафиксируется их часть, равная 1/N. Вот вам и искомое значение Pfix:
V = M• Pfix M = N•m Pfix =1/N
Подставляем в первую формулу две другие, получаем:
Численность популяции великолепным образом сокращается! Итого:
V = m
Поэтому у нас есть молекулярные часы. Мы их имеем благодаря генетическому дрейфу и благодаря тому, что многие мутации нейтральны. Принцип молекулярных часов:
где D – количество нейтральных различий между двумя видами, t – время, прошедшее с тех пор, когда жил последний общий предок этих двух видов. Двойка тут появляется потому, что нейтральные мутации накапливались обоими видами. Каждый из них накопил за время t количество нейтральных мутаций, равное D/2.
Дарвин (рекомендации по эссе).
Главный вклад Дарвина в умственное развитие человечества. До Дарвина нам был известен только ОДИН механизм возникновения сложных объектов, предназначенных для какой-то цели или функции – разумный дизайн. Пейли и часы. Все подобные объекты трактовались как результат чьего-то сознательного проектирования. Или придумывались некие невнятные дополнительные сущности – жизненная сила, стремление к совершенству и т.п. (ламаркизм, номогенез). Что с научной точки зрения ничуть не лучше гипотезы о разумном дизайне. Дарвин открыл другой механизм – механизм самопроизвольного возникновения сложных объектов, приспособленных для выполнения функций – объектов, которые выглядят (для нас) так, как будто бы их кто-то нарочно спроектировал.
Читайте также: