
Морфология метафазной хромосомы кратко
Обновлено: 05.07.2024
Хромосомы — это основные структурные элементы клеточного ядра, являющиеся носителями генов, в которых закодирована наследственная информация. Обладая способностью к самовоспроизведению, хромосомы обеспечивают генетическую связь поколений.
Метафазная хромосома состоит из двух продольных субъединиц — хроматид.
Так как к моменту деления хромосомы удвоены, то сост. из двух нитей – хроматид. Обе хроматиды объединены между собой в области первичной перетяжки – центромеры. Центромера делит хромосому поперек на две части – плечи, которые бывают короткими (p) и длинными (q). В зависимости от расположения центромеры различают 3 типа хромосом
Метацентрические или равноплечие (с центромерой посередине)
Субметацентрические или неравноплечие (с центромерой,сдвинутой к одному из концов)
Субтелоцентрические или резко неравноплечие
4.акроцентрические или палочковидные (с центромерой,расположенной очень близко к концу хромосомы)
Размеры хромосом растений и животных значительно колеблются: от долей микрона до десятков микрон.
Химической основой строения хромосом являются нуклеопротеиды — комплексы нуклеиновых кислот с основными белками — гистонами и протаминами.
Метафаза занимает значительную часть периода митоза, и отличается относительно стабильным состоянием. Все это время хромосомы удерживаются в экваториальной плоскости веретена за счёт сбалансированных сил натяжения микротрубочек. В метафазе, также как и в течение других фаз митоза, продолжается активное обновление микротрубочек веретена путём интенсивной сборки и деполимеризации молекул тубулина. К окончанию метафазы наблюдается чёткое обособление сестринских хроматид, соединение между которыми сохраняется лишь в центромерных участках. Плечи хроматид располагаются параллельно друг другу, и становится отчетливо заметной разделяющая их щель.
Кариотиип — совокупность признаков (число, размеры, форма и т.д.) полного набора хромосом, присущий клеткам опр биологического вида (видовой кариотип), данного организма (индивидуальный кариотип) или линии (клона) клеток. Видовая спец-ть кариотипазаключается в том, что каждый вид имеет определенные число, форму и размеры хромосом. Каждая хромосома внутри одной группы сцепления занимает определенный, свой локус. Если говорить о парах гомологичных хромосом с их гомологичными локусами, то они отличаются лишь характером аллельных генов, но не по форме и размерам. Хромосомы имеют определенный набор генов, и каждый ген занимает определенное место относительно других (локус-это место внутри хромосомы, в котором находится ген). Это также очень специфичный признак не только для каждой хромосомы из генофонда вида, но, тем более, для хромосом из набора различных видов.Допустим, в одной хромосоме человека есть гены А,В иС, они расположены в соответствующем порядке. Если между локусами генов А и В расстояние 8 морганид,а между генами В и С-10 морганид, следовательно между А и С будет 18 морганид
. Н-р. ген А находится в одном локусе, ген В в другом, ген С -в третьем. Тогда, если знать расстояния ,измеряемые в морганидах,между генами А и В,В и С, то можно узнать и расстояние между генами А и С- сложить 8 и 10=18
Биологические аспекты полового диморфизма.
Половой диморфизм – это анатомические различия между самками и самцами одного и того же биологического вида, помимо половых органов. Выделяют такие компоненты полового диморфизма:
генетический; гормональный; морфологический; поведенческий; психологический.
Первые три связаны со строением организма, остальные, в основном, определяются особенностями воспитания и влияния общества.
Соотносительная роль наследственной программы и факторов среды в формировании фенотипа особи может быть прослежена на примере развития признаков половой принадлежности организма.
Пол организма представляет собой важную фенотипическую характеристику, которая проявляется в совокупности свойств, обеспечивающих воспроизведение потомства и передачу ему наследственной информации. В зависимости от значимости этих свойств различают первичные и вторичные половые признаки.
Под первичными половыми признаками понимают морфофизиологические особенности организма, обеспечивающие образование половых клеток — гамет, сближение и соединение их в процессе оплодотворения. Это наружные и внутренние органы размножения.
Вторичными половыми признаками называют отличительные особенности того или другого пола, не связанные непосредственно с гаметогенезом, спариванием и оплодотворением, но играющие важную роль в половом размножении. Их развитие контролируется гормонами, синтезируемыми первичными половыми органами.
Важным доказательством в пользу наследственной детерминированности половой принадлежности организмов является наблюдаемое у большинства видов соотношение по полу 1:1.
Такое соотношение может быть обусловлено образованием двух видов гамет представителями одного пола (гетерогаметный пол) и одного вида гамет — особями другого пола (гомогаметный пол).
У разных видов организмов хромосомный механизм определения пола реализуется по-разному. У человека и других млекопитающих, а также у дрозофилы гомогаметным является женский пол (XX), а гетерогаметным — мужской (XY). У некоторых насекомых (клопы рода Protenor) гетерогаметный мужской пол имеет лишь одну Х-хромосому (ХО). У птиц и некоторых насекомых женский пол является гетерогаметным (XY), а мужской — гомогаметным (XX).
Таким образом, хромосомный механизм определения половой принадлежности организмов обеспечивает равновероятность встречаемости представителей обоих полов. Это имеет большой биологический смысл, так как обусловливает максимальную вероятность встречи самки и самца, потомки получают более разнообразную наследственную информацию, поддерживается оптимальная численность особей в популяции.
Доказательства роли факторов среды в развитии признаков пола
У большинства видов развитие признаков пола осуществляется на основе наследственной программы, заключенной в генотипе. Однако известны примеры, когда половая принадлежность организма целиком зависит от условий, в которых он развивается. Так, у морского червя Bonellia viridis пол зависит от того, будет зигота развиваться в непосредственном контакте с материнским организмом или самостоятельно. В первом случае образуется самец, во втором —самка.
У высших организмов значение среды в определении признаков пола, как правило, невелико. Вместе с тем даже для них известны примеры, когда в определенных условиях развития происходило переопределение пола на противоположный, несмотря на имеющуюся комбинацию хромосом в зиготе.
Ведущим началом в дифференцировке пола являются гены, контролирующие уровень мужских и женских половых гормонов. Соотношение этих гормонов в организме является особенно важным для формирования и поддержания соответствующих признаков. Так как у млекопитающих и человека дифференцировка пола начинается очень рано, то полное переопределение пола в эмбриогенезе у них невозможно. (стр.230 Ярыгин)
Хромосомы лучше всего изучать во время метафазы митоза, т.к. в этой фазе они:
- располагаются в центре клетки, образуя метафазную пластинку;
- максимально конденсированы и легко различимы с использованием световой микроскопии;
- являются двухроматидными, а сестринские хроматиды соединены между собой в области центромеры, что позволяет различить их морфологию.

Морфологическими элементами метафазной хромосомы являются:
- ломкие (фрагильные) участки.
Хроматида представлена одной линейной молекулой ДНК, ассоциированной с гистоновыми и негистоновыми белками и максимально конденсированной. Метафазная хромосома состоит из двух сестринских хроматид, являющихся результатом репликации ДНК в фазе S и, таким образом, генетически идентичных. Хроматиды одной хромосомы соединены в области центромеры и остаются в таком состоянии до анафазы.
Центромера, или первичная перетяжка, представляет собой специфический участок хромосомы из ДНК и специальных центромерных белков (CENP-A,B,C,D,E). Центромерная ДНК состоит из высокоповторяющихся последовательностей (сателлитная ДНК), практически одинаковых для всех хромосом. Положение центромеры в хромосоме постоянно и специфично для каждой хромосомы/пары гомологичных хромосом. Центромера делит хромосому на два плеча: р (проксимальное) и q (дистальное). По положению центромеры хромосомы делятся на:
- метацентрические - центромера расположена посередине и плечи равные;
- субметацентрические - центромера несколько смещена к одному из концов, а плечи имеют разную длину;
- акроцентрические - центромера значительно смещена к концу хромосомы, из-за чего одно плечо намного короче другого.
Центромеры выполняют следующие функции:
- созревание кинетохоров для прикрепления хромосом к нитям веретена деления;
- продольное расщепление и разделение сестринских хроматид с образованием из одной двухроматидной хромосомы двух однохроматидных хромосом;
- точное и равное распределение генетического материала во время митоза, точная передача генетической информации от клетки к клетке.
Теломеры представлены специфическими последовательностями ДНК на концах хромосом в комплексе со специальными белками. В состав теломерной ДНК входят: (а) тандемно и многократно повторяющиеся короткие последовательности (TTAGGG), одинаковые у всех хромосом, (b) специфические для каждой хромосомы последовательности ДНК.
Теломеры выполняют следующие функции:
- защищают концы хромосом от действия нуклеаз;
- предотвращают слипание концов хромосом;
- обеспечивают репликацию всей ДНК;
- предотвращают укорачивание хромосом благодаря активности теломеразы;
- контролируют процессы старения клеток и многоклеточного организма;
- регулируют фиксацию хроматина к ядерной мембране в интерфазе, обеспечивая тем самым нормальную архитектуру интерфазных хромосом;
- обеспечивают правильную конъюгацию гомологичных хромосом в мейозе.
Вторичные перетяжки (h) представляют собой деспирализованные и слабо окрашенные участки повторяющейся ДНК; в норме они могут быть как в проксимальных плечах (р) акроцентрических хромосом 13, 14, 15, 21, 22, так и в дистальных плечах хромосом 1, 9, 16, реже 4, 6, 10 и Y. Вторичные перетяжки акроцентрических хромосом образуют область ядрышкового организатора. Длина вторичной перетяжки может варьировать в пределах нормального индивидуального полиморфизма.
Сателлиты - это терминальные участки коротких плеч акроцентрических хромосом 13, 14, 15, 21, 22, отделенные вторичной перетяжкой и состоящие из конститутивного гетерохроматина; число и размеры сателлитов варьируют от индивида к индивиду.
Ломкие (фрагильные) участки представляют собой деконденсированные сегменты хромосом, отличающиеся повышенной чувствительностью к действию мутагенных факторов, под влиянием которых в них легко происходят разрывы и, в результате этого, хромосомные перестройки. Фрагильные участки:
- являются маркерами нормального индивидуального полиморфизма;
- ассоциированы с некоторыми моногенными синдромами (например, FRAXA и семейная умственная отсталость);
- могут участвовать в опухолевой прогрессии (путем инактивации генов-супрессоров).


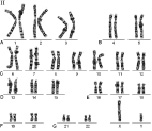
Рис. 1. Мужской кариотип: I – метафазная пластинка; II – классификация по группам и нумерация хромосом
Как указано выше, кариотип человека состоит из 46 хромосом, которые нумеруются от 1 до 22 (аутосомы) и делятся на 7 групп, – A, B, C, D, E, F, G и половые хромосомы (гоносомы) X и Y (рис. 1 и 2).
К первой группе А относятся хромосомы 1, 2 и 3, которые хорошо отличаются друг от друга. Хромосома 1 (размер – 11 мкм) – метацентрическая, содержит вторичную перетяжку в околоцентромерном участке длинного плеча. Хромосома 2 (10,8 мкм) по размерам почти равна хромосоме 1 и является субметацентрической. Хромосома 3 (размер – 8,3 мкм) – практически метацентрическая.
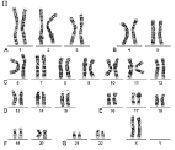
Рис. 2. Женский кариотип: I – метафазная пластинка; II – классификация по группам и нумерация хромосом
К группе В относятся хромосомы 4 и 5 (размер – 7,7 мкм каждая) – это крупные субметацентрические хромосомы, которые не отличаются друг от друга при рутинном окрашивании ни размером, ни положением центромер.
К группе С относятся хромосомы с 6 по 12 и Х. В основном, это субметацентрические хромосомы крупных и средних размеров. Наиболее крупные хромосомы из группы С – 6, 7 и Х (6,8–7,2 мкм). Хромосома Х является половой хромосомой (гоносомой). Хромосома 7 более метацентрична, чем хромосома 6. Хромосомы 8 и 9 – практически одинаковы по размеру (5,8 мкм). Хромосома 8 метацентричнее хромосомы 9, которая характеризуется регулярной вторичной перетяжкой в прицентромерном районе длинного плеча.
К группе D относятся хромосомы 13, 14 и 15 (4,2 мкм) – средних размеров акроцентрические хромосомы с почти терминальным расположением центромеры. Эти хромосомы между собой не различаются ни по размерам, ни морфологически после рутинного окрашивания. Короткое плечо всех трёх пар хромосом может формировать спутники (рис. 1, 2 и 5).
К группе Е относятся хромосомы 16, 17 и 18. Хромосома 16 (размер – 3,6 мкм) – сравнительно небольшая метацентрическая хромосома, содержащая вторичную перетяжку в длинном плече. Хромосома 17 (размер – 3,5 мкм) – сравнительно короткая субметацентрическая хромосома. Хромосома 18 (размер – 3,2 мкм) – самая короткая субметацентрическая хромосома.
Группа F представлена хромосомами 19 и 20 (размеры – 2,9 мкм). Это короткие метацентрические хромосомы, которые не отличаются между собой без дифференциального окрашивания по длине.
Хромосомы 21 и 22 (2,8 мкм) относятся к группе G. Это наиболее короткие акроцентрические хромосомы в кариотипе, которые обладают способностью формировать спутники на коротком плече (рис. 1, 2 и 5).
Хромосома Y (2,3 мкм) является маленькой акроцентрической хромосомой, сравнимой по размерам с хромосомами 21 и 22, но не имеющая спутников.
Следует отметить, что хромосомные синдромы и аномалии связаны с хромосомными (геномными) мутациями (аномалиями) в виде различных структурных перестроек хромосом или с изменением их числа (n). Численные изменения хромосом могут быть двух типов: полиплоидии – умножение хромосомного набора (3n, 4n и т. д.) или генома, кратное гаплоидному числу хромосом; анеуплоидии – увеличение или уменьшение числа хромосом, некратное гаплоидному. Структурные хромосомные (геномные) перестройки классифицируют по принципу линейной последовательности расположения генов: делеции (потеря хромосомных участков), дупликации (удвоение хромосомных участков), инверсии (перевертывание на 180° относительно нормальной последовательности хромосомных участков), инсерции (вставки хромосомных участков) и транслокации (изменение расположения хромосомных участков). Подробная информация по возможным аномалиям хромосом человека представлена в главе 3.5.
Как правило, эукариотическая клетка имеет одно ядро, но встречаются двуядерные (инфузории) и многоядерные клетки (опалина). Некоторые высокоспециализированные клетки вторично утрачивают ядро (эритроциты млекопитающих, ситовидные трубки покрытосеменных).
Форма ядра — сферическая, эллипсовидная, реже лопастная, бобовидная и др. Диаметр ядра — обычно от 3 до 10 мкм.

Строение ядра:
1 — наружная мембрана; 2 — внутренняя мембрана; 3 — поры; 4 — ядрышко; 5 — гетерохроматин; 6 — эухроматин.
Ядро отграничено от цитоплазмы двумя мембранами (каждая из них имеет типичное строение). Между мембранами — узкая щель, заполненная полужидким веществом. В некоторых местах мембраны сливаются друг с другом, образуя поры (3), через которые происходит обмен веществ между ядром и цитоплазмой. Наружная ядерная (1) мембрана со стороны, обращенной в цитоплазму, покрыта рибосомами, придающими ей шероховатость, внутренняя (2) мембрана гладкая. Ядерные мембраны являются частью мембранной системы клетки: выросты наружной ядерной мембраны соединяются с каналами эндоплазматической сети, образуя единую систему сообщающихся каналов.
Кариоплазма (ядерный сок, нуклеоплазма) — внутреннее содержимое ядра, в котором располагаются хроматин и одно или несколько ядрышек. В состав ядерного сока входят различные белки (в том числе ферменты ядра), свободные нуклеотиды.
Ядрышко (4) представляет собой округлое плотное тельце, погруженное в ядерный сок. Количество ядрышек зависит от функционального состояния ядра и варьирует от 1 до 7 и более. Ядрышки обнаруживаются только в неделящихся ядрах, во время митоза они исчезают. Ядрышко образуется на определенных участках хромосом, несущих информацию о структуре рРНК. Такие участки называются ядрышковым организатором и содержат многочисленные копии генов, кодирующих рРНК. Из рРНК и белков, поступающих из цитоплазмы, формируются субъединицы рибосом. Таким образом, ядрышко представляет собой скопление рРНК и рибосомальных субъединиц на разных этапах их формирования.
Хроматин — внутренние нуклеопротеидные структуры ядра, окрашивающиеся некоторыми красителями и отличающиеся по форме от ядрышка. Хроматин имеет вид глыбок, гранул и нитей. Химический состав хроматина: 1) ДНК (30–45%), 2) гистоновые белки (30–50%), 3) негистоновые белки (4–33%), следовательно, хроматин является дезоксирибонуклеопротеидным комплексом (ДНП). В зависимости от функционального состояния хроматина различают: гетерохроматин (5) и эухроматин (6). Эухроматин — генетически активные, гетерохроматин — генетически неактивные участки хроматина. Эухроматин при световой микроскопии не различим, слабо окрашивается и представляет собой деконденсированные (деспирализованные, раскрученные) участки хроматина. Гетерохроматин под световым микроскопом имеет вид глыбок или гранул, интенсивно окрашивается и представляет собой конденсированные (спирализованные, уплотненные) участки хроматина. Хроматин — форма существования генетического материала в интерфазных клетках. Во время деления клетки (митоз, мейоз) хроматин преобразуется в хромосомы.
Функции ядра: 1) хранение наследственной информации и передача ее дочерним клеткам в процессе деления, 2) регуляция жизнедеятельности клетки путем регуляции синтеза различных белков, 3) место образования субъединиц рибосом.
Хромосомы

Хромосомы — это цитологические палочковидные структуры, представляющие собой конденсированный хроматин и появляющиеся в клетке во время митоза или мейоза. Хромосомы и хроматин — различные формы пространственной организации дезоксирибонуклеопротеидного комплекса, соответствующие разным фазам жизненного цикла клетки. Химический состав хромосом такой же, как и хроматина: 1) ДНК (30–45%), 2) гистоновые белки (30–50%), 3) негистоновые белки (4–33%).
Основу хромосомы составляет одна непрерывная двухцепочечная молекула ДНК; длина ДНК одной хромосомы может достигать нескольких сантиметров. Понятно, что молекула такой длины не может располагаться в клетке в вытянутом виде, а подвергается укладке, приобретая определенную трехмерную структуру, или конформацию. Можно выделить следующие уровни пространственной укладки ДНК и ДНП: 1) нуклеосомный (накручивание ДНК на белковые глобулы), 2) нуклеомерный, 3) хромомерный, 4) хромонемный, 5) хромосомный.
В процессе преобразования хроматина в хромосомы ДНП образует не только спирали и суперспирали, но еще петли и суперпетли. Поэтому процесс формирования хромосом, который происходит в профазу митоза или профазу 1 мейоза, лучше называть не спирализацией, а конденсацией хромосом.

Хромосомы: 1 — метацентрическая; 2 — субметацентрическая; 3, 4 — акроцентрические. Строение хромосомы: 5 — центромера; 6 — вторичная перетяжка; 7 — спутник; 8 — хроматиды; 9 — теломеры.
Метафазная хромосома (хромосомы изучаются в метафазу митоза) состоит из двух хроматид (8). Любая хромосома имеет первичную перетяжку (центромеру) (5), которая делит хромосому на плечи. Некоторые хромосомы имеют вторичную перетяжку (6) и спутник (7). Спутник — участок короткого плеча, отделяемый вторичной перетяжкой. Хромосомы, имеющие спутник, называются спутничными (3). Концы хромосом называются теломерами (9). В зависимости от положения центромеры выделяют: а) метацентрические (равноплечие) (1), б) субметацентрические (умеренно неравноплечие) (2), в) акроцентрические (резко неравноплечие) хромосомы (3, 4).
Соматические клетки содержат диплоидный (двойной — 2n) набор хромосом, половые клетки — гаплоидный (одинарный — n). Диплоидный набор аскариды равен 2, дрозофилы — 8, шимпанзе — 48, речного рака — 196. Хромосомы диплоидного набора разбиваются на пары; хромосомы одной пары имеют одинаковое строение, размеры, набор генов и называются гомологичными.
Кариотип — совокупность сведений о числе, размерах и строении метафазных хромосом. Идиограмма — графическое изображение кариотипа. У представителей разных видов кариотипы разные, одного вида — одинаковые. Аутосомы — хромосомы, одинаковые для мужского и женского кариотипов. Половые хромосомы — хромосомы, по которым мужской кариотип отличается от женского.
Хромосомный набор человека (2n = 46, n = 23) содержит 22 пары аутосом и 1 пару половых хромосом. Аутосомы распределены по группам и пронумерованы:
| Группа | Число пар | Номер | Размер | Форма |
|---|---|---|---|---|
| A | 3 | 1, 2, 3 | Крупные | 1, 3 — метацентрические, 2 — субметацентрические |
| B | 2 | 4, 5 | Крупные | Субметацентрические |
| C | 7 | 6, 7, 8, 9, 10, 11, 12 | Средние | Субметацентрические |
| D | 3 | 13, 14, 15 | Средние | Акроцентрические, спутничные (вторичная перетяжка в коротком плече) |
| E | 3 | 16, 17, 18 | Мелкие | Субметацентрические |
| F | 2 | 19, 20 | Мелкие | Метацентрические |
| G | 2 | 21, 22 | Мелкие | Акроцентрические, спутничные (вторичная перетяжка в коротком плече) |
Половые хромосомы не относятся ни к одной из групп и не имеют номера. Половые хромосомы женщины — ХХ, мужчины — ХУ. Х-хромосома — средняя субметацентрическая, У-хромосома — мелкая акроцентрическая.
В области вторичных перетяжек хромосом групп D и G находятся копии генов, несущих информацию о строении рРНК, поэтому хромосомы групп D и G называются ядрышкообразующими.
Функции хромосом: 1) хранение наследственной информации, 2) передача генетического материала от материнской клетки к дочерним.
Читайте также: